
Suicide and Changes in Expression of Neuronal miRNA Predicted by an Algorithm Search through miRNA Databases


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Subjects
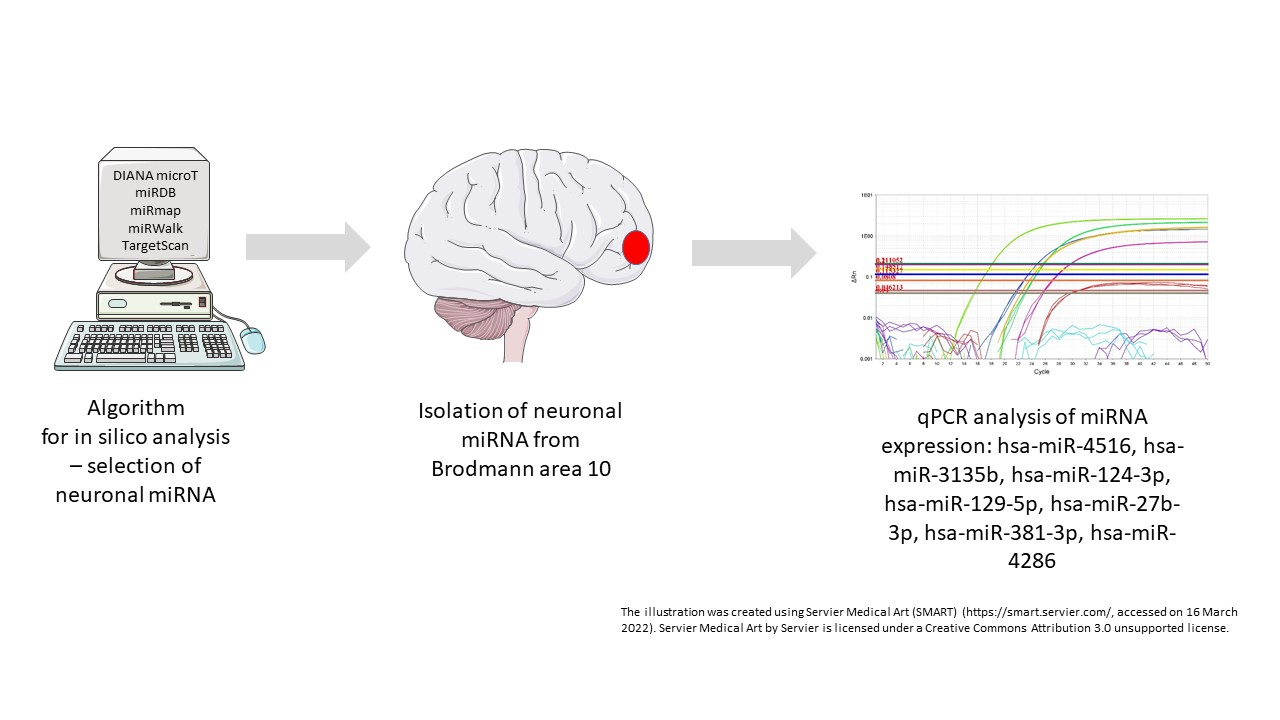
2.2. Algorithm Design and in Silico Selection of miRNAs
2.3. Isolation and Gene Expression of miRNA and Target Gene mRNA
2.4. Statistical Analysis
3. Results
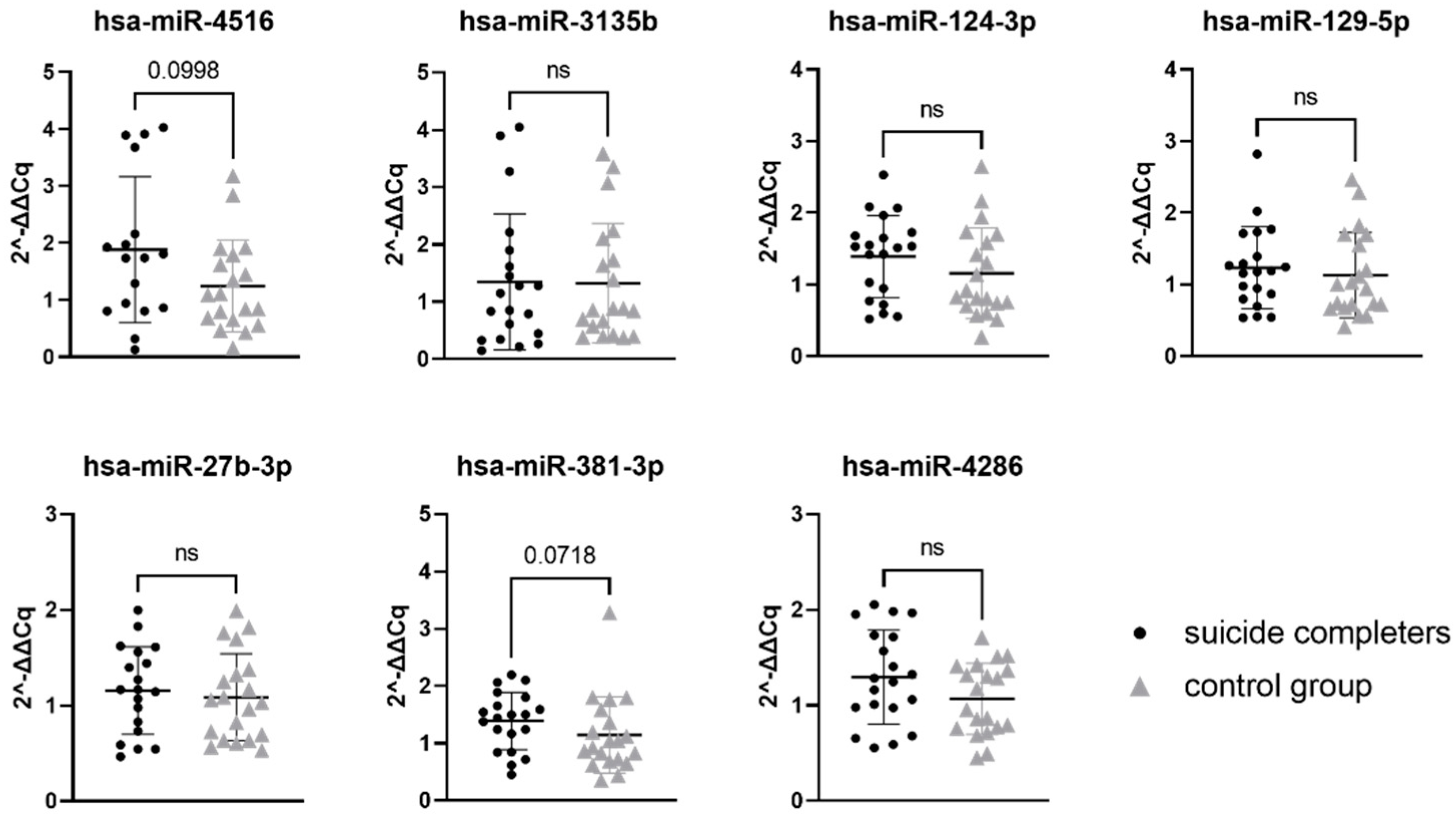
3.1. MiRNA Gene Expression
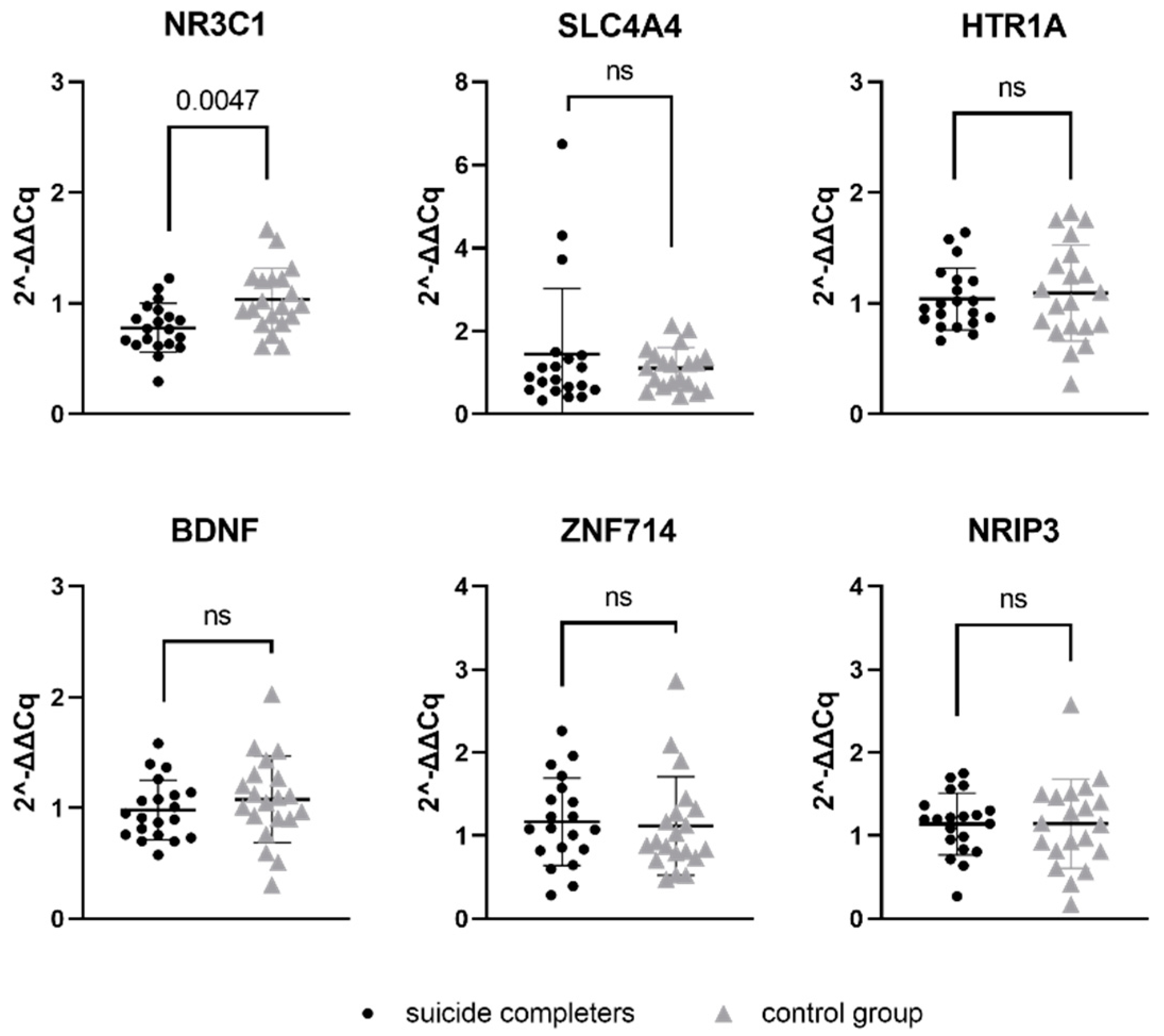
3.2. Target Gene mRNA Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Suicide in the World: Global Health Estimates; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- World Health Organization. Live Life: An Implementation Guide for Suicide Prevention in Countries; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Turecki, G.; Brent, D.A.; Gunnell, D.; O’Connor, R.C.; Oquendo, M.A.; Pirkis, J.; Stanley, B.H. Suicide and suicide risk. Nat. Rev. Dis. Primers 2019, 5, 74. [Google Scholar] [CrossRef] [PubMed]
- Bondy, B.; Buettner, A.; Zill, P. Genetics of suicide. Mol. Psychiatry 2006, 11, 336–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brent, D.A.; Melhem, N. Familial transmission of suicidal behavior. Psychiatr. Clin. N. Am. 2008, 31, 157–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voracek, M. Genetic factors in suicide: Reassessment of adoption studies and individuals’ beliefs about adoption study findings. Psychiatr. Danub. 2007, 19, 139–153. [Google Scholar]
- Voracek, M.; Loibl, L.M. Genetics of suicide: A systematic review of twin studies. Wien. Klin. Wochenschr. 2007, 119, 463–475. [Google Scholar] [CrossRef]
- Fanelli, G.; Serretti, A. The influence of the serotonin transporter gene 5-httlpr polymorphism on suicidal behaviors: A meta-analysis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 88, 375–387. [Google Scholar] [CrossRef]
- Mirkovic, B.; Laurent, C.; Podlipski, M.A.; Frebourg, T.; Cohen, D.; Gerardin, P. Genetic association studies of suicidal behavior: A review of the past 10 years, progress, limitations, and future directions. Front. Psychiatry 2016, 7, 158. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.Y.; Jia, C.X.; Lian, Y.; Sun, S.H.; Lyu, M.; Wu, A. Association of the htr2a 102t/c polymorphism with attempted suicide: A meta-analysis. Psychiatr. Genet. 2015, 25, 168–177. [Google Scholar] [CrossRef]
