
Upregulation of miR-33 Exacerbates Heat-Stress-Induced Apoptosis in Granulosa Cell and Follicular Atresia of Nile Tilapia (Oreochromis niloticus) by Targeting TGFβ1I1


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Fish
2.2. Experimental Procedure
2.3. Sampling
2.4. Observation of Oocyte Development and Apoptosis
2.5. miRNA Sequencing and Validation
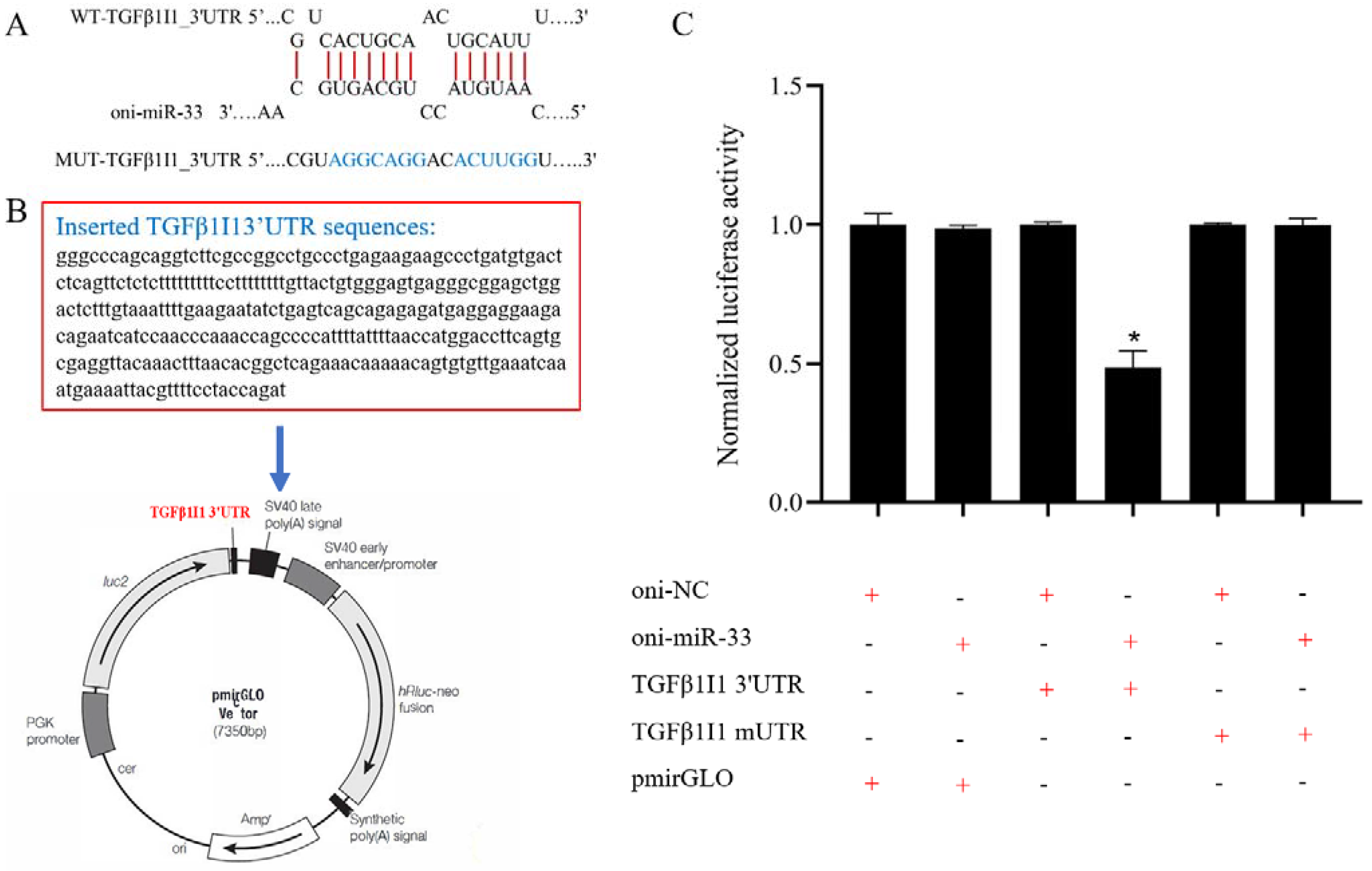
2.6. Validation of miRNA Target Genes
2.7. Detection of FSH, LH, and E2 Contents
2.8. Western Blot (WB) Analyses
2.9. Immunohistochemical Staining
2.10. Analysis of Gene Transcript Levels
2.11. Data Analysis
3. Results
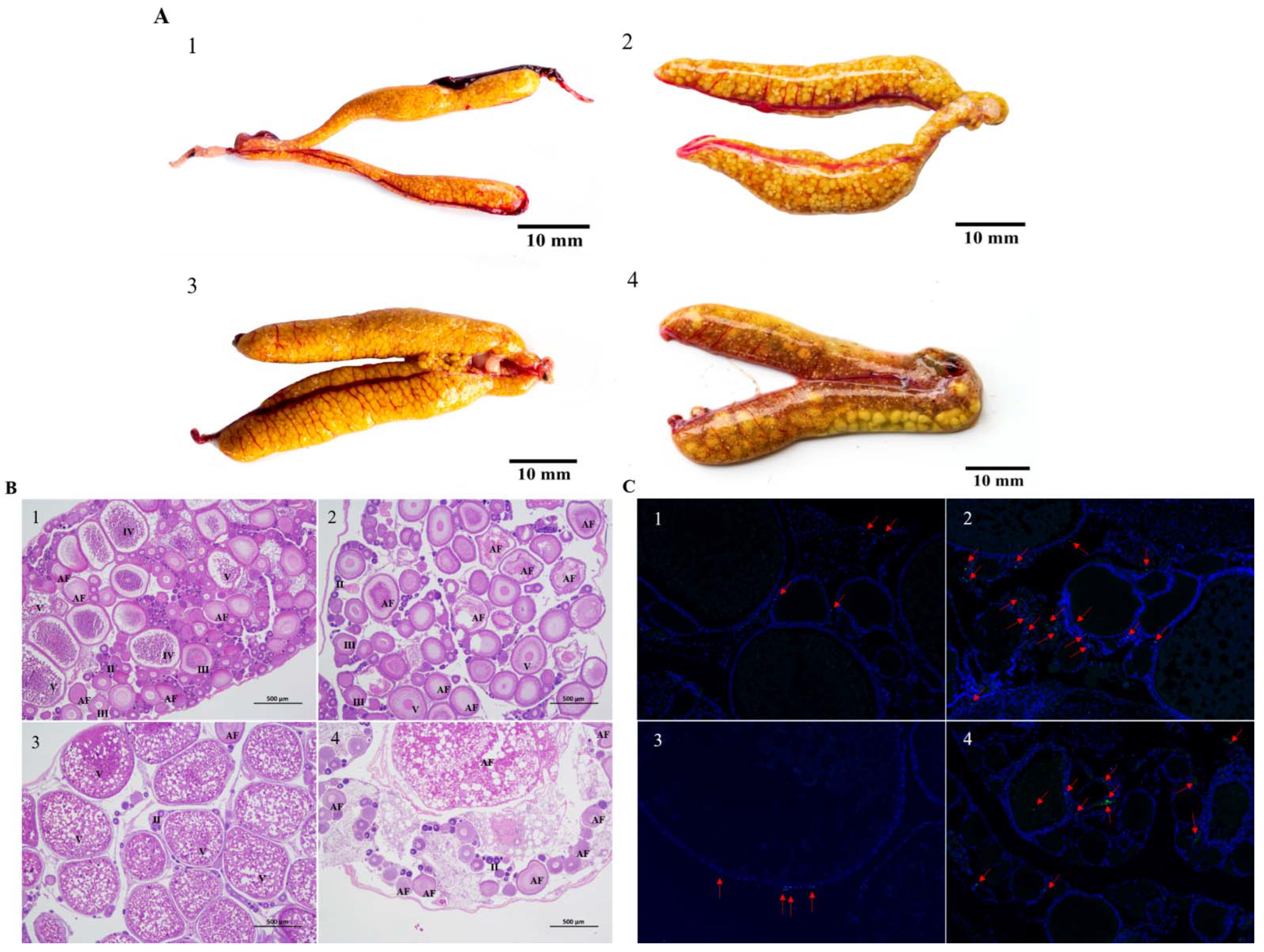
3.1. Morphological and Histological Observations of Nile Tilapia Ovaries at 6 d and 12 d of Heat Stress
3.2. Apoptosis in Nile Tilapia Ovarian Granulosa Cells at 6 d and 12 d of Heat Stress
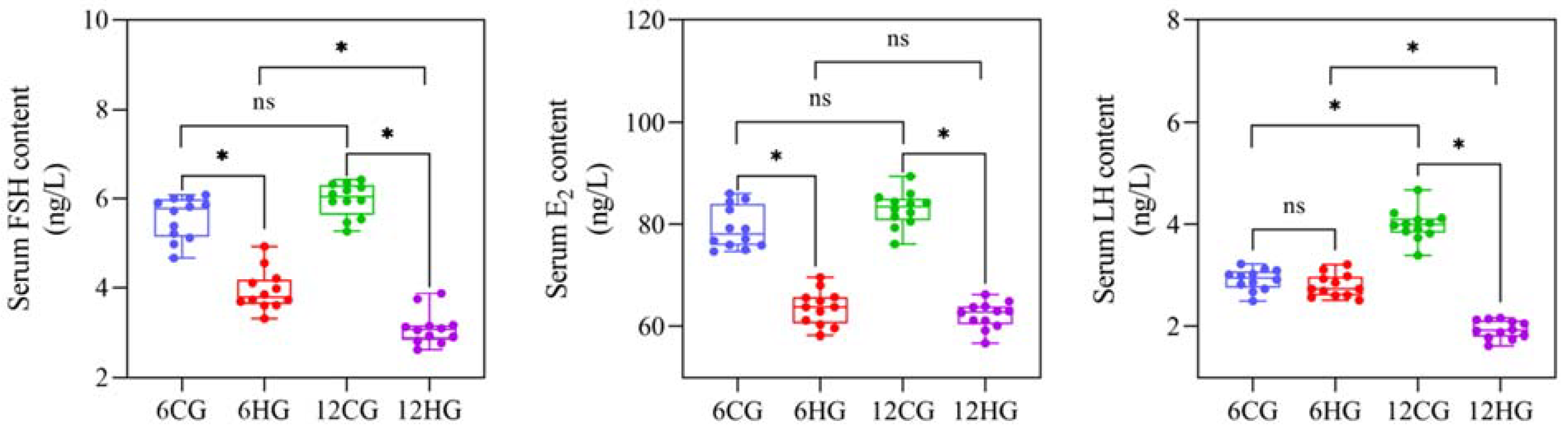
3.3. Serum FSH, LH, and E2 Contents of Nile Tilapia at 6 d and 12 d of Heat Stress
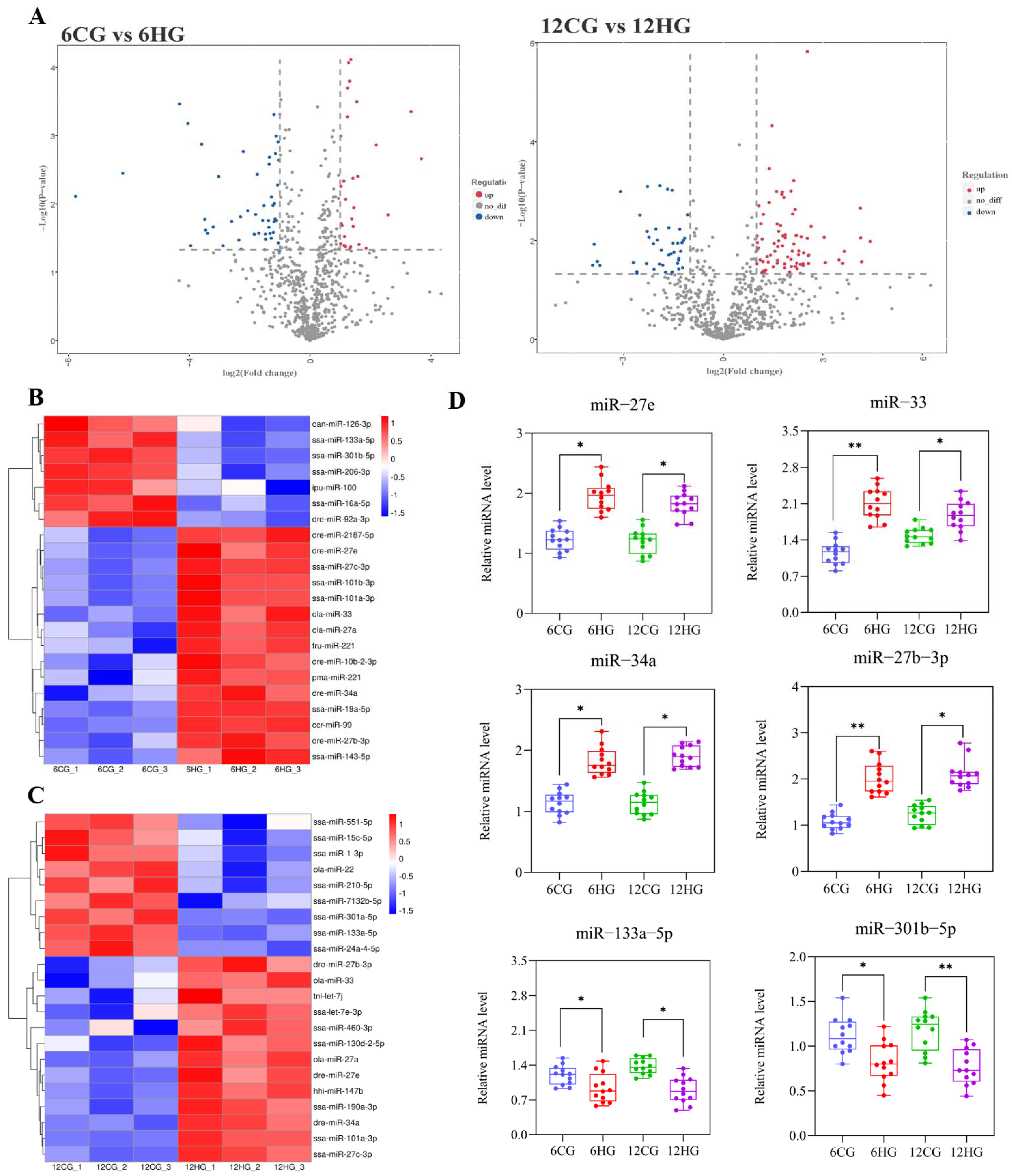
3.4. DE miRNAs between Control and Heat-Stressed Nile Tilapi
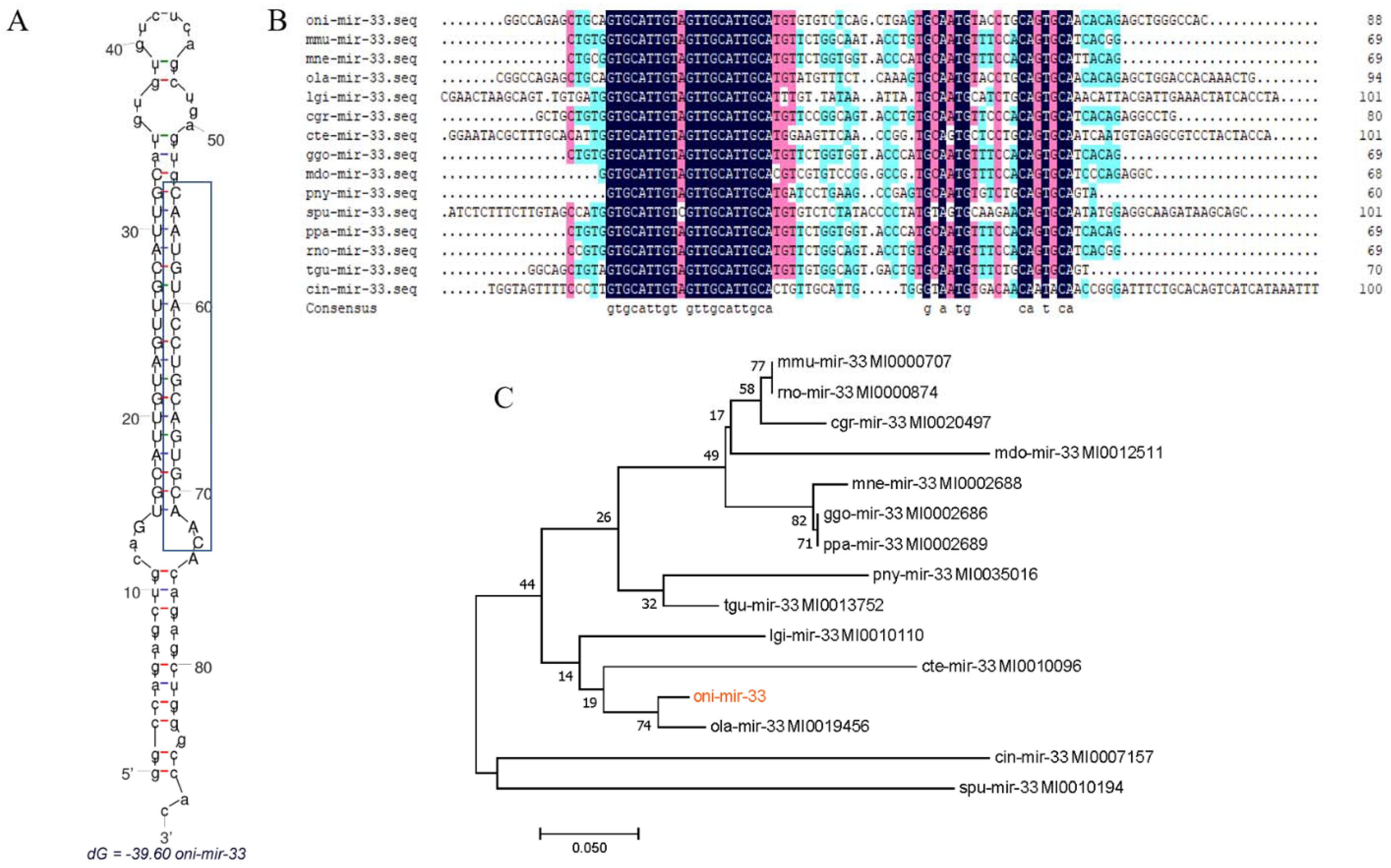
3.5. Analysis of miR-33 Sequence
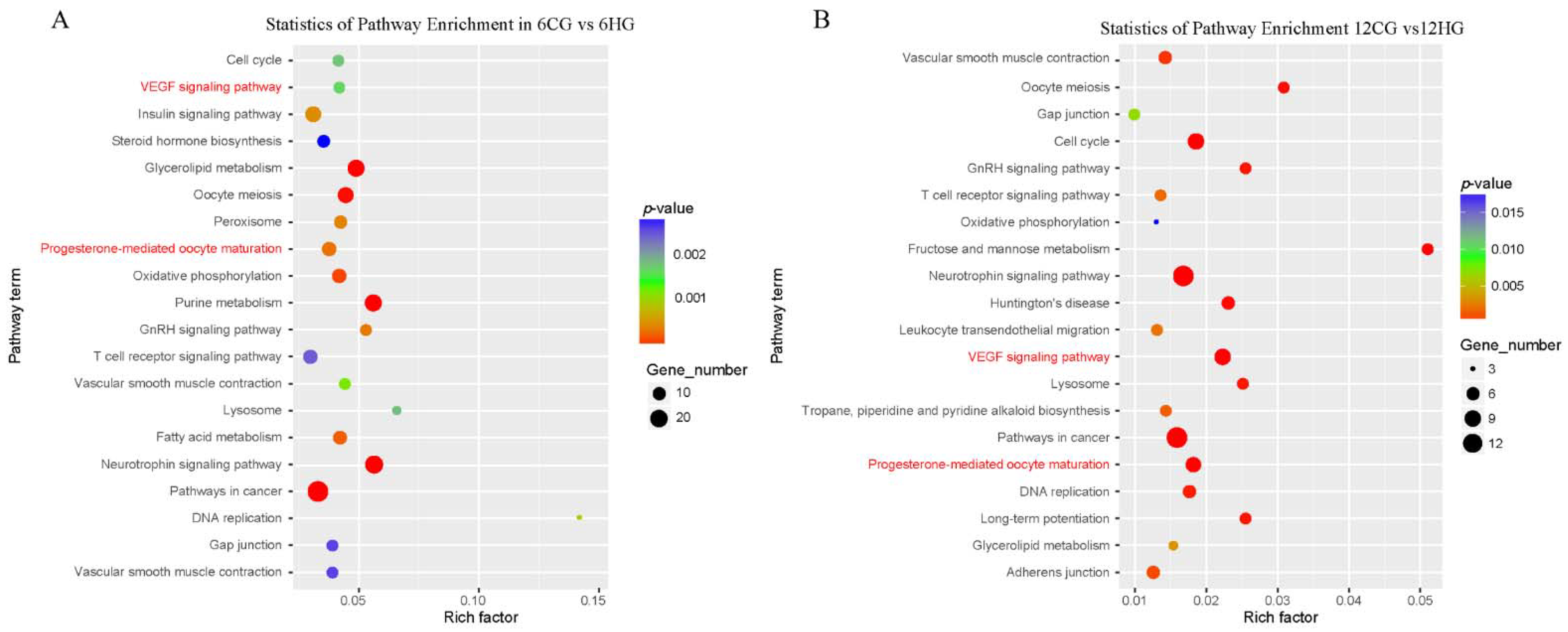
3.6. Enrichment Analysis to Identify Potential Target Genes of miR-33
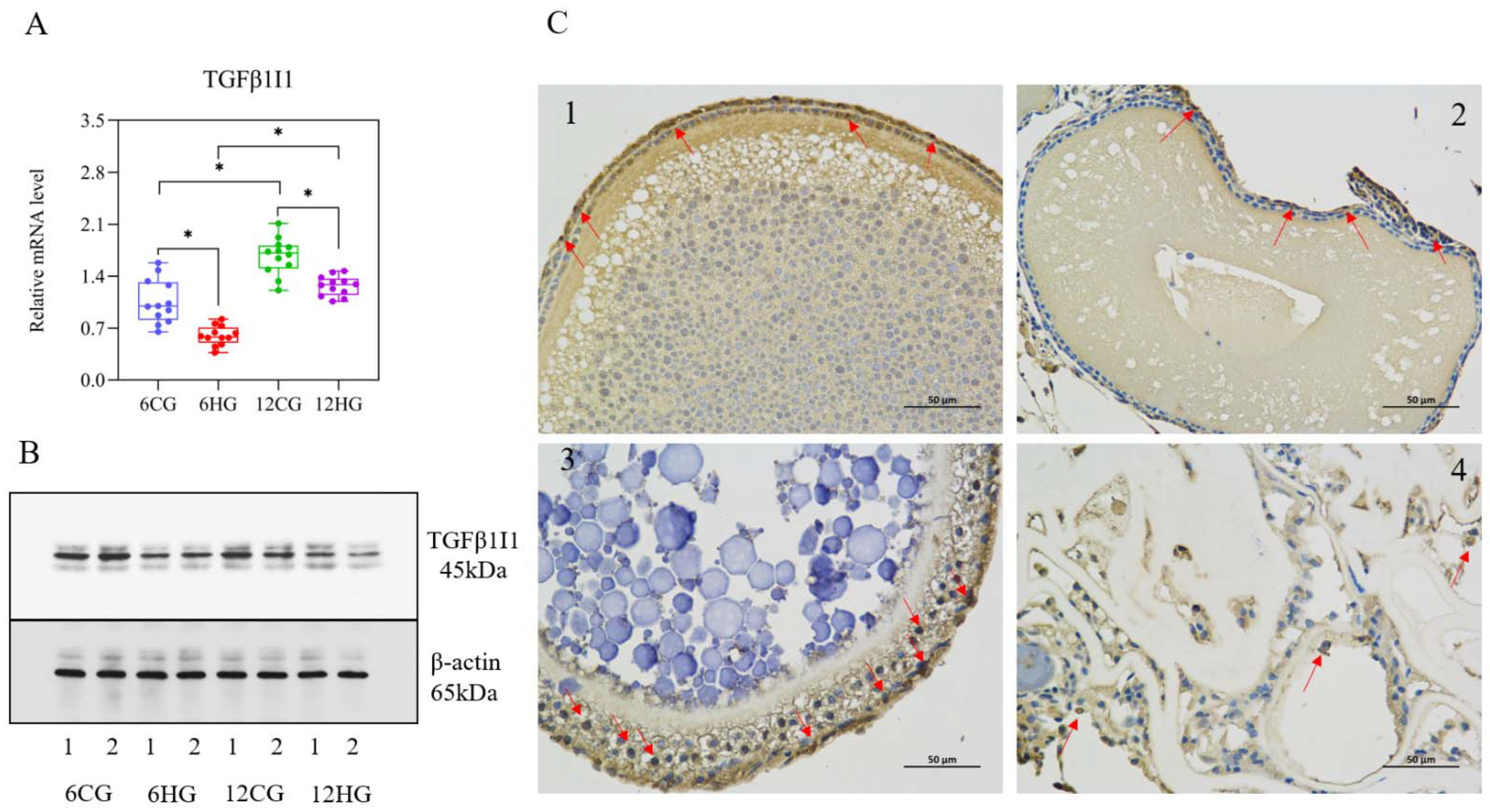
3.7. miR-33 Directly Regulates the Expression of TGFβ1I1
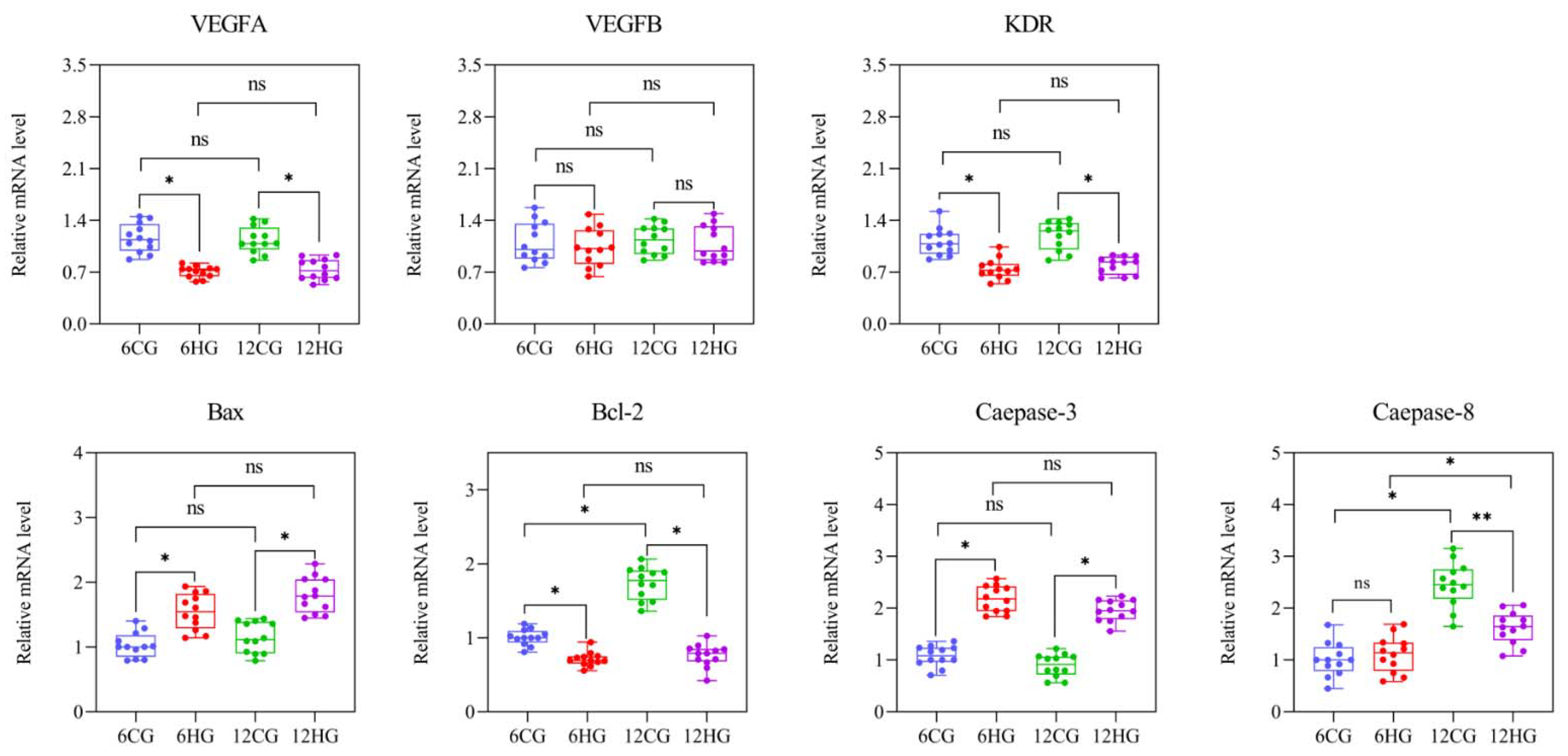
3.8. miR-33 Participates in Apoptosis of Nile Tilapia Follicle Granulosa Cells through VEGF Signaling
4. Discussion
4.1. miRNA Profiles Reveal Multiple Key miRNAs Involved in Development and Atresia
4.2. miR-33 Directly Interacts with the 3′-UTR of TGFβ1I1
4.3. miR-33 Interferes with Steroid Hormone Synthesis by Targeting TGFβ1I1
4.4. miR-33 Targets TGFβ1I1 to Mediate VEGF Signaling, Contributing to the Regulation of Granulosa Cell Apoptosis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Bromage, N.R.; Roberts, R.J. Broodstock Management and Egg and Larval Quality; Blackwell Science: Oxford, UK, 1995; p. 424. [Google Scholar]
- Brooks, S.; Tyler, C.R.; Sumpter, J.P. Egg quality in fish: What makes a good egg? Rev. Fish Biol. Fish. 1997, 7, 387–416. [Google Scholar] [CrossRef]
