
Genomic Analysis of Resistance to Fall Armyworm (Spodoptera frugiperda) in CIMMYT Maize Lines

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Artificial Infestation and Phenotypic Evaluation
2.3. Phenotypic and Genotypic Data Analyses
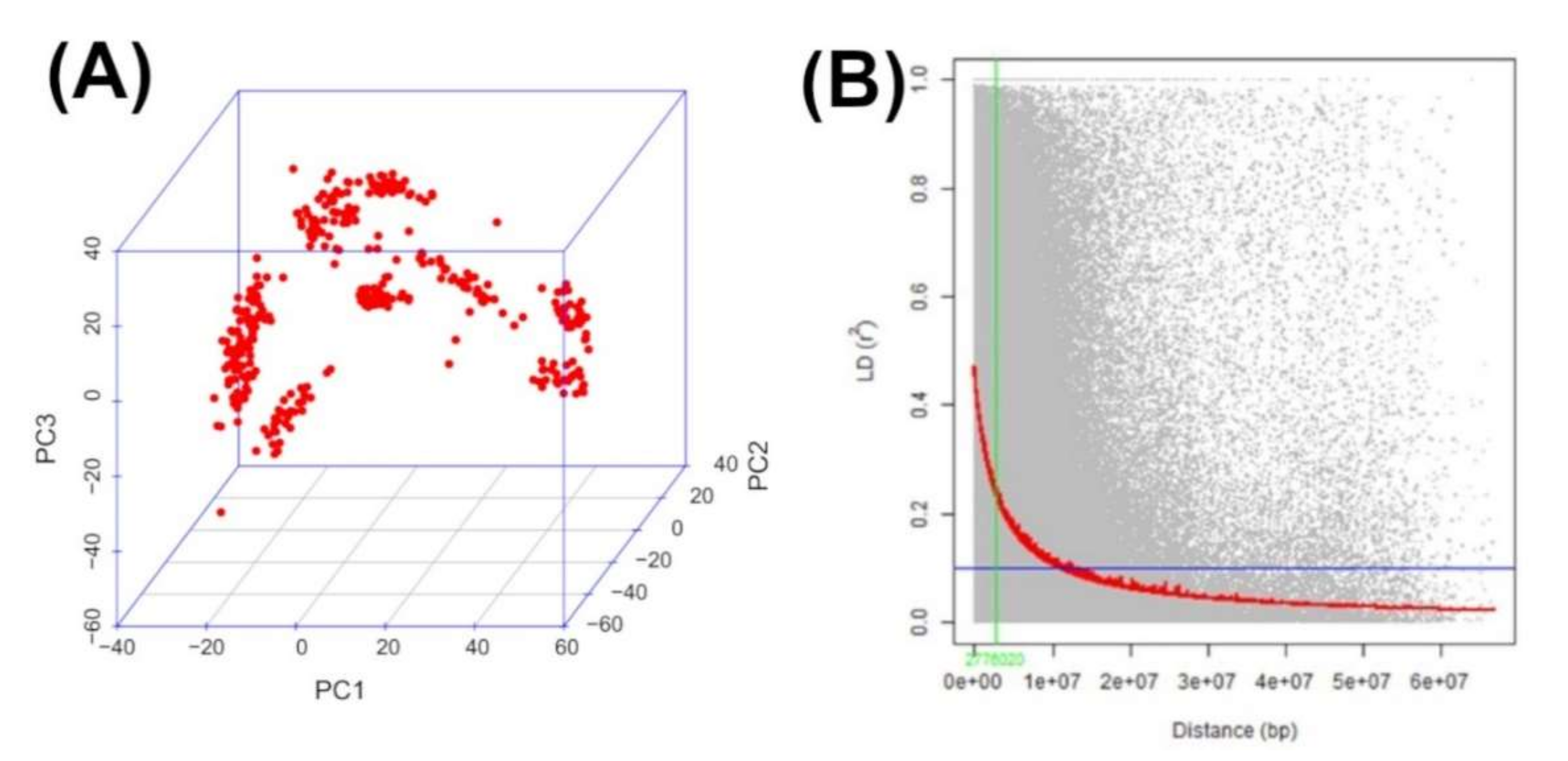
2.4. Population Structure, Kinship, Linkage Disequilibrium, and GWAS
3. Results
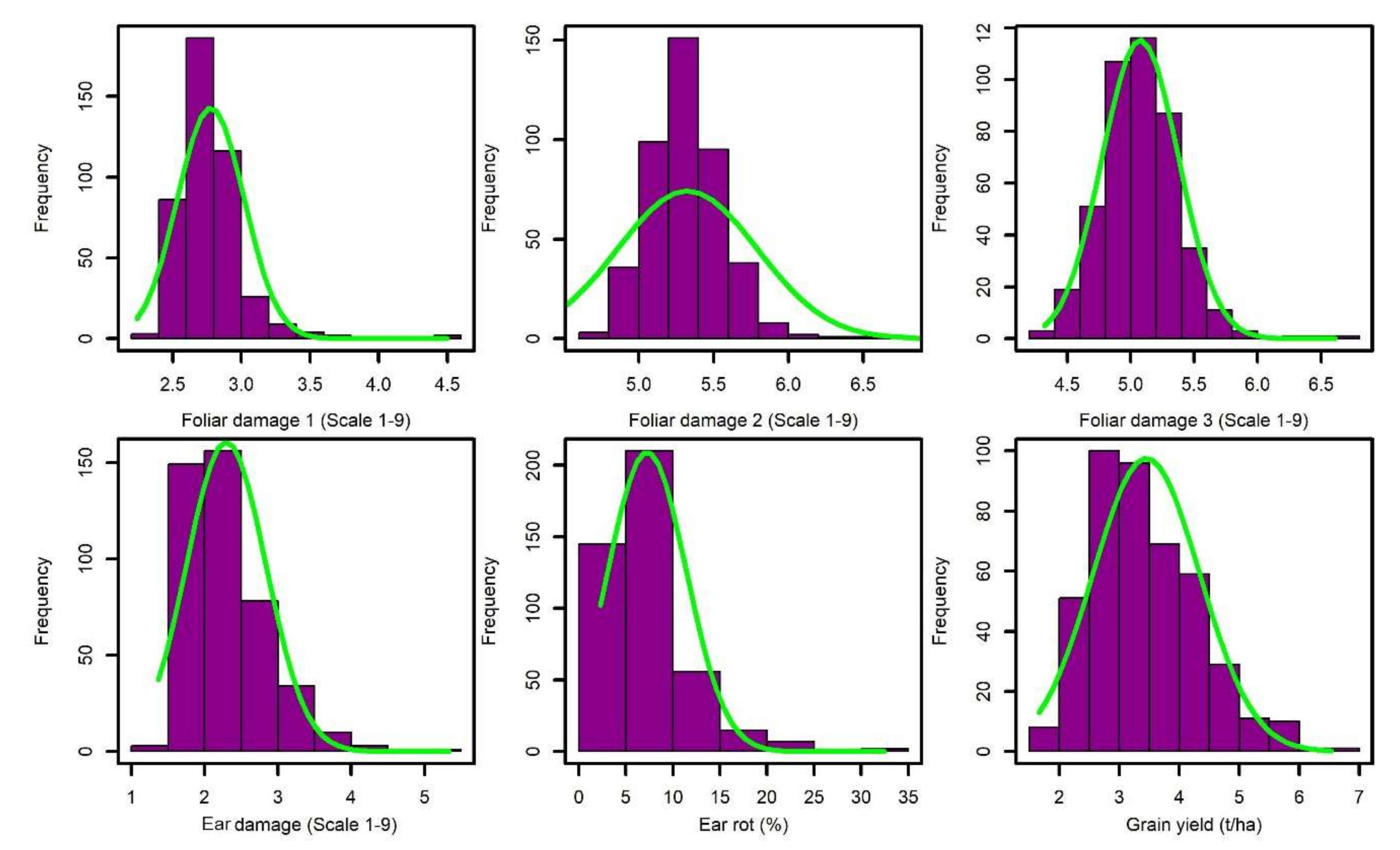
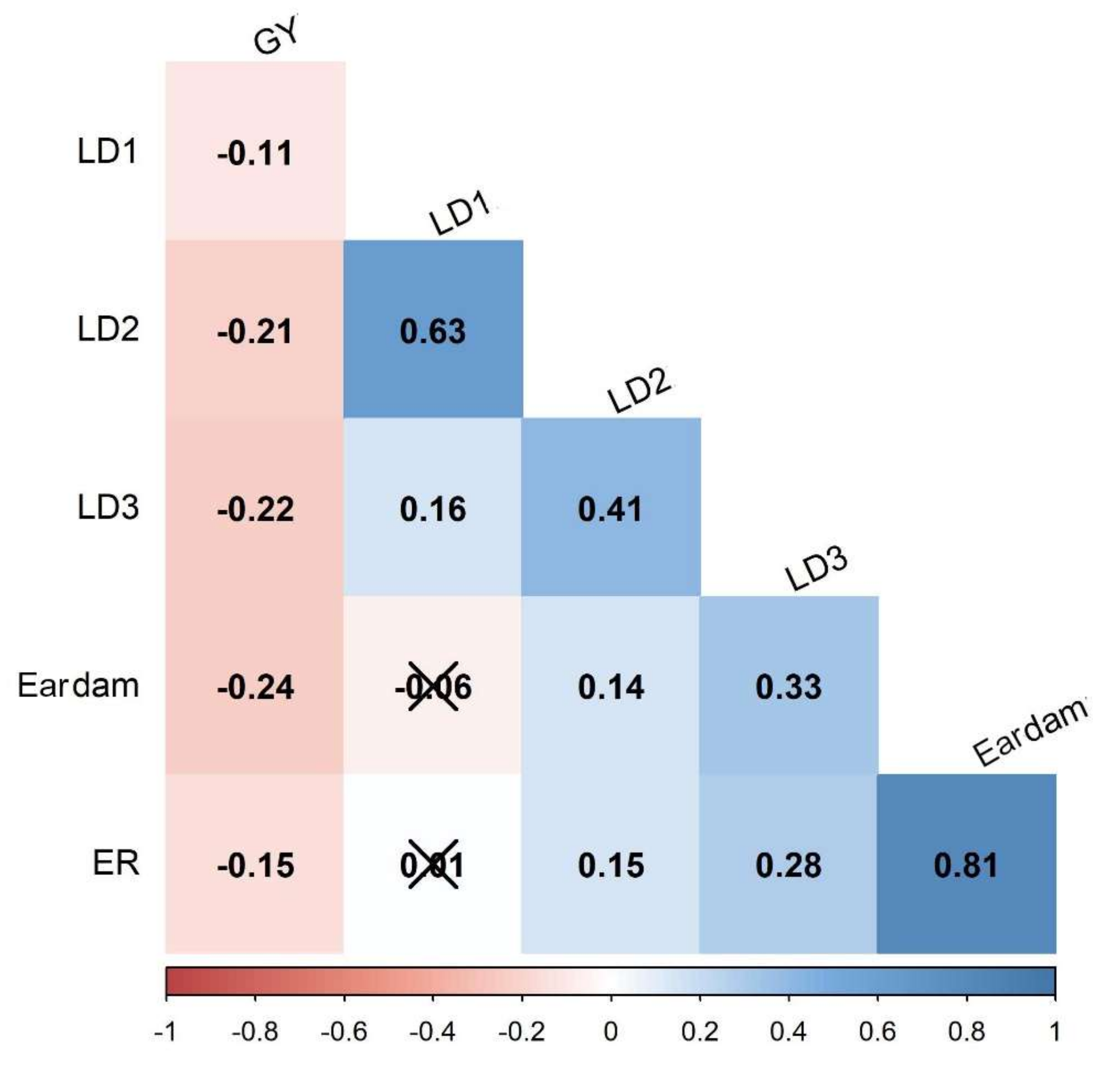
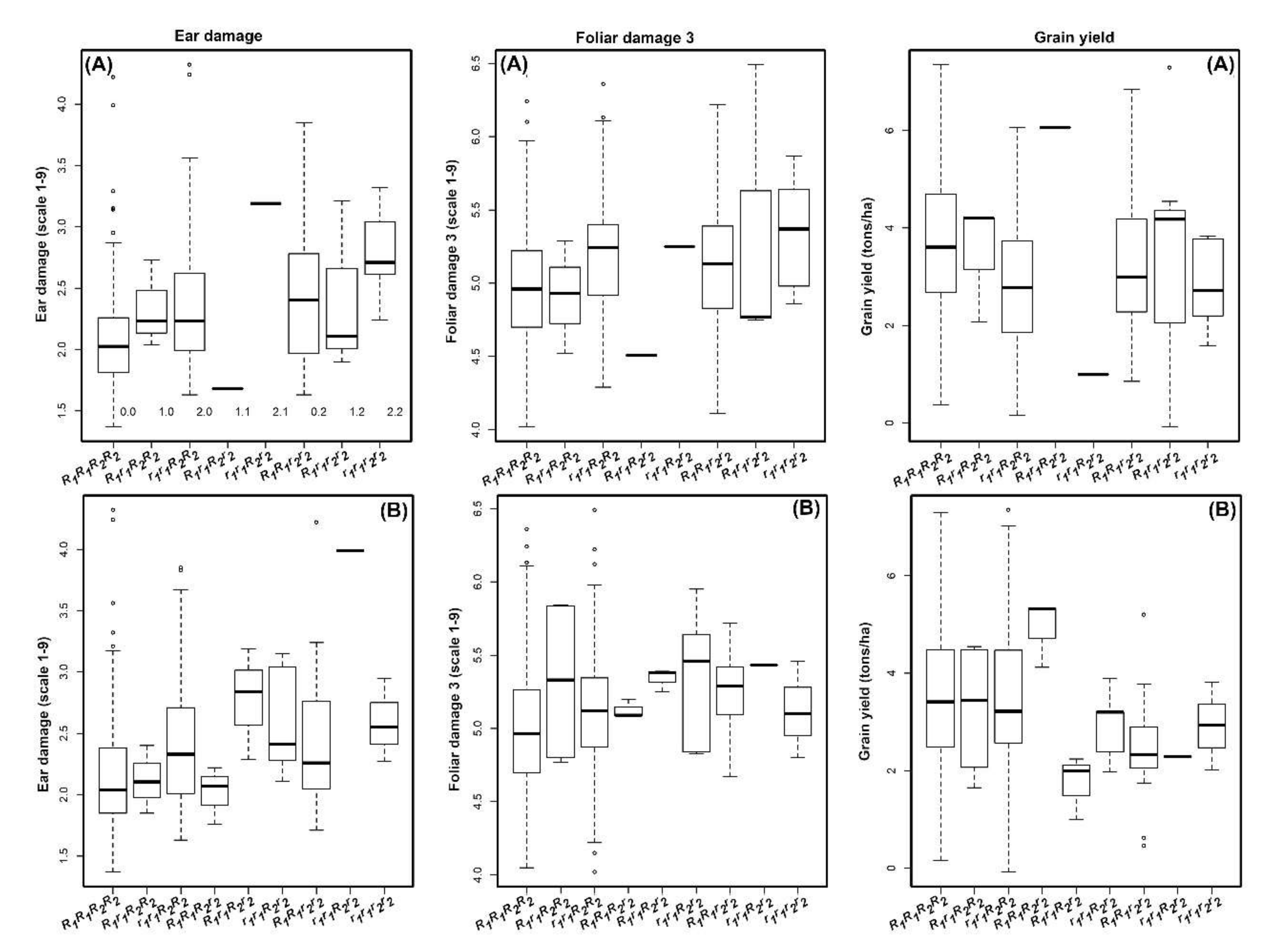
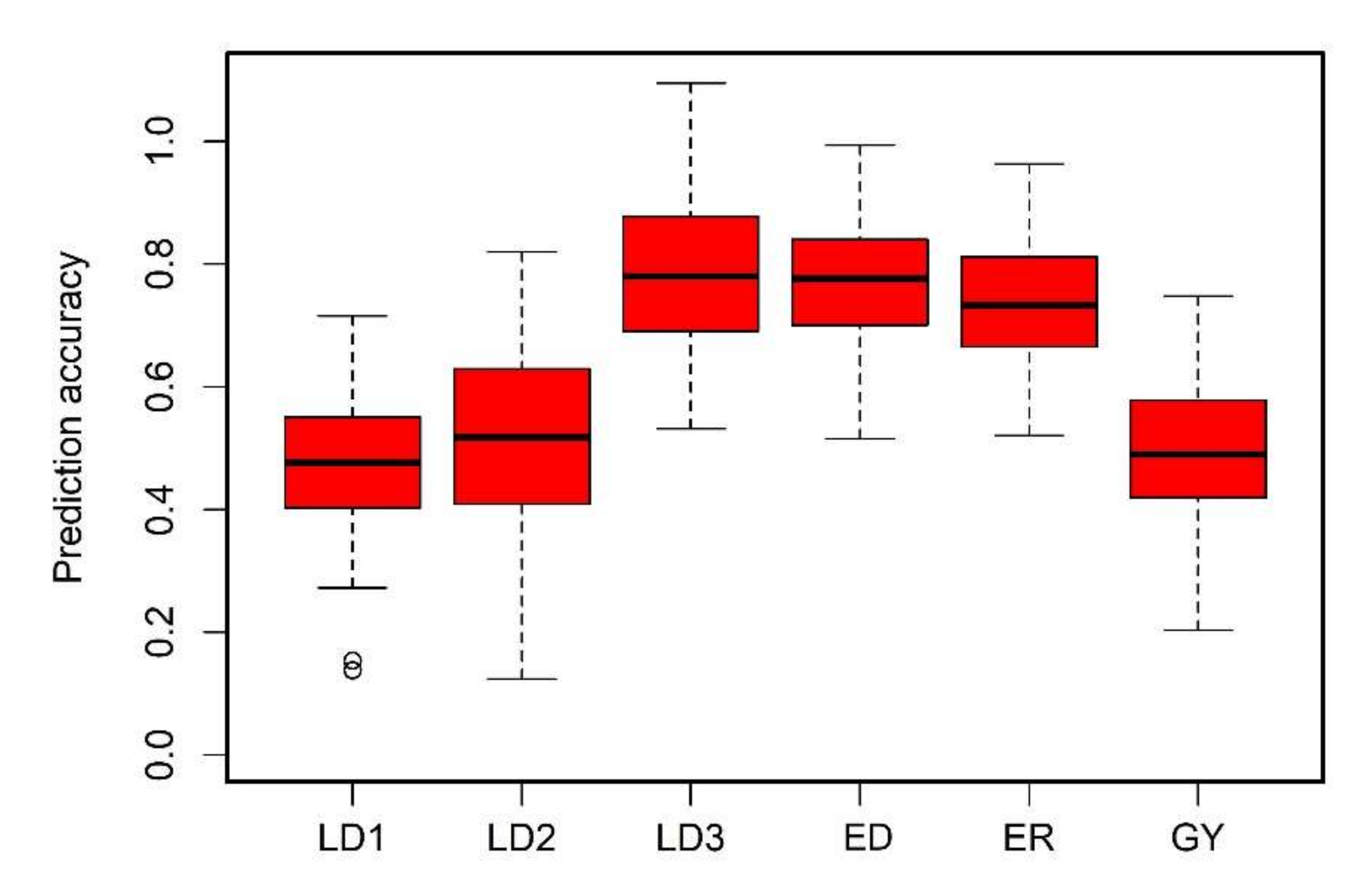
3.1. Phenotypic Data Analyses
3.2. PCA, Kinship and LD Analysis
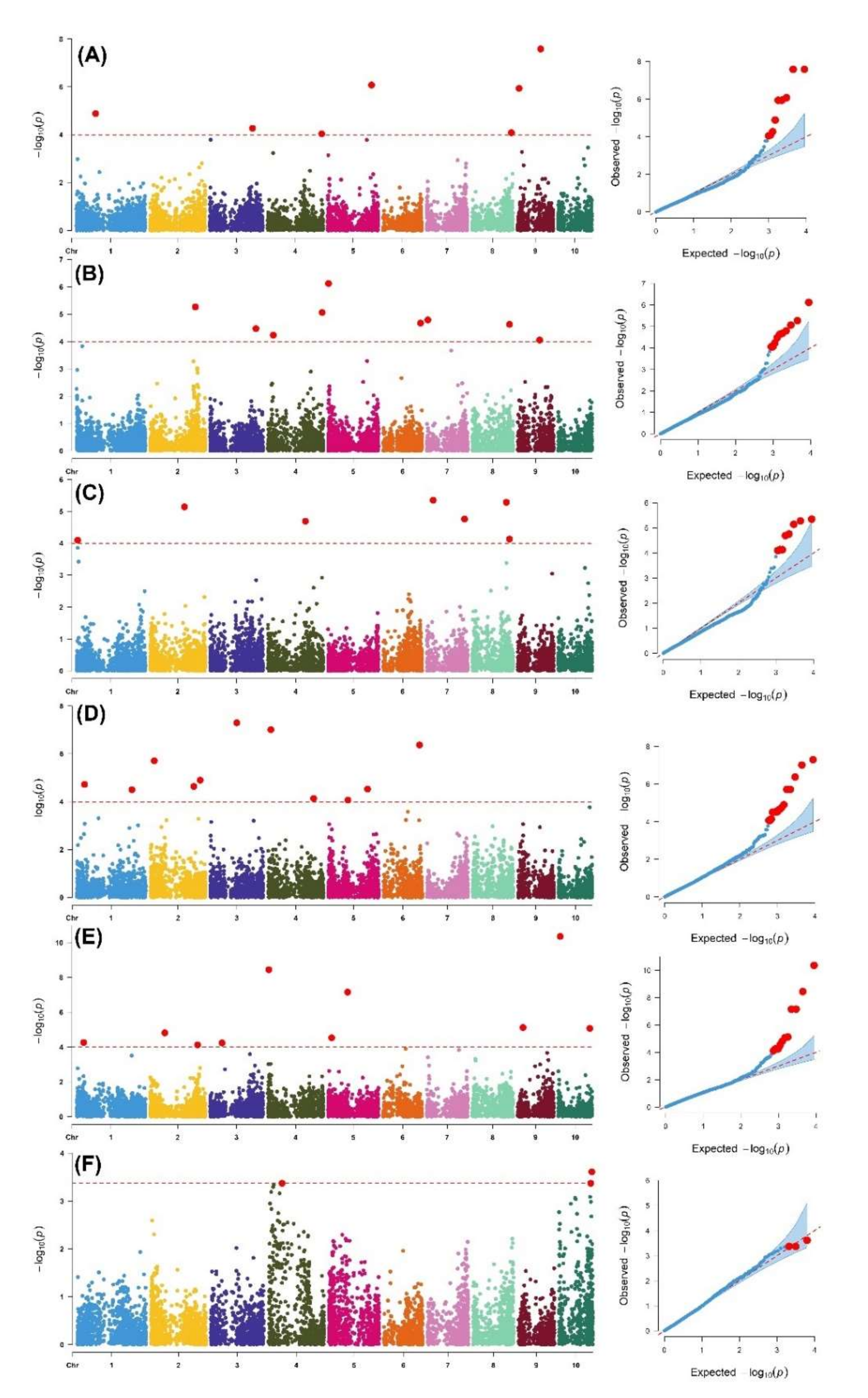
3.3. Genome-Wide Association Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shiferaw, B.; Prasanna, B.M.; Hellin, J.; Bänziger, M. Crops That Feed the World 6. Past Successes and Future Challenges to the Role Played by Maize in Global Food Security. Food Secur. 2011, 3, 307–327. [Google Scholar] [CrossRef] [Green Version]
- Santpoort, R. The Drivers of Maize Area Expansion in Sub-Saharan Africa. How Policies to Boost Maize Production Overlook the Interests of Smallholder Farmers. Land 2020, 9, 68. [Google Scholar] [CrossRef] [Green Version]
- Girvetz, E.; Ramirez-Villegas, J.; Claessens, L.; Lamanna, C.; Navarro-Racines, C.; Nowak, A.; Thornton, P.; Rosenstock, T.S. Future Climate Projections in Africa: Where Are We Headed? In The Climate-Smart Agriculture Papers; Springer: Cham, Switzerland, 2019; pp. 15–27. [Google Scholar]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First Report of Outbreaks of the Fall Armyworm Spodoptera Frugiperda (J.E. Smith) (Lepidoptera, Noctuidae), a New Alien Invasive Pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [Green Version]
- Prasanna, B.M.; Huesing, J.E.; Eddy, R.; Peschke, V.M.; Regina, E.; Virginia, M.P. Fall Armyworm in Africa: A Guide for Integrated Pest Management, 1st ed.; Prasanna, B.M., Regina, E., Virginia, M.P., Eds.; CIMMYT: Queretaro, Mexico, 2018. [Google Scholar]
- De Groote, H.; Kimenju, S.C.; Munyua, B.; Palmas, S.; Kassie, M.; Bruce, A. Spread, and impact of fall armyworm (Spodoptera frugiperda J.E. Smith) in maize production areas of Kenya. Agriculture, Ecosystems & Environment 2020, 292, 106804. [Google Scholar] [CrossRef]
- Ni, A.; David, G.; Steve, L. Physiological Basis of Fall Armyworm (Lepidoptera: Noctuidae) Resistance in Seedlings of Maize Inbred Lines with Varying Levels of Silk Maysin. Florida Entomol. 2008, 91, 537–545. [Google Scholar] [CrossRef]