- Martinowich, K.; Lu, B. Interaction between bdnf and serotonin: Role in mood disorders. Neuropsychopharmacology 2008, 33, 73–83. [Google Scholar] [CrossRef]
- Popova, N.K.; Naumenko, V.S. Neuronal and behavioral plasticity: The role of serotonin and bdnf systems tandem. Expert Opin. Ther. Targets 2019, 23, 227–239. [Google Scholar] [CrossRef]
- González-Castro, T.B.; Salas-Magaña, M.; Juárez-Rojop, I.E.; López-Narváez, M.L.; Tovilla-Zárate, C.A.; Hernández-Díaz, Y. Exploring the association between bdnf val66met polymorphism and suicidal behavior: Meta-analysis and systematic review. J. Psychiatr. Res. 2017, 94, 208–217. [Google Scholar] [CrossRef] [PubMed]
- González-Castro, T.B.; Tovilla-Zárate, C.A.; Genis-Mendoza, A.D.; Juárez-Rojop, I.E.; Nicolini, H.; López-Narváez, M.L.; Martínez-Magaña, J.J. Identification of gene ontology and pathways implicated in suicide behavior: Systematic review and enrichment analysis of gwas studies. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2019, 180, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Cheung, S.; Woo, J.; Maes, M.S.; Zai, C.C. Suicide epigenetics, a review of recent progress. J. Affect. Disord. 2020, 265, 423–438. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, Y. Micrornas in depression and suicide: Recent insights and future perspectives. J. Affect. Disord. 2018, 240, 146–154. [Google Scholar] [CrossRef]
- McGowan, P.O.; Sasaki, A.; D’Alessio, A.C.; Dymov, S.; Labonte, B.; Szyf, M.; Turecki, G.; Meaney, M.J. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat. Neurosci. 2009, 12, 342–348. [Google Scholar] [CrossRef] [Green Version]
- Alles, J.; Fehlmann, T.; Fischer, U.; Backes, C.; Galata, V.; Minet, M.; Hart, M.; Abu-Halima, M.; Grässer, F.A.; Lenhof, H.-P.; et al. An estimate of the total number of true human mirnas. Nucleic Acids Res. 2019, 47, 3353–3364. [Google Scholar] [CrossRef] [Green Version]