- Liu, H.W.; Stickney, R.R.; Dickhoff, W.W.; McCaughran, D.A. Effects of environmental factors on egg development and hatching of pacific halibut Hippoglossus stenolepis. J. World Aquac. Soc. 1994, 25, 317–321. [Google Scholar] [CrossRef]
- Getinet, G.T. Effects of maternal age on fecundity, spawning interval, and egg quality of Nile tilapia, Oreochromis niloticus (L.). J. World Aquac. Soc. 2008, 39, 671–677. [Google Scholar] [CrossRef]
- Qiang, J.; He, J.; Zhu, J.-H.; Tao, Y.-F.; Bao, J.-W.; Yan, Y.; Xu, P.; Zhu, X.W. Optimal combination of temperature and photoperiod for sex steroid hormone secretion and egg development of Oreochromis niloticus as determined by response surface methodology. J. Therm. Biol. 2021, 97, 102889. [Google Scholar] [CrossRef] [PubMed]
- Taranger, G.L.; Haux, C.; Stefansson, S.O.; Björnsson, B.T.; Hansen, T. Abrupt changes in photoperiod affect age at maturity, timing of ovulation and plasma testosterone and oestradiol-17b profiles in Atlantic salmon, Salmo salar. Aquaculture 1998, 162, 85–98. [Google Scholar] [CrossRef]
- Bonnet, E.; Montfort, J.; Esquerre, D.; Hugot, K.; Fostier, A.; Bobe, J. Effect of photoperiod manipulation on rainbow trout (Oncorhynchus mykiss) egg quality: A genomic study. Aquaculture 2007, 268, 13–22. [Google Scholar] [CrossRef]
- Corriero, A.; Zupa, R.; Bello, G.; Mylonas, C.C.; Deflorio, M.; Genovese, S.; Basilone, G.; Buscaino, G.; Buffa, G.; Pousis, C.; et al. Evidence that severe acute stress and starvation induce rapid atresia of ovarian vitellogenic follicles in Atlantic bluefin tuna, Thunnus thynnus (L.) (Osteichthyes: Scombridae). J. Fish Dis. 2011, 34, 853–860. [Google Scholar] [CrossRef]
- Qiang, J.; Duan, X.J.; Zhu, H.J.; He, J.; Tao, Y.F.; Bao, J.W.; Zhu, X.W.; Xu, P. Some ‘white’ oocytes undergo atresia and fail to mature during the reproductive cycle in female genetically improved farmed tilapia (Oreochromis niloticus). Aquaculture 2021, 534, 736278. [Google Scholar] [CrossRef]
- Aegerter, S.; Jalabert, B.; Bobe, J. Large scale real-time PCR analysis of mRNA abundance in rainbow trout eggs in relationship with egg quality and post-ovulatory ageing. Mol. Reprod. Dev. 2005, 72, 377–385. [Google Scholar] [CrossRef]
- Campbell, P.M.; Pottinger, T.G.; Sumpter, J.P. Stress reduces the quality of gametes produced by rainbow trout. Biol. Reprod. 1992, 47, 1140–1150. [Google Scholar] [CrossRef]
