
Transfer of the Resistance to Multiple Diseases from a Triticum-Secale-Thinopyrum Trigeneric Hybrid to Ningmai 13 and Yangmai 23 Wheat Using Specific Molecular Markers and GISH

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
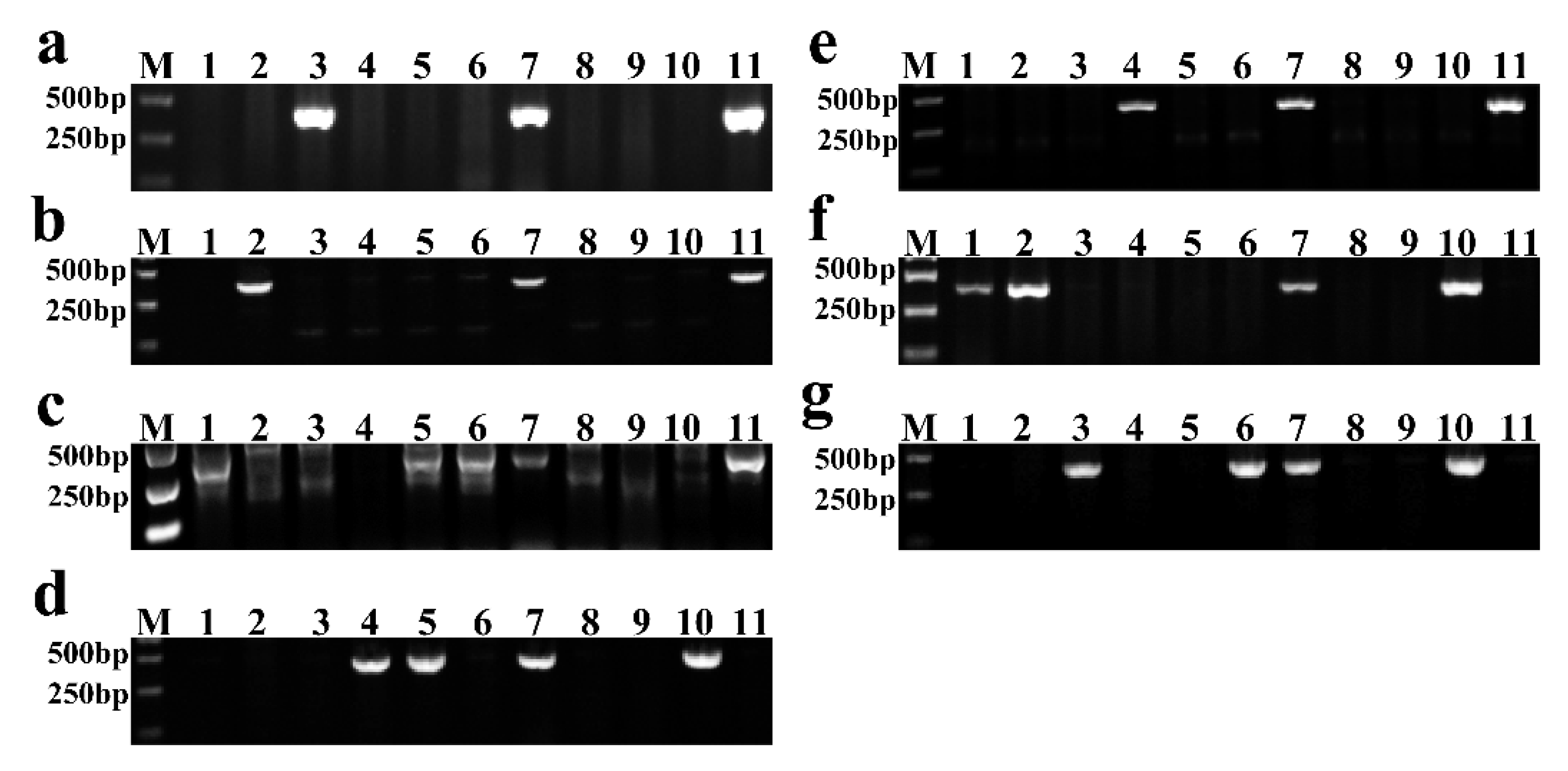
2.2. Molecular Marker Analysis
2.3. Cytogenetic Analyses
2.4. Evaluation of Disease Resistance
3. Results
3.1. Creating New Breeding Lines Carrying Alien Chromosomes from Trigeneric Hybrid Line YZU21
3.2. Individual Alien Chromosome Identification of New Breeding Lines Using Chromosome-Specific Markers
3.3. Chromosome Constitution Analysis of New Breeding Lines by Multicolor GISH
3.4. Evaluation of Disease Resistance
4. Discussion
4.1. The Application of Related Species Is an Effective Approach to Wheat Genetic Improvement
4.2. The Specific Molecular Marker and GISH Can Accurately Identify Trigeneric Hybrid
4.3. The novel Lines of Trigeneric Hybrid Provide New Parents in Wheat Breeding for Multiple Disease Resistance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mayer, K.F.X.; Rogers, J.; Dolezel, J.; Pozniak, C.; Eversole, K.; Feuillet, C.; Gill, B.; Friebe, B.; Lukaszewski, A.J.; Sourdille, P.; et al. A chromosome-based draft sequence of the hexaploid bread wheat (Triticum aestivum) genome. Science 2014, 345, 6194. [Google Scholar] [CrossRef]
- Geiger, H.H.; Miedaner, T. Rye Breeding. In Cereals, 2nd ed.; Carena, M.J., Ed.; Springer Science + Business Media, LLC: Berlin/Heidelberg, Germany, 2009; Volume 3, pp. 157–181. [Google Scholar] [CrossRef] [Green Version]
- Heun, M.; Friebe, B. Introgression of powdery mildew resistance from rye into wheat. Phytopathology 1990, 80, 242–245. [Google Scholar] [CrossRef]
- Hsam, S.L.K.; Zeller, F.J. Evidence of allelism between genes Pm8 and Pm17 and chromosomal location of powdery mildew and leaf rust resistance genes in the common wheat cultivar ‘Amigo’. Plant Breed. 1997, 116, 119–122. [Google Scholar] [CrossRef]
- Friebe, B.; Heun, M.; Tuleen, N.; Zeller, F.J.; Gill, B.S. Cytogenetically monitored transfer of powdery mildew resistance from rye into wheat. Crop Sci. 1994, 34, 621. [Google Scholar] [CrossRef]
- Hao, M.; Liu, M.; Luo, J.T.; Fan, C.L.; Yi, Y.Y.; Zhang, L.Q.; Yuan, Z.W.; Ning, S.Z.; Zheng, Y.L.; Liu, D.C. Introgression of powdery mildew resistance gene Pm56 on rye chromosome arm 6RS into wheat. Front. Plant Sci. 2018, 9, 1040. [Google Scholar] [CrossRef]
- Rao, M.P.; Ramanujam, S. The transfer of alien genes for stem rust resistance to durum wheat. In Proceedings of the International Wheat Genetics Symposium, New Delhi, India, 23–28 February 1978. [Google Scholar]