- Slota, J.A.; Booth, S.A. Micrornas in neuroinflammation: Implications in disease pathogenesis, biomarker discovery and therapeutic applications. Noncoding RNA 2019, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- Saçar, M.D.; Allmer, J. Data mining for microrna gene prediction: On the impact of class imbalance and feature number for microrna gene prediction. In Proceedings of the 8th International Symposium on Health Informatics and Bioinformatics, Ankara, Turkey, 25–27 September 2013; pp. 1–6. [Google Scholar]
- Vishnoi, A.; Rani, S. Mirna biogenesis and regulation of diseases: An overview. Methods Mol. Biol. 2017, 1509, 1–10. [Google Scholar]
- Gantier, M.P.; McCoy, C.E.; Rusinova, I.; Saulep, D.; Wang, D.; Xu, D.; Irving, A.T.; Behlke, M.A.; Hertzog, P.J.; Mackay, F.; et al. Analysis of microrna turnover in mammalian cells following dicer1 ablation. Nucleic Acids Res. 2011, 39, 5692–5703. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. Metazoan micrornas. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef] [Green Version]
- Griffiths-Jones, S.; Grocock, R.J.; van Dongen, S.; Bateman, A.; Enright, A.J. Mirbase: Microrna sequences, targets and gene nomenclature. Nucleic Acids Res. 2006, 34, D140–D144. [Google Scholar] [CrossRef] [PubMed]
- Conaco, C.; Otto, S.; Han, J.J.; Mandel, G. Reciprocal actions of rest and a microrna promote neuronal identity. Proc. Natl. Acad. Sci. USA 2006, 103, 2422–2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geaghan, M.; Cairns, M.J. Microrna and posttranscriptional dysregulation in psychiatry. Biol. Psychiatry 2015, 78, 231–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruzdev, S.K.; Yakovlev, A.A.; Druzhkova, T.A.; Guekht, A.B.; Gulyaeva, N.V. The missing link: How exosomes and mirnas can help in bridging psychiatry and molecular biology in the context of depression, bipolar disorder and schizophrenia. Cell. Mol. Neurobiol. 2019, 39, 729–750. [Google Scholar] [CrossRef]
- van den Berg, M.M.J.; Krauskopf, J.; Ramaekers, J.G.; Kleinjans, J.C.S.; Prickaerts, J.; Briedé, J.J. Circulating micrornas as potential biomarkers for psychiatric and neurodegenerative disorders. Prog. Neurobiol. 2020, 185, 101732. [Google Scholar] [CrossRef]