- Schreck, C.B.; Contreras-Sanchez, W.; Fitzpatrick, M.S. Effects of Stress on Fish Reproduction, Gamete Quality, and Progeny. In Reproductive Biotechnology in Finfish Aquaculture; Elsevier: Amsterdam, The Netherlands, 2001; pp. 3–24. [Google Scholar]
- Clearwater, S.J.; Pankhurst, N.W. The response to capture and confinement stress of plasma cortisol, plasma sex steroids and vitellogenic oocytes in the marine teleost, red gurnard. J. Fish Biol. 1997, 50, 429–441. [Google Scholar] [CrossRef]
- Molly, A.H.W.; Joel, P.V.E.; Serge, I.D.; Moberg, G.P. Preliminary observations on the effects of holding temperature on reproductive performance of female white sturgeon, Acipenser transmontanus Richardson. Aquaculture 1999, 176, 315–329. [Google Scholar]
- Pankhurst, N.W.; Purser, G.J.; Kraak, G.V.D.; Thomas, P.M.; Forteath, G.N.R. Effect of holding temperature on ovulation, egg fertility, plasma levels of reproductive hormones and in vitro ovarian steroidogenesis in the rainbow trout Oncorhynchus mykiss. Aquaculture 1996, 146, 277–290. [Google Scholar] [CrossRef]
- Mansouri-Attia, N.; Tripurani, S.K.; Gokul, N.; Piard, H.; Anderson, M.L.; Eldin, K.; Pangas, S.A. TGFβ signaling promotes juvenile granulosa cell tumorigenesis by suppressing apoptosis. Mol. Endocrinol. 2014, 28, 1887–1898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Mu, X.; Guo, M.; Wang, Y.J.; Teng, Z.; Mao, J.P.; Niu, W.-B.; Feng, L.Z.; Zhao, L.H.; Xia, G.L. Transforming growth factor-β signaling participates in the maintenance of the primordial follicle pool in the mouse ovary. J. Biol. Chem. 2014, 289, 8299–8311. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.L.; Wang, C.; Gu, W.B.; Zhu, Q.H.; Wang, L.Z.; Zhou, Z.K.; Liu, Z.P.; Chen, Y.Y.; Shu, M.A. Identification and functional analysis of transforming growth factor-β type I receptor (T β R1) from Scylla paramamosain: The first evidence of T β R1 Check for involved in development and innate immunity in crustaceans. Dev. Comp. Immunol. 2018, 88, 144–151. [Google Scholar] [CrossRef]
- Yang, M.; Zhou, H. Grass carp transforming growth factor-β 1 (TGF-β 1): Molecular cloning, tissue distribution and immunobiological activity in teleost peripheral blood lymphocytes. Mol. Immunol. 2008, 45, 1792–1798. [Google Scholar] [CrossRef]