- Farhan, Y.; Smith, J.L.; Limay-Rios, V.; Schaafsma, A.W. The Effect of Simulated Lepidopteran Ear Feeding Injury on Mycotoxin Accumulation in Grain Corn (Poales: Poaceae). J. Econ. Entomol. 2020, 113, 2187–2196. [Google Scholar] [CrossRef] [PubMed]
- Oerke, E.C. Crop Losses to Pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Schlum, K.; Lamour, K.; Tandy, P.; Emrich, S.J.; de Bortoli, C.P.; Rao, T.; Viteri Dillon, D.M.; Linares-Ramirez, A.M.; Jurat-Fuentes, J.L. Genetic Screening to Identify Candidate Resistance Alleles to Cry1F Corn in Fall Armyworm Using Targeted Sequencing. Insects 2021, 12, 618. [Google Scholar] [CrossRef]
- Overton, K.; Maino, J.L.; Day, R.; Umina, P.A.; Bett, B.; Carnovale, D.; Ekesi, S.; Meagher, R.; Reynolds, O.L. Global crop impacts, yield losses and action thresholds for fall armyworm (Spodoptera frugiperda): A review. Crop. Prot. 2021, 145, 105641. [Google Scholar] [CrossRef]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall armyworm: Impacts and implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Matova, P.M.; Kamutando, C.N.; Magorokosho, C.; Kutywayo, D.; Gutsa, F.; Labuschagne, M. Fall-armyworm invasion, control practices and resistance breeding in Sub-Saharan Africa. Crop. Sci. 2020, 60, 2951–2970. [Google Scholar] [CrossRef] [PubMed]
- Storer, N.P.; Kubiszak, M.E.; Ed King, J.; Thompson, G.D.; Santos, A.C. Status of Resistance to Bt Maize in Spodoptera Frugiperda: Lessons from Puerto Rico. J. Invertebr. Pathol. 2012, 110, 294–300. [Google Scholar] [CrossRef]
- Farrar, J.J.; Ellsworth, P.C.; Sisco, R.; Baur, M.E.; Crump, A.; Fournier, A.J.; Katie Murray, M.; Jepson, P.C.; Tarutani, C.M.; Dorschner, K.W. Assessing Compatibility of a Pesticide in an IPM Program. J. Integr. Pest Manag. 2018, 9, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Liu, B.; Zheng, W.; Liu, C.; Zhang, D.; Zhao, S.; Li, Z.; Xu, P.; Wilson, K.; Withers, A.; et al. Genetic Structure and Insecticide Resistance Characteristics of Fall Armyworm Populations Invading China. Mol. Ecol. Resour. 2020, 20, 1682–1696. [Google Scholar] [CrossRef] [PubMed]
- Hruska, A.J. Fall Armyworm (Spodoptera frugiperda) Management by Smallholders. CAB Rev. 2019, 14, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Khan, Z.R.; Midega, C.A.O.; Amudavi, D.M.; Hassanali, A.; Pickett, J.A. On-Farm Evaluation of the ‘ Push—Pull ’Technology for the Control of Stemborers and Striga Weed on Maize in Western Kenya. Field Crops Res. 2008, 106, 224–233. [Google Scholar] [CrossRef]