- Paraskevopoulou, M.D.; Georgakilas, G.; Kostoulas, N.; Vlachos, I.S.; Vergoulis, T.; Reczko, M.; Filippidis, C.; Dalamagas, T.; Hatzigeorgiou, A.G. Diana-microt web server v5.0: Service integration into mirna functional analysis workflows. Nucleic Acids Res. 2013, 41, W169–W173. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, X. Mirdb: An online database for prediction of functional microrna targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef] [Green Version]
- Vejnar, C.E.; Zdobnov, E.M. Mirmap: Comprehensive prediction of microrna target repression strength. Nucleic Acids Res. 2012, 40, 11673–11683. [Google Scholar] [CrossRef]
- Sticht, C.; De La Torre, C.; Parveen, A.; Gretz, N. Mirwalk: An online resource for prediction of microrna binding sites. PLoS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microrna target sites in mammalian mrnas. Elife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Lopez, J.P.; Fiori, L.M.; Gross, J.A.; Labonte, B.; Yerko, V.; Mechawar, N.; Turecki, G. Regulatory role of mirnas in polyamine gene expression in the prefrontal cortex of depressed suicide completers. Int. J. Neuropsychopharmacol. 2014, 17, 23–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, N.; Leidinger, P.; Becker, K.; Backes, C.; Fehlmann, T.; Pallasch, C.; Rheinheimer, S.; Meder, B.; Stähler, C.; Meese, E.; et al. Distribution of mirna expression across human tissues. Nucleic Acids Res. 2016, 44, 3865–3877. [Google Scholar] [CrossRef] [PubMed]
- Kouter, K.; Zupanc, T.; Videtič Paska, A. Genome-wide DNA methylation in suicide victims revealing impact on gene expression. J. Affect. Disord. 2019, 253, 419–425. [Google Scholar] [CrossRef]
- Schmaal, L.; van Harmelen, A.-L.; Chatzi, V.; Lippard, E.T.C.; Toenders, Y.J.; Averill, L.A.; Mazure, C.M.; Blumberg, H.P. Imaging suicidal thoughts and behaviors: A comprehensive review of 2 decades of neuroimaging studies. Mol. Psychiatry 2020, 25, 408–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The miqe guidelines: Minimum information for