- Mago, R.; Miah, H.; Lawrence, G.J.; Wellings, C.R.; Spielmeyer, W.; Bariana, H.S.; McIntosh, R.A.; Pryor, A.J.; Ellis, J.G. High-resolution mapping and mutation analysis separate the rust resistance genes Sr31, Lr26 and Yr9 on the short arm of rye chromosome 1. Theor. Appl. Genet. 2005, 112, 41–50. [Google Scholar] [CrossRef]
- Mago, R.; Zhang, P.; Vautrin, S.; Šimková, H.; Bansal, U.; Luo, M.C.; Rouse, M.; Karaoglu, H.; Periyannan, S.; Kolmer, J.; et al. The wheat Sr50 gene reveals rich diversity at a cereal disease resistance locus. Nat. Plants 2015, 1, 15186. [Google Scholar] [CrossRef]
- Rahmatov, M.; Rouse, M.N.; Nirmala, J.; Danilova, T.; Friebe, B.; Steffenson, B.J.; Johansson, E. A new 2DS·2RL robertsonian translocation transfers stem rust resistance gene Sr59 into wheat. Theor. Appl. Genet. 2016, 129, 1383–1392. [Google Scholar] [CrossRef]
- Mcintosh, R.A.; Wellings, C.R.; Park, R.F. The genes for resistance to stripe rust in wheat and triticale. In Wheat Rusts: An Atlas of Resistance Genes, 2nd ed.; Jeans, K., Cloud-Guest, A., Eds.; CSIRO Publications: Clayton, Australia, 1995; pp. 29–81. [Google Scholar] [CrossRef]
- Ren, T.H.; Jiang, Q.; Sun, Z.X.; Zhao, L.Q.; Peng, W.H.; Ren, Z.L.; Tan, F.Q.; Luo, P.G.; Li, Z. Development and molecular cytogenetic characterization of novel primary wheat-rye 1RS.1BL translocation lines from multiple rye sources with resistance to stripe rust. Plant Dis. 2022, 106, 2191–2200. [Google Scholar] [CrossRef]
- Lafiandra, D.; Riccardi, G.; Shewry, P.R. Improving cereal grain carbohydrates for diet and health. J. Cereal Sci. 2014, 59, 312–326. [Google Scholar] [CrossRef] [Green Version]
- Góral, T.; Wiśniewska, H.; Ochodzki, O.; Walentyn-Góral, D. Higher Fusarium toxin accumulation in grain of winter Triticale lines inoculated with Fusarium culmorum as compared with wheat. Toxins 2016, 8, 301. [Google Scholar] [CrossRef] [Green Version]
- Jauhar, P.P.; Peterson, T.S.; Xu, S.S. Cytogenetic and molecular characterization of a durum alien disomic addition line with enhanced tolerance to Fusarium head blight. Genome 2009, 52, 467–483. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Ohm, H. Fusarium head blight resistance derived from Lophopyrum elongatum chromosome 7E and its augmentation with Fhb1 in wheat. Plant Breed. 2006, 125, 424–429. [Google Scholar] [CrossRef]
- Liu, H.P.; Dai, Y.; Chi, D.; Huang, S.; Li, H.; Duan, Y.M.; Cao, W.G.; Gao, Y.; Fedak, G.; Chen, J.M. Production and molecular cytogenetic characterization of a durum Wheat-Thinopyrum elongatum 7E disomic addition line with resistance to Fusarium head blight. Cytogenet. Genome Res. 2018, 153, 165–173. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, X.L.; Hou, Y.L.; Cai, J.J.; Shen, X.R.; Zhou, T.T.; Xu, H.H.; Ohm, H.W.; Wang, H.W.; Li, A.F.; et al. High-density mapping of the major FHB resistance gene Fhb7 derived from Thinopyrum ponticum and its pyramiding with Fhb1 by marker-assisted selection. Theor. Appl. Genet. 2015, 128, 2301–2316. [Google Scholar] [CrossRef]