- Harrison, R.D.; Thierfelder, C.; Baudron, F.; Chinwada, P.; Midega, C.; Schaffner, U.; Van Den Berg, J. Agro-Ecological Options for Fall Armyworm (Spodoptera frugiperda J.E. Smith) Management: Providing Low-Cost, Smallholder Friendly Solutions to an Invasive Pest. J. Environ. Manage. 2019, 243, 318–330. [Google Scholar] [CrossRef]
- Sharma, H.C.; Ortiz, R. Host Plant Resistance to Insects: An Eco-Friendly Approach for Pest Management and Environment Conservation. J. Environ. Biol. 2002, 23, 111–135. [Google Scholar]
- Ankala, A.; Kelley, R.Y.; Rowe, D.E.; Williams, W.P.; Luthe, D.S. Plant Science Foliar Herbivory Triggers Local and long Distance Defense Responses in Maize. Plant Sci. 2013, 199–200, 103–112. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of Plant Defense against Insect Herbivores. Plant Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [Green Version]
- Howe, G.A.; Jander, G. Plant Immunity to Insect Herbivores. Annu. Rev. Plant Biol. 2008, 59, 41–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Liu, H.; Wu, L.; Warburton, M.; Yan, J. Genome-Wide Association Studies in Maize: Praise and Stargaze. Mol. Plant. 2017, 10, 359–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowda, M.; Beyene, Y.; Makumbi, D.; Semagn, K.; Olsen, M.S. Discovery and Validation of Genomic Regions Associated with Resistance to Maize Lethal Necrosis in Four Biparental Populations. Mol. Breeding 2018, 38, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crossa, J.; Pérez-Rodríguez, P.; Cuevas, J.; Montesinos-lópez, O.; Jarquín, D.; Campos, G.D.L.; Burgueño, J.; González-Camacho, J.M.; Pérez-Elizalde, S.; Beyene, Y.; et al. Genomic Selection in Plant Breeding: Methods, Models, and Perspectives. Trends Plant Sci. 2017, 22, 961–975. [Google Scholar] [CrossRef]
- Foiada, F.; Westermeier, P.; Kessel, B.; Ouzunova, M.; Wimmer, V.; Mayerhofer, W.; Presterl, T.; Dilger, M.; Kreps, R.; Eder, J.; et al. Improving Resistance to the European Corn Borer: A Comprehensive Study in Elite Maize Using QTL Mapping and Genome-Wide Prediction. Theor. Appl. Genet. 2015, 128, 875–891. [Google Scholar] [CrossRef]
- Xia, L.I.; Kang-lai, H.E.; Zhen-ying, W.; Shu-xiong, B.A.I. Quantitative Trait Loci for Asian Corn Borer Resistance in Maize Population. Agric. Sci. China 2010, 9, 77–84. [Google Scholar] [CrossRef]