publication of quantitative real-time pcr experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2−ΔΔc(t) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Fries, G.R.; Lima, C.N.C.; Valvassori, S.S.; Zunta-Soares, G.; Soares, J.C.; Quevedo, J. Preliminary investigation of peripheral extracellular vesicles’ micrornas in bipolar disorder. J. Affect. Disord. 2019, 255, 10–14. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, Y.; Teng, P.; Yang, Q. Differential expression of mir-381-3p in alzheimer’s disease patients and its role in beta-amyloid-induced neurotoxicity and inflammation. Neuroimmunomodulation 2021. [Google Scholar] [CrossRef]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. G:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [Green Version]
- Angelopoulou, E.; Paudel, Y.N.; Piperi, C. Mir-124 and parkinson’s disease: A biomarker with therapeutic potential. Pharmacol. Res. 2019, 150, 104515. [Google Scholar] [CrossRef]
- Serafini, G.; Pompili, M.; Innamorati, M.; Giordano, G.; Montebovi, F.; Sher, L.; Dwivedi, Y.; Girardi, P. The role of micrornas in synaptic plasticity, major affective disorders and suicidal behavior. Neurosci. Res. 2012, 73, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.; Mu, R.H.; Li, C.F.; Dong, S.Q.; Geng, D.; Liu, Q.; Yi, L.T. Microrna-124 targets glucocorticoid receptor and is involved in depression-like behaviors. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 79, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.; Dunbar, M.; Shelton, R.C.; Dwivedi, Y. Identification of microrna-124-3p as a putative epigenetic signature of major depressive disorder. Neuropsychopharmacology 2017, 42, 864–875. [Google Scholar] [CrossRef] [Green Version]
- Kadmiel, M.; Cidlowski, J.A. Glucocorticoid receptor signaling in health and disease. Trends Pharmacol. Sci. 2013, 34, 518–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, I.C.; Cervoni, N.; Champagne, F.A.; D’Alessio, A.C.; Sharma, S.; Seckl, J.R.; Dymov, S.; Szyf, M.; Meaney, M.J. Epigenetic programming by maternal behavior. Nat. Neurosci. 2004, 7, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.; Shelton, R.C.; Dwivedi, Y. DNA methylation and expression of stress related genes in pbmc of mdd patients with and without serious suicidal ideation. J. Psychiatr. Res. 2017, 89, 115–124. [Google Scholar] [CrossRef]