- Wang, H.W.; Sun, S.L.; Ge, W.Y.; Zhao, L.F.; Hou, B.Q.; Wang, K.; Lyu, Z.F.; Chen, L.Y.; Xu, S.S.; Guo, J.; et al. Horizontal gene transfer of Fhb7 from fungus underlies Fusarium head blight resistance in wheat. Science 2020, 368, eaba5435. [Google Scholar] [CrossRef]
- Dai, Y.; Duan, Y.M.; Liu, H.P.; Chi, D.; Cao, W.G.; Xue, A.; Yong, G.; George, F.; Chen, J.M. Molecular cytogenetic characterization of two Triticum-Secale-Thinopyrum trigeneric hybrids exhibiting superior resistance to Fusarium head blight, leaf rust, and stem rust race Ug99. Front. Plant Sci. 2017, 8, 797. [Google Scholar] [CrossRef] [Green Version]
- Lei, J.; Zhou, J.W.; Sun, H.J.; Wan, W.T.; Xiao, J.; Yuan, C.X.; Karafiátová, M.; Doležel, J.; Wang, H.Y.; Wang, X.E. Development of oligonucleotide probes for FISH karyotyping in Haynaldia villosa, a wild relative of common wheat. Crop J. 2020, 8, 676–681. [Google Scholar] [CrossRef]
- Zhao, R.H.; Wang, H.Y.; Xiao, J.; Bie, T.D.; Cheng, S.H.; Jia, Q.; Yuan, C.X.; Zhang, R.Q.; Cao, A.Z.; Chen, P.D.; et al. Induction of 4VS chromosome recombinants using the CS ph1b mutant and mapping of the wheat yellow mosaic virus resistance gene from Haynaldia villosa. Theor. Appl. Genet. 2013, 126, 2921–2930. [Google Scholar] [CrossRef]
- Yang, H.; Zhong, S.F.; Chen, C.; Yang, H.; Chen, W.; Tan, F.Q.; Zhang, M.; Chen, W.Q.; Ren, T.H.; Li, Z.; et al. Identification and cloning of a CC-NBS-NBS-LRR gene as a candidate of Pm40 by integrated analysis of both the available transcriptional data and published linkage mapping. Int. J. Mol. Sci. 2021, 22, 10239. [Google Scholar] [CrossRef]
- Ren, T.H.; Sun, Z.X.; Ren, Z.L.; Tan, F.Q.; Luo, P.G.; Tang, Z.X.; Fu, S.L.; Li, Z. Molecular and cytogenetic characterization of a Wheat-Rye 7BS.7RL translocation line with resistance to stripe rust, powdery mildew, and Fusarium head blight. Phytopathology 2020, 110, 1713–1720. [Google Scholar] [CrossRef]
- Xie, C.; Sun, Q.; Ni, Z.; Yang, T.; Nevo, E.; Fahima, T. Identification of resistance gene analogue markers closely linked to wheat powdery mildew resistance gene Pm31. Plant Breed. 2004, 123, 198–200. [Google Scholar] [CrossRef]
- Warburton, M.L.; Crossa, J.; Franco, J.; Kazi, M.; Trethowan, R.; Rajaram, S.; Pfeiffer, W.; Zhang, P.; Dreisigacker, S.; Ginkel, M. Bringing wild relatives back into the family: Recovering genetic diversity in CIMMYT improved wheat germplasm. Euphytica 2006, 149, 289–301. [Google Scholar] [CrossRef]
- Luo, P.G.; Ren, Z.L.; Wu, X.H.; Zhang, H.Y.; Zhang, H.Q.; Feng, J.A. Structural and biochemical mechanism responsible for the stay-green phenotype in common wheat. Chin. Sci. Bull. 2006, 51, 2595–2603. [Google Scholar] [CrossRef]