- Khairallah, M.M.; Bohn, M.; Jiang, C.; Deutsch, J.A.; Jewell, D.C.; Mihm, J.A.; Melchinger, A.E.; González-De-León, D.; Hoisington, D.A. Molecular Mapping of QTL for Southwestern Corn Borer Resistance, Plant Height and Flowering in Tropical Maize. Plant Breed. 1998, 117, 309–318. [Google Scholar] [CrossRef]
- Bohn, M.; Khairallah, M.M.; González-de-León, D.; Hoisington, D.A.; Utz, H.F.; Deutsch, J.A.; Jewell, D.C.; Mihm, J.A.; Melchinger, A.E. QTL Mapping in Tropical Maize: I. Genomic Regions Affecting Leaf Feeding Resistance to Sugarcane Borer and Other Traits. Crop. Sci. 1996, 36, 1352–1361. [Google Scholar] [CrossRef] [Green Version]
- Brooks, T.D.; Shaun Bushman, B.; Paul Williams, W.; McMullen, M.D.; Buckley, P.M. Genetic Basis of Resistance to Fall Armyworm (Lepidoptera: Noctuidae) and Southwestern Corn Borer (Lepidoptera: Crambidae) Leaf-Feeding Damage in Maize. J. Econ. Entomol. 2007, 100, 1470–1475. [Google Scholar] [CrossRef]
- Badji, A.; Kwemoi, D.B.; Machida, L.; Okii, D.; Mwila, N.; Agbahoungba, S.; Kumi, F.; Ibanda, A.; Bararyenya, A.; Solemanegy, M. Genetic Basis of Maize Resistance to Multiple Insect Pests: Integrated Genome-Wide Comparative Mapping and Candidate Gene Prioritization. Genes 2020, 11, 689. [Google Scholar] [CrossRef]
- Wang, Q.; Tian, F.; Pan, Y.; Buckler, E.S.; Zhang, Z. A SUPER Powerful Method for Genome Wide Association Study. PLoS ONE 2014, 9, e107684. [Google Scholar] [CrossRef] [PubMed]
- Bentley, A.R.; Scutari, M.; Gosman, N.; Faure, S.; Bedford, F.; Howell, P.; Cockram, J.; Rose, G.A.; Barber, T.; Irigoyen, J. Applying Association Mapping and Genomic Selection to the Dissection of Key Traits in Elite European Wheat. Theor. Appl. Genet. 2014, 127, 2619–2633. [Google Scholar] [CrossRef] [PubMed]
- Kumar Joshi, R.; Nayak, S. Gene Pyramiding-A Broad Spectrum Technique for Developing Durable Stress Resistance in Crops. Biotechnol. Mol. Biol. Rev. 2010, 5, 51–60. [Google Scholar]
- Tefera, T.; Mugo, S.; Tende, R.; Likhayo, P. Mass Rearing of Stem Borers. In Maize Weevil, and Larger Grain Borer Insect Pests of Maize; CIMMYT: Nairobi, Kenya, 2010; pp. 9–27. Available online: https://repository.cimmyt.org/bitstream/handle/10883/815/94546.pdf?sequence=1&isAllowed=y (accessed on 14 December 2021).
- Abendroth, L.J.; Elmore, R.W.; Boyer, M.J.; Marlay, S.K. Corn Growth and Development; PMR 1009; Iowa State University Extension: Ames, IA, USA, 2011. [Google Scholar]