- Li, B.; Zhou, Y.L.; Gu, W.B.; Wang, L.Z.; Xu, Y.P.; Cheng, Y.X.; Chen, D.Y.; Li, B.W.; Xiao, Y.; Dong, W.R.; et al. Identification and functional analysis of transforming growth factor-β type III receptor (TβR3) from Scylla paramamosain: The first evidence of TβR3 involved in development and innate immunity in invertebrates. Fish Shellfish Immunol. 2020, 105, 41–52. [Google Scholar] [CrossRef]
- Shinji, J.P.; Miyanishi, H.; Gotoh, H.; Kaneko, T. Appendage Regeneration After Autotomy is Mediated by Baboon in the Crayfish Procambarus Fallax F. Virginalis Martin, Dorn, Kawai, Heiden and Scholtz, 2010 (Decapoda: Astacoidea: Cambaridae). J. Crustacean Biol. 2016, 36, 649–657. [Google Scholar] [CrossRef] [Green Version]
- Kohli, G.; Hu, S.Q.; Clelland, E.; Muccio, D.T.; Rothenstein, J.; Peng, C. Cloning of transforming growth factor-β1 (TGF-β1) and its type II receptor from zebrafish ovary and role of TGF-β1 in oocyte maturation. Endocrinology 2003, 144, 1931–1941. [Google Scholar] [CrossRef]
- Kohli, G.; Clelland, E.; Peng, C. Potential targets of transforming growth factor-beta1 during inhibition of oocyte maturation in zebrafish. Reprod. Biol. Endocrinol. 2005, 53, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Q.; Zagrodny, A.; Bernaudo, S.; Peng, C. Regulation of membrane progestin receptors in the zebrafish ovary by gonadotropin, activin, TGF-β and BMP-15. Mol. Cell. Endocrinol. 2009, 312, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Qiang, J.; Tao, Y.F.; Zhu, H.J.; Lu, S.Q.; Cao, Z.M.; Ma, J.L.; He, J.; Xu, P. Effects of heat stress on follicular development and atresia in Nile tilapia (Oreochromis niloticus) during one reproductive cycle and its potential regulation by autophagy and apoptosis. Aquaculture 2022, 555, 738171. [Google Scholar] [CrossRef]
- Ferrari, G.; Cook, D.B.; Terushkin, V.; Pintucci, G.; Mignatti, P. Transforming growth factor-β 1 (TGF-β1) induces angiogenesis through vascular endothelial growth factor (VEGF)-mediated apoptosis. J. Cell. Physiol. 2009, 219, 449–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, H.; Zhang, Q.Y.; Li, J.L.; Zhang, N.; Hua, Y.P.; Xu, L.X.; Deng, Y.; Lai, J.; Peng, Z.; Peng, B.; et al. Apatinib inhibits VEGF signaling and promotes apoptosis in intrahepatic cholangiocarcinoma. Oncotarget 2016, 7, 17220–17229. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Li, X.Y.; Chen, Y.; Zhang, X.; Wu, Y.; Wang, Z.X.; Chen, P.H.; Dai, H.Q.; Feng, J.; Chatterjee, S.; et al. Quercetin induces apoptosis via downregulation of vascular endothelial growth factor/Akt signaling pathway in acute myeloid leukemia cells. Front. Pharmacol. 2020, 11, 534171. [Google Scholar] [CrossRef]
- Ferrari, G.; Pintucci, G.; Seghezzi, G.; Hyman, K.; Galloway, A.C.; Mignatti, P. VEGF, a prosurvival factor, acts in concert with TGF-1 to induce endothelial cell apoptosis. Proc. Natl. Acad. Sci. USA 2006, 103, 17260–17265. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.Y.; Yao, W.; Yao, Y.; Du, X.; Zhou, J.L.; Ma, B.Q. MiR-92a inhibits porcine ovarian granulosa cell apoptosis by targeting Smad7 gene. FEBS Lett. 2014, 588, 4497–4503. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Liu, L.; Li, Q.Q.; Zhang, L.F.; Pan, Z.X.; Li, Q.F. NORFA, long intergenic noncoding RNA, maintains sow fertility by inhibiting granulosa cell death. Commun. Biol. 2020, 3, 131. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.Y.; Zhao, J.; Xu, Q.L.; Guo, Y.L.; Liu, M.Z.; Zhang, C.L.; Schinckel, A.P.; Zhou, B. MiR-31 targets HSD17B14 and FSHR, and miR-20b targets HSD17B14 to affect apoptosis and steroid hormone metabolism of porcine ovarian granulosa cells. Theriogenology 2022, 180, 94–102. [Google Scholar] [CrossRef]
- Qiang, J.; Zhu, X.W.; He, J.; Tao, Y.F.; Bao, J.W.; Zhu, H.J.; Xu, P. miR-34a regulates the activity of HIF-1a and P53 signaling pathways by promoting GLUT1 in genetically improved farmed tilapia (GIFT, Oreochromis niloticus) under hypoxia stress. Front. Physiol. 2020, 11, 670. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.M.; Qiang, J.; Zhu, H.J.; Li, H.X.; Tao, Y.F.; He, J.; Xu, P.; Dong, Z.J. Transcriptional inhibition of steroidogenic factor 1 in vivo in Oreochromis niloticus increased weight and suppressed gonad development. Gene 2022, 809, 146023. [Google Scholar] [CrossRef] [PubMed]