- Qi, W.L.; Tang, Y.; Zhu, W.; Li, D.Y.; Diao, C.D.; Xu, L.L.; Zeng, J.; Wang, Y.; Fan, X.; Sha, L.N.; et al. Molecular cytogenetic characterization of a new wheat-rye 1BL1RS translocation line expressing superior stripe rust resistance and enhanced grain yield. Planta 2016, 244, 405–416. [Google Scholar] [CrossRef]
- Li, S.Q.; Tang, H.P.; Zhang, H.; Mu, Y.; Lan, X.J.; Ma, J. A 1BL/1RS translocation contributing to kernel length increase in three wheat recombinant inbred line populations. Czech J. Genet. Plant Breed. 2020, 56, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Spetsov, P.; Daskalova, N. Resistance to pathogens in wheat-rye and triticale genetic stocks. J. Plant Pathol. 2022, 104, 99–114. [Google Scholar] [CrossRef]
- Hou, L.Y.; Jia, J.Q.; Zhang, X.J.; Li, X.; Yang, Z.J.; Ma, J.; Guo, H.J.; Zhan, H.X.; Qiao, L.Y.; Chang, Z.J. Molecular mapping of the stripe rust resistance gene Yr69 on wheat chromosome 2AS. Plant Dis. 2016, 100, 1717–1724. [Google Scholar] [CrossRef]
- Li, H.J.; Wang, X.M. Thinopyrum ponticum and Th. intermedium: The promising source of resistance to fungal and viral diseases of wheat. J. Genet. Genom. 2009, 36, 557–565. [Google Scholar] [CrossRef]
- Zhang, R.Q.; Xiong, C.X.; Mu, H.Q.; Yao, R.N.; Meng, X.R.; Kong, L.R.; Xing, L.P.; Wu, J.Z.; Feng, Y.G.; Cao, A.Z. Pm67, a new powdery mildew resistance gene transferred from Dasypyrum villosum chromosome 1V to common wheat (Triticum aestivum L.). Crop J. 2021, 9, 882–888. [Google Scholar] [CrossRef]
- Zhang, R.Q.; Fan, Y.L.; Kong, L.N.; Wang, Z.J.; Wu, J.Z.; Xing, L.P.; Cao, A.Z.; Feng, Y.G. Pm62, an adult-plant powdery mildew resistance gene introgressed from Dasypyrum villosum chromosome arm 2VL into wheat. Theor. Appl. Genet. 2018, 131, 2613–2620. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.Q.; Sun, B.X.; Chen, J.; Cao, A.Z.; Xing, L.P.; Feng, Y.G.; Lan, C.X.; Chen, P.D. Pm55, a developmental-stage and tissue-specific powdery mildew resistance gene introgressed from Dasypyrum villosum into common wheat. Theor. Appl. Genet. 2016, 129, 1975–1984. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.D.; Qi, L.L.; Zhou, B.; Zhang, S.Z.; Liu, D.J. Development and molecular cytogenetic analysis of wheat-Haynaldia villosa 6VS/6AL translocation lines specifying resistance to powdery mildew. Theor. Appl. Genet. 1995, 91, 1125–1128. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, A.; Jones, S.S.; Murray, T.D.; Line, R.F. Evaluation of Dasypyrum villosum populations for resistance to cereal eyespot and stripe rust pathogens. Plant Dis. 2000, 84, 40–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Pace, C.; Snidaro, D.; Ciaffi, M.; Vittori, D.; Ciofo, A.; Cenci, A.; Tanzarella, O.A.; Qualset, C.O.; Mugnozza, G.T.S. Introgression of Dasypyrum villosum chromatin into common wheat improves grain protein quality. Euphytica 2001, 117, 67–75. [Google Scholar] [CrossRef]