- Wiseman, B.R.; Davis, F.M.; Williams, W.P.; Widstrom, N.W. Resistance of a maize population, FAWCC(C5), to fall armyworm larvae (Lepidoptera: Noctuidae). Florida Entomologist 1996, 79, 329–336. [Google Scholar] [CrossRef]
- Williams, P.W.; Buckley, P.M.; Davis, F.M. Larval growth, and behavior of the fall armyworm (Lepidoptera: Noctuidae) on callus initiated from susceptible and resistant corn hybrids1. J. Econ. Entomol. 1985, 78, 951–954. [Google Scholar] [CrossRef]
- Videla, G.W.; Davis, F.M.; Williams, P.W.; Seong Ng, S. Fall armyworm (Lepidoptera: Noctuidae) larval growth and survivorship on susceptible and resistant corn at different vegetative growth stages. J. Econ. Entomol. 1992, 85, 2486–2491. [Google Scholar] [CrossRef]
- Davis, F.M.; Wiseman, B.R.; Williams, W.P.; Widstrom, N.W. Insect colony, planting date, and plant growth stage effects on screening maize for leaf-feeding resistance to fall armyworm (Lepidoptera: Noctuidae). Florida Entomologist 1996, 79, 317. [Google Scholar] [CrossRef]
- Bosque-Perez, N.A.; Dabrowski, Z.T. Toward insect resistant maize for the third world. In Proceedings of the International Symposium on Methodologies for Developing Host Plant Resistance to Maize Insects; CIMMYT: México-Veracruz, Mexico, 1989; Available online: https://repository.cimmyt.org/bitstream/handle/10883/706/15875.pdf?sequence=1&isAllowed=y (accessed on 14 December 2021).
- Draper, N.R.; Smith, H. Applied Regression Analysis; John Wiley & Sons: Hoboken, NJ, USA, 1998; Volume 326. [Google Scholar]
- Alvarado, G.; López, M.; Vargas, M.; Pacheco, A.; Rodríguez, F. Multi-Environment Trial Analysis with R. Version 6.04”, hdl11529/10201, CIMMYT. Res. Data Softw. Repos. Netw. 2015. Available online: https://data.cimmyt.org/dataset.xhtml?persistentId=hdl:11529/10201 (accessed on 1 November 2021).
- Peterson, B.G.; Carl, P.; Boudt, K.; Bennett, R.; Ulrich, J.; Zivot, E.; Cornilly, D.; Hung, E.; Lestel, M.; Balkissoon, K. Econometric Tools for Performance and Risk Analysis. R Packag. ‘PerformanceAnalytics’ Version 2019, 153. [Google Scholar]
- CIMMYT. Laboratory Protocols: CIMMYT Applied Molecular Genetics Laboratory, 3rd ed.; CIMMYT: México-Veracruz, Mexico, 2005. [Google Scholar]
- Glaubitz, A.J.; Harriman, J.; Casstevens, T. TASSEL 3.0 Genotyping by Sequencing (GBS) Pipeline Documentation. Pipeline Doc. 2012, 1–20. Available online: https://biohpc.cornell.edu/lab/doc/TasselPipelineGBS20120215.pdf (accessed on 1 November 2021).