- Wang, H.; Gou, X.; Jiang, T.; Ouyang, J. The effects of micrornas on glucocorticoid responsiveness. J. Cancer Res. Clin. Oncol. 2017, 143, 1005–1011. [Google Scholar] [CrossRef]
- Labonté, B.; Suderman, M.; Maussion, G.; Navaro, L.; Yerko, V.; Mahar, I.; Bureau, A.; Mechawar, N.; Szyf, M.; Meaney, M.J.; et al. Genome-wide epigenetic regulation by early-life trauma. Arch. Gen. Psychiatry 2012, 69, 722–731. [Google Scholar] [CrossRef]
- Hernández-Díaz, Y.; Genis-Mendoza, A.D.; González-Castro, T.B.; Tovilla-Zárate, C.A.; Juárez-Rojop, I.E.; López-Narváez, M.L.; Nicolini, H. Association and genetic expression between genes involved in hpa axis and suicide behavior: A systematic review. Genes 2021, 12, 1608. [Google Scholar] [CrossRef]
- Weber, M.J. New human and mouse microrna genes found by homology search. FEBS J. 2005, 272, 59–73. [Google Scholar] [CrossRef]
- Roy, B.; Yoshino, Y.; Allen, L.; Prall, K.; Schell, G.; Dwivedi, Y. Exploiting circulating micrornas as biomarkers in psychiatric disorders. Mol. Diagn. Ther. 2020, 24, 279–298. [Google Scholar] [CrossRef] [PubMed]
- Serafini, G.; Pompili, M.; Hansen, K.; Obrietan, K.; Dwivedi, Y.; Amore, M.; Shomron, N.; Girardi, P. Micrornas: Fundamental regulators of gene expression in major affective disorders and suicidal behavior? Front. Cell. Neurosci. 2013, 7, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yáñez-Mó, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Suicide Victims | Control Group | p-Value | |
|---|---|---|---|
| Age (years + SD) | 44.6 ± 10.4 | 54.5 ± 7.4 | 0.0016, t = 3.391, df = 38 |
| PMI (hours + SD) | 27.5 ± 14.0 | 27.1 ± 21.6 | 0.9430, t = 0.07202, df = 38 |
| RIN | 7.7 ± 0.22 | 7.26 ± 0.28 | 0.0794, t = 2.557, df = 8 |
| Database | BDNF | HTR1A | SLC6A4 | NR3C1 | ZNF714 | NRIP3 |
|---|---|---|---|---|---|---|
| miRWalk | 2353 | 1462 | 1942 | 2349 | 1858 | 1916 |
| miRmap | 979 | 48 | 1300 | 370 | 65 | 1202 |