- Qi, L.; Cao, M.; Chen, P.; Li, W.; Liu, D. Identification, mapping, and application of polymorphic DNA associated with resistance gene Pm21 of wheat. Genome 1996, 39, 191–197. [Google Scholar] [CrossRef]
- Zhang, R.Q.; Hou, F.; Feng, Y.G.; Zhang, W.; Zhang, M.Y.; Chen, P.D. Characterization of a Triticum aestivum-Dasypyrum villosum T2VS center dot 2DL translocation line expressing a longer spike and more kernels traits. Theor. Appl. Genet. 2015, 128, 2415–2425. [Google Scholar] [CrossRef]
- Feng, Z.Y.; Song, L.; Song, W.J.; Qi, Z.Q.; Yuan, J.; Li, R.; Han, H.M.; Wang, H.F.; Chen, Z.Y.; Guo, W.L.; et al. The decreased expression of GW2 homologous genes contributed to the increased grain width and thousand-grain weight in wheat-Dasypyrum villosum 6VS center dot 6DL translocation lines. Theor. Appl. Genet. 2021, 134, 3873–3894. [Google Scholar] [CrossRef]
- Chen, P.D.; You, C.F.; Hu, Y.; Chen, S.W.; Zhou, B.; Cao, A.Z.; Wang, X.E. Radiation-induced translocations with reduced Haynaldia villosa chromatin at the Pm21 locus for powdery mildew resistance in wheat. Mol. Breeding 2013, 31, 477–484. [Google Scholar] [CrossRef]
- Zhou, S.H.; Zhang, J.P.; Han, H.M.; Zhang, J.; Ma, H.H.; Zhang, Z.; Lu, Y.Q.; Liu, W.H.; Yang, X.M.; Li, X.Q.; et al. Full-length transcriptome sequences of Agropyron cristatum facilitate the prediction of putative genes for thousand-grain weight in a wheat-A. cristatum translocation line. BMC Genom. 2019, 20, 1025. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhang, J.P.; Liu, W.H.; Wu, X.Y.; Yang, X.M.; Li, X.Q.; Lu, Y.Q.; Li, L.H. An intercalary translocation from Agropyron cristatum 6P chromosome into common wheat confers enhanced kernel number per spike. Planta 2016, 244, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Heslop-Harrison, J.S. RNA, genes, genomes and chromosomes: Repetitive DNA sequences in plants. In Chromosomes Today; Olmo, E., Redl, C.A., Eds.; John Innes Centre: Norwich, UK, 2000; Volume 13, pp. 45–56. [Google Scholar] [CrossRef]
- Du, P.; Zhuang, L.F.; Wang, Y.Z.; Yuan, L.; Wang, Q.; Wang, D.R.; Dawadondup; Tan, L.J.; Shen, J.; Xu, H.B.; et al. Development of oligonucleotides and multiplex probes for quick and accurate identification of wheat and Thinopyrum bessarabicum chromosomes. Genome 2017, 60, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.H.; Sun, Z.X.; Hu, Y.L.; Ren, Z.L.; Tan, F.Q.; Luo, P.G.; Li, Z. Molecular cytogenetic identification of new wheat-rye 6R, 6RS, and 6RL addition lines with resistance to stripe rust and powdery mildew. Front. Plant Sci. 2022, 13, 992016. [Google Scholar] [CrossRef] [PubMed]
- Zaharieva, M.; Monneveux, P. Spontaneous hybridization between bread wheat (Triticum aestivum L.) and its wild relatives in Europe. Crop Sci. 2006, 46, 512–527. [Google Scholar] [CrossRef]