- Wang, S.-B.; Feng, J.-Y.; Ren, W.-L.; Huang, B.; Zhou, L.; Wen, Y.-J.; Zhang, J.; Dunwell, J.M.; Xu, S.; Zhang, Y.-M. Improving Power and Accuracy of Genome-Wide Association Studies via a Multi-Locus Mixed Linear Model Methodology. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Stich, B.; Möhring, J.; Piepho, H.P.; Heckenberger, M.; Buckler, E.S.; Melchinger, A.E. Comparison of Mixed-Model Approaches for Association Mapping. Genetics 2008, 178, 1745–1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for Association Mapping of Complex Traits in Diverse Samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Remington, D.L.; Thornsberry, J.M.; Matsuoka, Y.; Wilson, L.M.; Whitt, S.R.; Doebley, J.; Kresovich, S.; Goodman, M.M.; Buckler IV, E.S. Structure of Linkage Disequilibrium and Phenotypic Associations in the Maize Genome. Proc. Natl. Acad. Sci. USA 2001, 98, 11479–11484. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Liu, X.; Wang, J.; Li, M.; Wang, Q.; Tian, F.; Su, Z.; Pan, Y.; Liu, D.; Lipka, A.E.; et al. GAPIT Version 2: An Enhanced Integrated Tool for Genomic Association and Prediction. Plant Genome 2016, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, S.D. Qqman: An R Package for Visualizing GWAS Results Using QQ and Manhattan Plots. J. Open Source Software 2014, 3, 5165. [Google Scholar]
- Woodhouse, M.R.; Cannon, E.K.; Portwood, J.L.; Harper, L.C.; Gardiner, J.M.; Schaeffer, M.L.; Andorf, C.M. A Pan-Genomic Approach to Genome Databases Using Maize as a Model System. BMC Plant Biol. 2021, 21, 385. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Gowda, M.; Liu, W.; Würschum, T.; Maurer, H.P.; Longin, F.H.; Ranc, N.; Reif, J.C. Accuracy of Genomic Selection in European Maize Elite Breeding Populations. Theor. Appl. Genet. 2012, 124, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, J.C.M. Prediction of Response to Marker-Assisted and Genomic Selection Using Selection Index Theory. Clin. Electrophysiol. 2007, 124, 331–341. [Google Scholar] [CrossRef]
- Myles, S.; Peiffer, J.; Brown, P.J.; Ersoz, E.S.; Zhang, Z.; Costich, D.E.; Buckler, E. Association Mapping: Critical Considerations Shift from Genotyping to Experimental Design. Plant Cell 2009, 21, 2194–2202. [Google Scholar] [CrossRef] [Green Version]
- Fraval, S.; Hammond, J.; Bogard, J.R.; Ng, M.; van Etten, J.; Herrero, M.; Oosting, S.J.; de Boer, I.J.; Lannerstad, M.; Teufel, N.; et al. Food Access Deficiencies in Sub-Saharan Africa: Prevalence and Implications for Agricultural Interventions. Front. Sustain. Food Syst. 2019, 3, 104. [Google Scholar] [CrossRef] [Green Version]
- FAO. Multi-Stakeholder Regional Workshop in Africa: Innovations for Smallholder Farmers for Sustainable Management of Fall Armyworm; FAO: Praia, Cabo Verde, 2019. [Google Scholar]
- Nabity, P.D.; Zavala, J.A.; DeLucia, E.H. Indirect Suppression of Photosynthesis on Individual Leaves by Arthropod Herbivory. Ann. Bot. 2009, 103, 655–663. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Du, Y.; Warburton, M.L.; Xiao, Y.; Yan, J. Phenotypic Plasticity Contributes to Maize Adaptation and Heterosis. Mol. Biol. Evol. 2021, 38, 1262–1275. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.S.; Aggarwal, V.D.; Chirwa, R.M. Adaptability and Stability of Bean Cultivars as Determined via Yield-Stability Statistic and GGE Biplot Analysis. J. Crop. Improv. 2006, 15, 97–120. [Google Scholar] [CrossRef]
- Manolio, T.A.; Collins, F.S.; Cox, N.J.; Goldstein, D.B.; Hindorff, L.A.; Hunter, D.J.; McCarthy, M.I.; Ramos, E.M.; Cardon, L.R.; Chakravarti, A.; et al. Finding the Missing Heritability of Complex Diseases. Nature 2009, 461, 747–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.H.; Gail, M.H.; Weinberg, C.R.; Carroll, R.J.; Chung, C.C.; Wang, Z.; Chanock, S.J.; Fraumeni, J.F.; Chatterjee, N. Distribution of Allele Frequencies and Effect Sizes and Their Interrelationships for Common Genetic Susceptibility Variants. Proc. Natl. Acad. Sci. USA 2011, 108, 18026–18031. [Google Scholar] [CrossRef] [Green Version]
- Akinwale, M.G.; Gregorio, G.; Nwilene, F.; Akinyele, B.O.; Ogunbayo, S.A.; Odiyi, A.C. Heritability and Correlation Coefficient Analysis for Yield and Its Components in Rice (Oryza Sativa, L.). Afr. J. plant Sci. 2011, 5, 207–212. [Google Scholar]
- Alqudah, A.M.; Sallam, A.; Baenziger, P.S.; Börner, A. GWAS: Fast-Forwarding Gene Identification and Characterization in Temperate Cereals: Lessons from Barley—A Review. J. Adv. Res. 2020, 22, 119–135. [Google Scholar] [CrossRef]
- Flint-Garcia, S.A.; Thornsberry, J.M.; Edward, S.B., IV. Structure of Linkage Disequilibrium in Plants. Annu. Rev. Plant Biol. 2003, 54, 357–374. [Google Scholar] [CrossRef] [Green Version]
- Badji, A.; Otim, M.; Machida, L.; Odong, T.; Kwemoi, D.B.; Okii, D.; Agbahoungba, S.; Mwila, N.; Kumi, F.; Ibanda, A.; et al. Maize Combined Insect Resistance Genomic Regions and Their Co-Localization with Cell Wall Constituents Revealed by Tissue-Specific QTL Meta-Analyses. Front. Plant Sci. 2018, 9, 895. [Google Scholar] [CrossRef]
- Kibe, M.; Nyaga, C.; Nair, S.K.; Beyene, Y.; Das, B.; Suresh, L.M.; Bright, J.M.; Makumbi, D.; Kinyua, J.; Olsen, M.S.; et al. Combination of Linkage Mapping, GWAS, and GP to Dissect the Genetic Basis of Common Rust Resistance in Tropical Maize Germplasm. Int. J. Mol. Sci. 2020, 21, 6518. [Google Scholar] [CrossRef]
- Lu, Y.; Yan, J.; Guimarães, C.T.; Taba, S.; Hao, Z.; Gao, S.; Chen, S.; Li, J.; Zhang, S.; Vivek, B.S.; et al. Molecular Characterization of Global Maize Breeding Germplasm Based on Genome-Wide Single Nucleotide Polymorphisms. Theor. Appl. Genet. 2009, 120, 93–115. [Google Scholar] [CrossRef] [PubMed]