- Reading, J.B.; Andersen, K.L.; Ryu, Y.W.; Mushirobira, Y.J.; Todo, T.; Hiramatsu, N. Oogenesis and egg quality in finfish: Yolk formation and other factors influencing female fertility. Fishes 2018, 3, 45. [Google Scholar] [CrossRef] [Green Version]
- Shiota, M.; Sugai, N.; Tamura, M.; Yamaguchi, R.; Fukushima, N.; Miyano, T.; Miyazaki, H. Correlation of mitogen-activated protein kinase activities with cell survival and apoptosis in porcine granulosa cells. Zool. Sci. 2003, 20, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.N.; Wang, L.; Wang, L.; Chen, Y.R.; Li, F.G. miR-17-5p effects porcine granulosa cell growth and oestradiol synthesis by targeting E2F1 gene. Reprod. Domest. Anim. 2019, 54, 11. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, A.L.; Zhang, Z.; Yuan, X.L.; Chen, Z.M.; Zhang, H.; Li, J.Q. MicroRNA-34c regulates porcine granulosa cell function by targeting forkhead box O3a. J. Integr. Agric. 2017, 16, 2019–2028. [Google Scholar] [CrossRef] [Green Version]
- Nie, M.Y.; Peng, S.Y.S.; Fang, Y.; Wang, H.M.; Yang, X.K. miR-23a and miR-27a promote human granulosa cell apoptosis by targeting SMAD5. Biol. Reprod. 2015, 93, 98. [Google Scholar] [CrossRef]
- Wang, M.; Liu, M.; Sun, J.; Jia, L.; Ma, S.; Gao, J.; Xu, Y.; Zhang, H.; Tsang, S.Y.; Li, X. MicroRNA-27a-3p affects estradiol and androgen imbalance by targeting Creb1 in the granulosa cells in mouse polycytic ovary syndrome model. Biol. Reprod. 2017, 17, 295–304. [Google Scholar] [CrossRef]
- Wang, L.; Li, C.; Li, R.; Deng, Y.; Tan, Y.; Tong, C.; Qi, H. MicroRNA-764-3p regulates 17β-estradiol synthesis of mouse ovarian granulosa cells by targeting steroidogenic factor-1. Vitr. Cell. Dev. Biol. Anim. 2016, 52, 365–373. [Google Scholar] [CrossRef]
- Gnanaguru, G.; Wagschal, A.; Oh, J.; Saez-Torres, K.L.; Li, T.; Temel, R.E.; Kleinman, M.E.; Näär, A.M.; D’Amore, P.A. Targeting of miR-33 ameliorates phenotypes linked to age-related macular degeneration. Mol. Ther. 2021, 29, 2281–2293. [Google Scholar] [CrossRef]
- Zhao, W.S.; Xu, L.; Wang, X.; Li, X.L.; Yang, X.C. Endothelium-specific endothelin-1 expression promotes pro-inflammatory macrophage activation by regulating miR-33/NR4A axis. Exp. Cell. Res. 2021, 399, 112443. [Google Scholar]
