
Copy Number Variation among Resistance Genes Analogues in Brassica napus

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
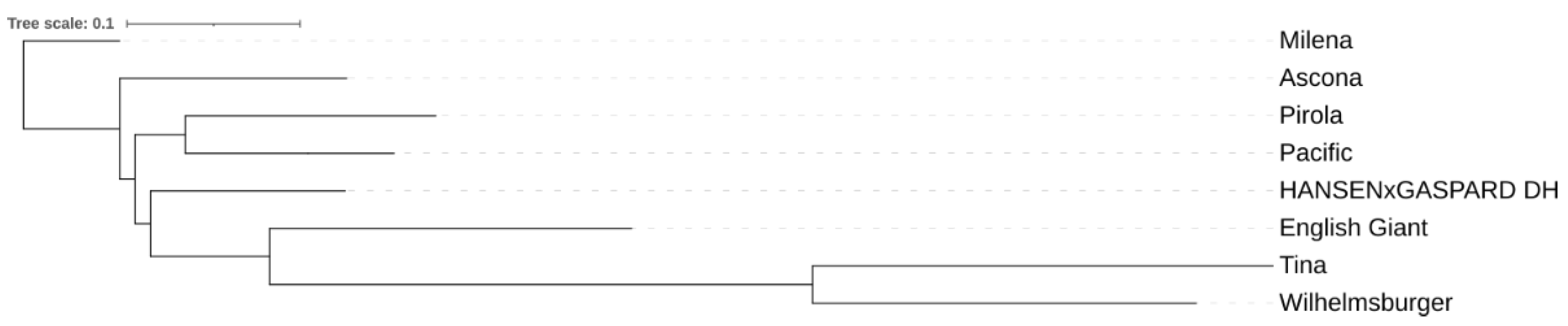
2.1. Plant Materials
2.2. DNA Extraction and Quantification