- Munyiri, S.W.; Mugo, S.N. Quantitative Trait Loci for Resistance to Spotted and African Maize Stem Borers (Chilo partellus and Busseola fusca) in a Tropical Maize (Zea mays L.) Population. Afr. J. Biotechnol. 2017, 16, 1579–1589. [Google Scholar] [CrossRef] [Green Version]
- Womack, E.D.; Williams, W.P.; Smith, J.S.; Warburton, M.L.; Bhattramakki, D.; Hesler, L. Mapping Quantitative Trait Loci for Resistance to Fall Armyworm (Lepidoptera: Noctuidae) Leaf-Feeding Damage in Maize Inbred Mp705. J. Econ. Entomol. 2020, 113, 956–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Womack, E.D.; Erika, D.; Warburton, M.L.; Williams, W.P. Mapping of quantitative trait loci for resistance to fall armyworm and southwestern corn borer leaf-feeding damage in maize. Crop. Sci. 2018, 58, 529–539. [Google Scholar] [CrossRef]
- Navarro, J.A.R.; Willcox, M.; Burgueño, J.; Romay, C.; Swarts, K.; Trachsel, S.; Preciado, E.; Terron, A.; Delgado, H.V.; Vidal, V. A Study of Allelic Diversity Underlying Flowering-Time Adaptation in Maize Landraces. Nat. Genet. 2017, 49, 476–480. [Google Scholar] [CrossRef]
- Whitt, S.R.; Wilson, L.M.; Tenaillon, M.I.; Gaut, B.S.; Buckler, E.S., IV. Genetic Diversity and Selection in the Maize Starch Pathway. Proc. Natl. Acad. Sci. USA 2002, 99, 12959–12962. [Google Scholar] [CrossRef] [Green Version]
- De Lange, E.S.; Balmer, D.; Mauch-Mani, B.; Turlings, T.C.J. Insect and Pathogen Attack and Resistance in Maize and Its Wild Ancestors, the Teosintes. New Phytol. 2014, 204, 329–341. [Google Scholar] [CrossRef]
- Afzal, A.J.; Wood, A.J.; Lightfoot, D.A. Plant Receptor-Like Serine Threonine Kinases: Roles in Signaling and Plant Defense. APS 2008, 21, 507–517. [Google Scholar] [CrossRef] [Green Version]
- Palmer, N.A.; Basu, S.; Heng-Moss, T.; Bradshaw, J.D.; Sarath, G.; Louis, J. Fall Armyworm (Spodoptera frugiperda Smith) Feeding Elicits Differential Defense Responses in Upland and Lowland Switchgrass. PLoS ONE 2019, 14, 1–16. [Google Scholar] [CrossRef]
- Attaran, E.; Major, I.T.; Cruz, J.A.; Rosa, B.A.; Koo, A.J.K.; Chen, J.; Kramer, D.M.; He, S.Y.; Howe, G.A. Temporal Dynamics of Growth and Photosynthesis Suppression in Response to Jasmonate Signaling 1. Plant Physiol. 2014, 165, 1302–1314. [Google Scholar] [CrossRef] [Green Version]
- Beyene, Y.; Gowda, M.; Olsen, M.; Robbins, K.R. Empirical Comparison of Tropical Maize Hybrids Selected Through Genomic and Phenotypic Selections. Front. Plant Sci. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowda, M.; Makumbi, D.; Das, B.; Nyaga, C.; Kosgei, T.; Crossa, J.; Beyene, Y.; Montesinos-López, O.A.; Olsen, M.S.; Prasanna, B.M. Genetic Dissection of Striga Hermonthica (Del.) Benth. Resistance via Genome-Wide Association and Genomic Prediction in Tropical Maize Germplasm. Theor. Appl. Genet. 2021, 134, 941–958. [Google Scholar] [CrossRef] [PubMed]
- Sitonik, C.; Yoseph, L.M.S.; Michael, B.; Dan, S.O.; Kiplagat, M. Genetic Architecture of Maize Chlorotic Mottle Virus and Maize Lethal Necrosis through GWAS, Linkage Analysis and Genomic Prediction in Tropical Maize Germplasm. Theor. Appl. Genet. 2019, 132, 2381–2399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowda, M.; Das, B.; Makumbi, D.; Babu, R.; Semagn, K.; Prasanna, B.M. Genome—Wide Association and Genomic Prediction of Resistance to Maize Lethal Necrosis Disease in Tropical Maize Germplasm. Theor. Appl. Genet. 2015, 128, 1957–1968. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | LD1 | LD2 | LD3 | Eardam | ER (%) | GY (tons/ha) |
|---|---|---|---|---|---|---|
| CKIR04005 | 2.85 | 4.83 | 4.82 | 1.98 | 3.16 | 6.54 |
| CKDHL1920804 | 2.93 | 5.41 | 4.94 | 2.09 | 4.14 | 6.31 |
| CKDHL1922286 | 2.64 | 5.09 | 4.94 | 2.11 | 11.18 | 5.99 |
| CKDHL1920877 | 2.33 | 4.83 | 4.45 | 2.16 | 5.96 | 5.95 |
| CKDHL1923759 | 3.04 | 5.56 | 4.74 | 1.92 | 3.42 | 5.84 |
| CKDHL1921755 | 2.62 | 5.48 | 5.49 | 2.26 | 9.67 | 5.80 |
| CKDHL1922245 | 2.62 | 5.06 | 4.89 | 2.03 | 5.55 | 5.63 |
| CKDHL1922428 | 2.60 | 5.43 | 5.02 | 2.26 | 5.72 | 5.57 |
| CKDHL1922266 | 2.87 | 5.24 | 4.98 | 2.03 | 5.87 | 5.57 |
| CKDHL1922467 | 2.66 | 5.11 | 5.22 | 2.68 | 13.53 | 5.57 |
| CKDHL1920715 | 2.47 | 4.93 | 4.82 | 2.01 | 4.34 | 5.54 |
| CKDHL1920580 | 3.08 | 5.51 | 5.11 | 2.21 | 8.32 | 5.54 |
| CKDHL1924395 | 2.77 | 5.39 | 4.96 | 2.93 | 15.31 | 5.45 |
| CKDHL1922258 | 3.43 | 5.81 | 4.53 | 2.36 | 5.86 | 5.44 |
| CKDHL1924391 | 2.61 | 5.27 | 5.09 | 2.28 | 5.15 | 5.40 |
| Trait | Mean | σ2G | σ2GxE | σ2e | h2 | LSD5% | CV (%) |
|---|---|---|---|---|---|---|---|
| LD1 | 2.78 | 0.06 ** | 0.04 ** | 0.10 | 0.58 | 0.42 | 7.71 |
| LD2 | 5.32 | 0.09 ** | 0.05 ** | 0.35 | 0.45 | 0.65 | 6.21 |
| LD3 | 5.07 | 0.08 * | 0.18 ** | 0.18 | 0.38 | 0.59 | 5.91 |
| Eardam | 2.28 | 0.14 ** | 0.14 ** | 0.19 | 0.55 | 0.62 | 13.82 |
| ER | 7.26 | 14.78 ** | 16.19 ** | 21.97 | 0.52 | 6.63 | 46.44 |
| GY | 3.45 | 0.97 ** | 0.42 ** | 1.54 | 0.62 | 1.74 | 25.69 |
| SNP | Chr | MLM-p Value | MAF | Allele Effect | Putative Candidate Genes | Predicted Function of Candidate Gene |
|---|---|---|---|---|---|---|
| Foliar damage 1 | ||||||
| DT1_86151758 | 1 | 1.32 × 10−05 | 0.34 | −0.04 | No hit | - |
| DT3_189787778 | 3 | 5.43 × 10−05 | 0.17 | −0.06 | GRMZM2G024992 | Uncharacterized |
| DT3_3627388 | 3 | 1.63 × 10−04 | 0.31 | 0.05 | GRMZM2G045259 | ATP binding, protein serine threonine kinase activity |
| DT4_240657423 | 4 | 9.17 × 10−05 | 0.17 | 0.06 | GRMZM2G124151 | Transferring glycosyl groups |