- Moradi-Chaleshtori, M.; Bandehpour, M.; Heidari, N.; Mohammadi-Yeganeh, S.; Hashemi, S.M. Exosome-mediated miR-33 transfer induces M1 polarization in mouse macrophages and exerts antitumor effect in 4T1 breast cancer cell line. Int. Immunopharmacol. 2021, 90, 107198. [Google Scholar] [CrossRef] [PubMed]
- Ke, F.C.; Chuang, L.C.; Lee, M.T.; Chen, Y.J.; Lin, S.W.; Wang, S.P.; Stocco, D.M.; Hwang, J. The modulatory role of transforming growth factor beta1 and androstenedione on follicle-stimulating hormone-induced gelatinase secretion and steroidogenesis in rat granulosa cells. Biol. Reprod. 2004, 70, 1292–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Boerboom, D.; Carriere, P.D. Transforming growth factor-beta1 inhibits luteinization and promotes apoptosis in bovine granulosa cells. Reproduction 2009, 137, 969–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, X.; Wang, L.F.; Li, Q.Q.; Wu, W.J.; Shang, P.; Chamba, Y.; Pan, Z.; Li, Q. miR-130a/TGF-b1 axis is involved in sow fertility by controlling granulosa cell apoptosis. Theriogenology 2020, 157, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Kaufman, R.J. From endoplasmic-reticulum stress to the inflammatory response. Nature 2008, 454, 455–462. [Google Scholar] [CrossRef] [Green Version]
- Kuo, S.W.; Ke, F.C.; Chang, G.D.; Lee, M.T.; Hwang, J.J. Potential role of follicle-stimulating hormone (FSH) and transforming growth factor (TGFbeta1) in the regulation of ovarian angiogenesis. J. Cell. Physiol. 2011, 226, 1608–1619. [Google Scholar] [CrossRef] [PubMed]
- Nagahama, Y.; Yamashita, M. Regulation of oocyte maturation in fish. Dev. Growth Differ. 2008, 50, S195–S219. [Google Scholar] [CrossRef]
- Liu, N.N.; Cheng, X.K.; Yang, Y.; Liu, J.; Gu, H.R.; Wang, Z.J. Effects of low temperature on the ovarian development of Sinibrama taeniatus. J. Fish. China 2021. Available online: https://kns.cnki.net/kcms/detail/31.1283.S.20210415.1036.004.html (accessed on 15 April 2021).
- Nichols, J.A.; Perego, M.C.; Schütz, L.F.; Hemple, M.A.; Spicer, J.L. Hormonal regulation of vascular endothelial growth factor A (VEGFA) gene expression in granulosa and theca cells of cattle. J. Anim. Sci. 2019, 97, 3034–3045. [Google Scholar] [CrossRef]