- Fernández-Escobar, J.; Martin, A. A self-fertile trigeneric hybrid in the Triticeae involving Triticum, Hordeum, and Secale. Euphytica 1989, 42, 291–296. [Google Scholar] [CrossRef]
- Yu, C.J.; Jia, X.; Hu, S.Q.; Zhuang, J.J. Cytogenetics of the hybrids and their pollen plants of three genera Triticum Secale and Thinopyrum. Acta Genetica Sinica 1994, 21, 447–452. [Google Scholar]
- Li, X.F.; Song, Z.Q.; Liu, S.B.; Gao, J.R.; Wang, H.G. Cytogenetic study of a trigeneric (Triticale × Trileymus) hybrid. Euphytica 2006, 150, 117–122. [Google Scholar] [CrossRef]
- Gupta, P.K.; Fedak, G. Intergeneric hybrids between × Triticosecale cv. Welsh (2n = 42) and three genotypes of Agropyron intermedium (2n = 42). Genome 2011, 28, 176–179. [Google Scholar] [CrossRef]
- Orellana, J.; Vazquez, J.F.; Carrillo, J.M. Genome analysis in wheat-rye-Aegilops caudata trigeneric hybrids. Genome 1989, 32, 169–172. [Google Scholar] [CrossRef]
- Kang, H.Y.; Tang, L.; Li, D.Y.; Diao, C.D.; Zhu, W.; Tang, Y.; Wang, Y.; Fan, X.; Xu, L.L.; Zeng, J.; et al. Cytogenetic study and stripe rust response of the derivatives from a wheat-Thinopyrum intermedium-Psathyrostachys huashanica trigeneric hybrid. Genome 2017, 60, 393–401. [Google Scholar] [CrossRef] [Green Version]
- Li, J.B.; Lang, T.; Li, B.; Yu, Z.H.; Wang, H.J.; Li, G.R.; Yang, E.N.; Yang, Z.J. Introduction of Thinopyrum intermedium ssp trichophorum chromosomes to wheat by trigeneric hybridization involving Triticum, Secale and Thinopyrum genera. Planta 2017, 245, 1121–1135. [Google Scholar] [CrossRef] [PubMed]
- Friebe, B.; Jiang, J.; Raupp, W.J.; McIntosh, R.A.; Gill, B.S. Characterization of wheat-alien translocations conferring resistance to diseases and pests: Current status. Euphytica 1996, 91, 59–87. [Google Scholar] [CrossRef]
- Johansson, E.; Henriksson, T.; Prieto-Linde, M.L.; Andersson, S.; Ashraf, R.; Rahmatov, M. Diverse wheat-alien introgression lines as a basis for durable resistance and quality characteristics in bread wheat. Front. Plant Sci. 2020, 11, 1067. [Google Scholar] [CrossRef] [PubMed]
- Li, J.B.; Dundas, L.; Dong, C.M.; Li, G.R.; Trethowan, R.; Yang, Z.J.; Hoxha, S.; Zhang, P. Identification and characterization of a new stripe rust resistance gene Yr83 on rye chromosome 6R in wheat. Theor. Appl. Genet. 2020, 133, 1095–1107. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.L.; Luo, J.; Yang, Z.J.; Li, G.R.; Tang, Z.X.; Fu, S.L. The physical location of stripe rust resistance genes on chromosome 6 of rye (Secale cereale L.) AR106BONE. Front. Plant Sci. 2022, 13, 928014. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Line | Hybrid Combination | Generation |
|---|---|---|
| YZU006/1 | NM13/YZU21 | F6 |