| TargetScan | 775 | 153 | 1115 | 976 | 1506 | 875 |
| DIANA microT-CDS | 272 | 1 | 201 | 301 | 303 | 211 |
| miRDB | 191 | 21 | 161 | 472 | 269 | 163 |
| Selected miRNAs | Target Interaction Analysis | Expression Analysis | Weighted Sum |
|---|---|---|---|
| hsa-miR-4516 | 13.63 | 17.17 | 30.80 |
| hsa-miR-3135b | 16.75 | 13.57 | 30.33 |
| hsa-miR-124-3p | 11.55 | 17.87 | 29.43 |
| hsa-miR-129-5p | 20.89 | 8.47 | 29.35 |
| hsa-miR-27b-3p | 15.80 | 12.34 | 28.14 |
| hsa-miR-381-3p | 20.01 | 7.48 | 27.49 |
| hsa-miR-4286 | 9.37 | 18.06 | 27.43 |
| Selected miRNAs | Suicide Completers Median | Control Group Median | U Test Statistic | p-Value |
|---|---|---|---|---|
| hsa-miR-4516 | 1.738 | 1.092 | 109 | 0.0998 |
| hsa-miR-3135b | 1.002 | 0.8698 | 187 | 0.7381 |
| hsa-miR-124-3p | 1.520 | 0.8713 | 157 | 0.2534 |
| hsa-miR-129-5p | 1.184 | 0.9738 | 170 | 0.4291 |
| hsa-miR-27b-3p | 1.167 | 1.044 | 171 | 0.6071 |
| hsa-miR-381-3p | 1.468 | 0.9845 | 133 | 0.0718 |
| hsa-miR-4286 | 1.266 | 1.065 | 152 | 0.2012 |
| mRNAs | Suicide Completers Median | Control Group Median | U Test Statistic | p-Value |
|---|---|---|---|---|
| NR3C1 | 0.7685 | 0.9780 | 97 | 0.0047 |
| SLC6A4 | 0.8601 | 1.163 | 179 | 0.5831 |
| HTR1A | 0.9735 | 1.054 | 188 | 0.7584 |
| BDNF | 0.9309 | 1.071 | 161 | 0.3013 |
| ZNF714 | 1.085 | 0.9057 | 177 | 0.5468 |
| NRIP3 | 1.194 | 1.143 | 192 | 0.8410 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Videtič Paska, A.; Alič, U.; Zupanc, T.; Kouter, K. Suicide and Changes in Expression of Neuronal miRNA Predicted by an Algorithm Search through miRNA Databases. Genes 2022, 13, 562. https://doi.org/10.3390/genes13040562
Videtič Paska A, Alič U, Zupanc T, Kouter K. Suicide and Changes in Expression of Neuronal miRNA Predicted by an Algorithm Search through miRNA Databases. Genes. 2022; 13(4):562. https://doi.org/10.3390/genes13040562
Chicago/Turabian StyleVidetič Paska, Alja, Urban Alič, Tomaž Zupanc, and Katarina Kouter. 2022. "Suicide and Changes in Expression of Neuronal miRNA Predicted by an Algorithm Search through miRNA Databases" Genes 13, no. 4: 562. https://doi.org/10.3390/genes13040562