- Irusta, G.; Abramovich, D.; Parborell, F.; Tesone, M. Direct survival role of vascular endothelial growth factor (VEGF) on rat ovarian follicular cells. Mol. Cell. Endocrinol. 2010, 325, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Bao, R.Q.; Peng, S.W.; Zhang, C.P. The molecular mechanism of ovarian granulosa cell tumors. J. Ovarian Res. 2018, 11, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, L. Overexpression of VEGF-A Isoform-VEGF164 in Mouse Ovarian Granulosa Cells by Lentiviral Vector-Mediated; Nanjing Agricultural University: Nanjing, China, 2012; pp. 55–68. [Google Scholar]
- Kosaka, N.; Sudo, N.; Miyamoto, A.; Shimizu, T. Vascular endothelial growth factor (VEGF) suppresses ovarian granulosa cell apoptosis in vitro. Biochem. Biophys. Res. Commun. 2007, 363, 733–737. [Google Scholar] [CrossRef] [PubMed]
- Quintana, R.; Kopcow, L.; Sueldo, C.; Guillermo, M.; Nidia, G.R.; Rosa, I.B. Direct injection of vascular endothelial growth factor into the ovary of mice promotes follicular development. Fertil. Steril. 2004, 82, 1101–1105. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurement | ||
|---|---|---|
| Sample at 6 d | Control group | Heat-stressed group |
| Gonadal weight (g) | 8.76 ± 1.23 b | 10.56 ± 1.79 a |
| The number of atretic oocytes | 48–69 | 72–108 |
| Mean of atretic follicles | 58 b | 90 a |
| Sample at 12 d | ||
| Gonadal weight (g) | 12.05 ± 2.21 a | 6.11 ± 1.84 b |
| The number of atretic oocytes | 44–78 | 84–122 |
| Mean of atretic follicles | 61 b | 103 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiang, J.; Tao, F.-Y.; Lu, Q.-S.; He, J.; Xu, P. Upregulation of miR-33 Exacerbates Heat-Stress-Induced Apoptosis in Granulosa Cell and Follicular Atresia of Nile Tilapia (Oreochromis niloticus) by Targeting TGFβ1I1. Genes 2022, 13, 1009. https://doi.org/10.3390/genes13061009
Qiang J, Tao F-Y, Lu Q-S, He J, Xu P. Upregulation of miR-33 Exacerbates Heat-Stress-Induced Apoptosis in Granulosa Cell and Follicular Atresia of Nile Tilapia (Oreochromis niloticus) by Targeting TGFβ1I1. Genes. 2022; 13(6):1009. https://doi.org/10.3390/genes13061009
Chicago/Turabian StyleQiang, Jun, Fan-Yi Tao, Qi-Si Lu, Jie He, and Pao Xu. 2022. "Upregulation of miR-33 Exacerbates Heat-Stress-Induced Apoptosis in Granulosa Cell and Follicular Atresia of Nile Tilapia (Oreochromis niloticus) by Targeting TGFβ1I1" Genes 13, no. 6: 1009. https://doi.org/10.3390/genes13061009