| DT5_193883551 | 5 | 8.48 × 10−07 | 0.11 | 0.08 | No hit | - |
| DT5_172818477 | 5 | 1.67 × 10−04 | 0.40 | −0.04 | GRMZM2G038536 | Base excision repair (BER) pathway, by catalyzing the ADP-ribosylation of acceptor proteins involved in chromatin architecture and DNA metabolism. |
| DT8_174365183 | 8 | 8.22 × 10−05 | 0.46 | −0.05 | GRMZM2G370044 | Uncharacterized |
| DT9_102187311 | 9 | 2.62 × 10−08 | 0.44 | −0.06 | GRMZMG036921 | Transfer RNA intron lyase |
| DT9_6789620 | 9 | 1.16 × 10−06 | 0.39 | −0.02 | GRMZM2G017257 | Chloroplast accumulation movement, ATP-dependent microtubule motor activity |
| Foliar damage 2 | ||||||
| DT1_26003816 | 1 | 1.49 × 10−04 | 0.43 | 0.04 | GRMZM2G015804 | Carbohydrate metabolic process |
| DT2_200197508 | 2 | 5.45 × 10−06 | 0.49 | 0.04 | GRMZM2G703307 | Integral component of membrane |
| DT3_203549581 | 3 | 3.39 × 10−05 | 0.14 | −0.06 | GRMZM2G078756 | Phenylalanyl-tRNA aminoacylation |
| DT4_241323024 | 4 | 8.75 × 10−06 | 0.48 | 0.05 | GRMZM2G051004 | NAD binding |
| DT4_24639576 | 4 | 5.93 × 10−05 | 0.39 | −0.04 | GRMZM2G088169 | Cell fate determination |
| DT5_1996596 | 5 | 7.65 × 10−07 | 0.45 | −0.05 | GRMZM2G415498 | DNA-mediated transposition, |
| DT6_166058896 | 6 | 2.12 × 10−05 | 0.27 | −0.04 | GRMZM2G094892 | Regulation of long-day photoperiodism, flowering |
| DT7_4773701 | 7 | 1.63 × 10−05 | 0.39 | 0.04 | GRMZM2G480002 | Uncharacterized |
| DT7_108615586 | 7 | 2.16 × 10−04 | 0.20 | 0.06 | No hit | - |
| DT8_165429441 | 8 | 2.36 × 10−05 | 0.21 | 0.06 | GRMZM2G016802 | Defense response |
| DT9_96875821 | 9 | 8.88 × 10−05 | 0.40 | 0.04 | GRMZM2GO48919 | Uncharacterized |
| Foliar damage 3 | ||||||
| DT1_5722917 | 1 | 8.05 × 10−05 | 0.30 | 0.07 | GRMZM2G319022 | Uncharacterized |
| DT1_6086007 | 1 | 1.40 × 10−04 | 0.29 | −0.05 | GRMZM2G341918 | Uncharacterized |
| DT2_151852785 | 2 | 7.22 × 10−06 | 0.43 | 0.06 | No hit | - |
| DT4_167218393 | 4 | 2.04 × 10−05 | 0.34 | −0.07 | GRMZM2G168369 | Uncharacterized |
| DT7_27787652 | 7 | 4.50 × 10−06 | 0.40 | 0.06 | GRMZM2G097719 | Uncharacterized |
| DT7_167536749 | 7 | 1.75 × 10−05 | 0.23 | −0.08 | GRMZM2G017145 | Protein dimerization activity |
| DT8_151149212 | 8 | 5.24 × 10−06 | 0.39 | −0.07 | GRMZM2G043117 | Hydrotropism |
| DT8_165270110 | 8 | 7.42 × 10−05 | 0.32 | 6.50 | GRMZM2G114046 | Cellular macromolecule, metabolic process, integral component of membrane |
| Ear damage | ||||||
| DT1_34838367 | 1 | 1.90 × 10−05 | 0.36 | −0.12 | No hit | - |
| DT1_245864468 | 1 | 3.17 × 10−05 | 0.49 | −0.12 | No hit | - |
| DT2_16546600 | 2 | 1.95 × 10−06 | 0.45 | 0.00 | GRMZM2G042756 | Dehydration responsive element binding protein, DNA-binding transcription factor activity |
| DT2_220742831 | 2 | 1.27 × 10−05 | 0.28 | −0.13 | GRMZM2G077256 | Uncharacterized |
| DT2_192225273 | 2 | 2.31 × 10−05 | 0.10 | −0.14 | No hit | - |
| DT3_117394631 | 3 | 5.07 × 10−08 | 0.10 | 0.20 | No hit | - |
| DT4_12954089 | 4 | 9.87× 10−08 | 0.34 | 0.16 | GRMZM2G377115 | Chlorophyll catabolic process, response to water deprivation |
| DT4_202434317 | 4 | 7.31 × 10−05 | 0.41 | −0.08 | No hit | - |
| DT5_174018428 | 5 | 2.99 × 10−05 | 0.27 | 0.12 | No hit | - |
| DT5_86480332 | 5 | 8.55 × 10−05 | 0.22 | −0.09 | No hit | - |
| DT6_159770879 | 6 | 4.30 × 10−07 | 0.11 | 0.12 | No hit | - |
| DT6_108383751 | 6 | 2.65 × 10−04 | 0.44 | 0.11 | GRMZM5G809695 | Regulation of cell growth |
| DT10_138678949 | 10 | 1.71 × 10−04 | 0.18 | −0.09 | No hit | - |
| Ear rot | ||||||
| DT1_32439894 | 1 | 5.56 × 10−05 | 0.08 | 0.15 | No hit | - |
| DT2_64332803 | 2 | 1.53 × 10−05 | 0.45 | −0.87 | GRMZM2G373828 | |
| DT2_210205836 | 2 | 7.53 × 10−05 | 0.19 | −1.06 | GRMZM2G102138 | |
| DT3_53768832 | 3 | 5.82 × 10−05 | 0.20 | −0.65 | No hit | - |
| DT4_4804227 | 4 | 3.62 × 10−09 | 0.33 | 1.00 | AC214255.3_FG008 | |
| DT5_86480332 | 5 | 7.01 × 10−08 | 0.22 | 1.37 | No hit | - |
| DT5_15869219 | 5 | 2.93 × 10−05 | 0.15 | 1.10 | No hit | - |
| DT6_99748720 | 6 | 1.26 × 10−04 | 0.17 | −0.75 | AC215906.3_FG001 | |
| DT7_142857000 | 7 | 1.46 × 10−04 | 0.22 | 0.83 | GRMZM2G074472 | |
| DT9_23210980 | 9 | 7.53 × 10−06 | 0.36 | 0.63 | No hit | - |
| DT9_130788433 | 9 | 2.17 × 10−04 | 0.47 | 0.39 | GRMZM2G043295 | UDP-glycosyltransferase activity |
| DT10_8711707 | 10 | 4.37 × 10−11 | 0.15 | −1.76 | GRMZM2G167999 | Transducin/WD40 repeat-like superfamily protein |
| DT10_140174620 | 10 | 8.46 × 10−06 | 0.20 | 1.01 | GRMZM5G887345 | PF00179: Ubiquitin-conjugating enzyme |
| Grain yield | ||||||
| DT4_60154355 | 4 | 4.28 × 10−04 | 0.23 | −0.26 | No hit | - |
| DT10_145757365 | 10 | 2.46 × 10−04 | 0.40 | 0.25 | GRMZM2G0101264 | Uncharacterized |
| DT10_141438482 | 10 | 4.28 × 10−04 | 0.42 | −0.30 | No hit | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamweru, I.; Anani, B.Y.; Beyene, Y.; Makumbi, D.; Adetimirin, V.O.; Prasanna, B.M.; Gowda, M. Genomic Analysis of Resistance to Fall Armyworm (Spodoptera frugiperda) in CIMMYT Maize Lines. Genes 2022, 13, 251. https://doi.org/10.3390/genes13020251
Kamweru I, Anani BY, Beyene Y, Makumbi D, Adetimirin VO, Prasanna BM, Gowda M. Genomic Analysis of Resistance to Fall Armyworm (Spodoptera frugiperda) in CIMMYT Maize Lines. Genes. 2022; 13(2):251. https://doi.org/10.3390/genes13020251
Chicago/Turabian StyleKamweru, Isaac, Bruce Y. Anani, Yoseph Beyene, Dan Makumbi, Victor O. Adetimirin, Boddupalli M. Prasanna, and Manje Gowda. 2022. "Genomic Analysis of Resistance to Fall Armyworm (Spodoptera frugiperda) in CIMMYT Maize Lines" Genes 13, no. 2: 251. https://doi.org/10.3390/genes13020251