| YZU006/5 | NM13/YZU21 | F6 |
| YZU035 | YM23//YM23/YZU21 | BC1F4 |
| YZU047/1 | YM23//YM23/YZU21 | BC1F4 |
| YZU047/2 | YM23//YM23/YZU21 | BC1F4 |
| YZU048 | YM23//YM23/YZU21 | BC1F4 |
| Line | Alien Chromosome | Chromosome Number | The Chromosome Composition |
|---|---|---|---|
| YZU006/1 | 3E and 6R | 42 | 19″WW + 1″3E + 1″6R |
| YZU006/5 | 2E and 6R | 44 | 20″WW + 1″2E + 1″6R |
| YZU035 | 1E and 7R | 44 | 20″WW + 1″1E + 1″7R |
| YZU047/1 | 4R and 5E | 42 | 19″WW +1′W + 1′4R + 1″5E |
| YZU047/2 | 3E and 4R | 46 | 21″WW + 1″3E + 1″4R |
| YZU048 | 3E and 7R | 42 | 20″WW + 1″3E/7R translocation |
| Line | Average Disease Spikelet Rate (%) |
|---|---|
| YZU006/1 | 7.7 ± 2.8 de |
| YZU006/5 | 9.9 ± 0.6 d |
| YZU035 | 7.9 ± 2.3 d |
| YZU047/1 | 9.6 ± 0.5 d |
| YZU047/2 | 16.1 ± 2.6 c |
| YZU048 | 13.1 ± 2.3 cd |
| NM 13 | 16.4 ± 2.4 c |
| YZU21 | 6.3 ± 2.4 de |
| YM 23 | 20.3 ± 3.0 b |
| SU 3 | 5.9 ± 2.5 e |
| An8455 | 87.2 ± 2.0 a |
| Line | Bgt | Pst | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E09 Race | ITs | E15 Race | ITs | A13 Race | ITs | A44 Race | ITs | CYR32 Race | ITs | CYR34 Race | ITs | |
| YZU006/1 | 3 | MS | 4 | HS | 4 | HS | 3 | MS | 3 | HR | 7 | MS |
| YZU006/5 | 0; | HR | 4 | HS | 2 | MR | 4 | HS | 4 | MR | 7 | MS |
| YZU035 | 4 | HS | 4 | HS | 4 | HS | 4 | HS | 7 | MS | 7 | MS |
| YZU047/1 | 3 | MS | 4 | HS | 4 | HS | 4 | HS | 7 | MS | 7 | MS |
| YZU047/2 | 4 | HS | 4 | HS | 4 | HS | 4 | HS | 7 | MS | 7 | MS |
| YZU048 | 4 | HS | 3 | MS | 4 | HS | 4 | HS | 7 | MS | 7 | MS |
| NM13 | 3 | MS | 4 | HS | 4 | HS | 3 | MS | 7 | MS | 7 | MS |
| YZU21 | 0 | I | 0 | I | 4 | HS | 2 | MR | 7 | MS | 8 | MS |
| YM23 | 4 | HS | 4 | HS | 4 | HS | 4 | HS | 2 | HR | 1 | HR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, Y.; Shi, J.; Li, J.; Gao, Y.; Ma, H.; Wang, Y.; Wang, B.; Chen, J.; Cheng, P.; Ma, H. Transfer of the Resistance to Multiple Diseases from a Triticum-Secale-Thinopyrum Trigeneric Hybrid to Ningmai 13 and Yangmai 23 Wheat Using Specific Molecular Markers and GISH. Genes 2022, 13, 2345. https://doi.org/10.3390/genes13122345
Dai Y, Shi J, Li J, Gao Y, Ma H, Wang Y, Wang B, Chen J, Cheng P, Ma H. Transfer of the Resistance to Multiple Diseases from a Triticum-Secale-Thinopyrum Trigeneric Hybrid to Ningmai 13 and Yangmai 23 Wheat Using Specific Molecular Markers and GISH. Genes. 2022; 13(12):2345. https://doi.org/10.3390/genes13122345
Chicago/Turabian StyleDai, Yi, Juntao Shi, Jinfeng Li, Yujiao Gao, Haigang Ma, Yonggang Wang, Baotong Wang, Jianmin Chen, Peng Cheng, and Hongxiang Ma. 2022. "Transfer of the Resistance to Multiple Diseases from a Triticum-Secale-Thinopyrum Trigeneric Hybrid to Ningmai 13 and Yangmai 23 Wheat Using Specific Molecular Markers and GISH" Genes 13, no. 12: 2345. https://doi.org/10.3390/genes13122345