2.3. Construction of Genomic DNA Libraries and Sequencing
2.4. Sequencing Data Processing and Read Alignments
2.5. CNV Calling
2.6. RGA Prediction and Physical Clustering
2.7. QTL and Genomic Data Representation
3. Results
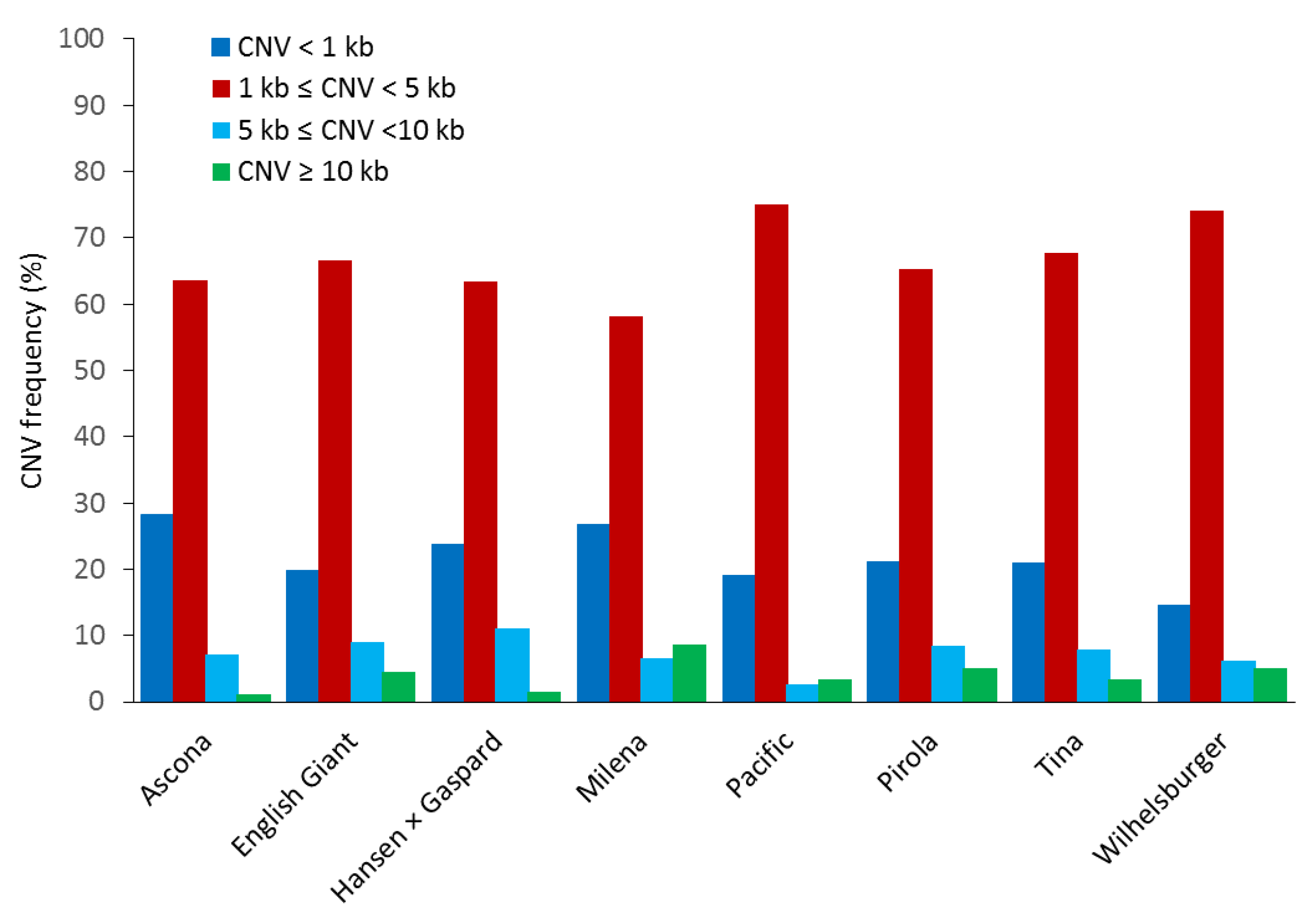
3.1. CNV Analysis
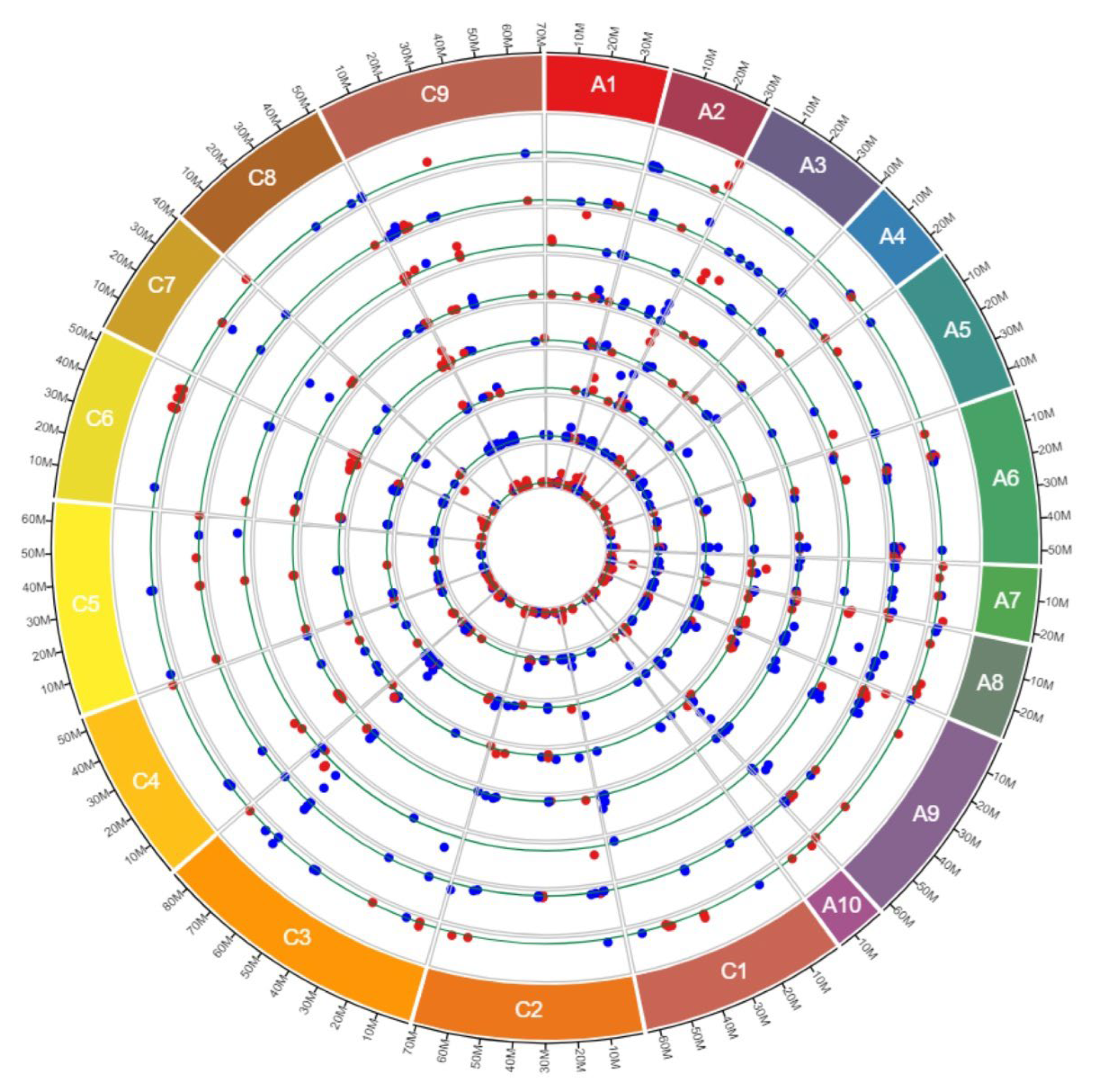
3.2. Distribution along Chromosomes and Sub-Genomes
3.3. CNVs across RGAs
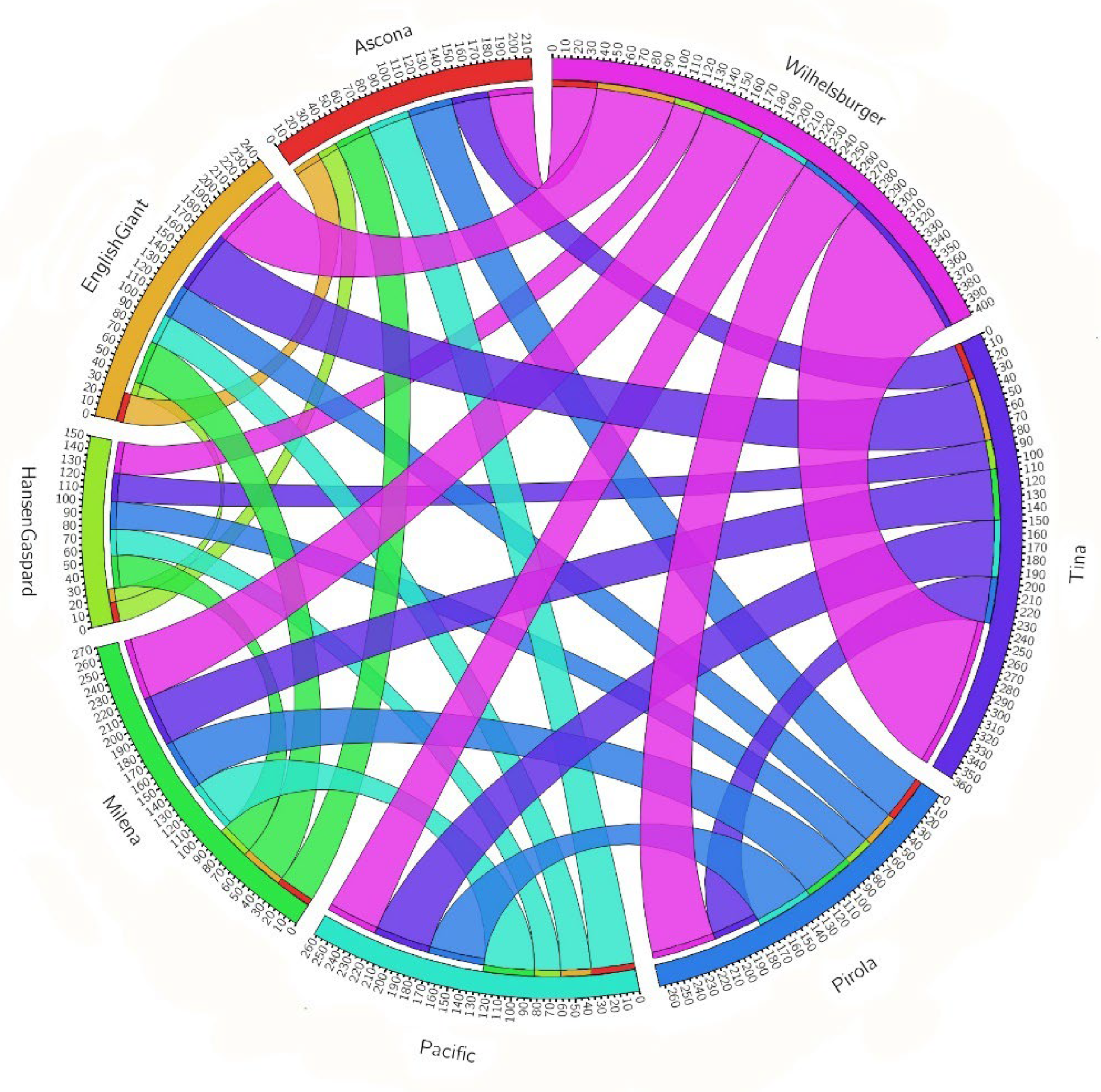
3.4. Gene-Physical Clustering
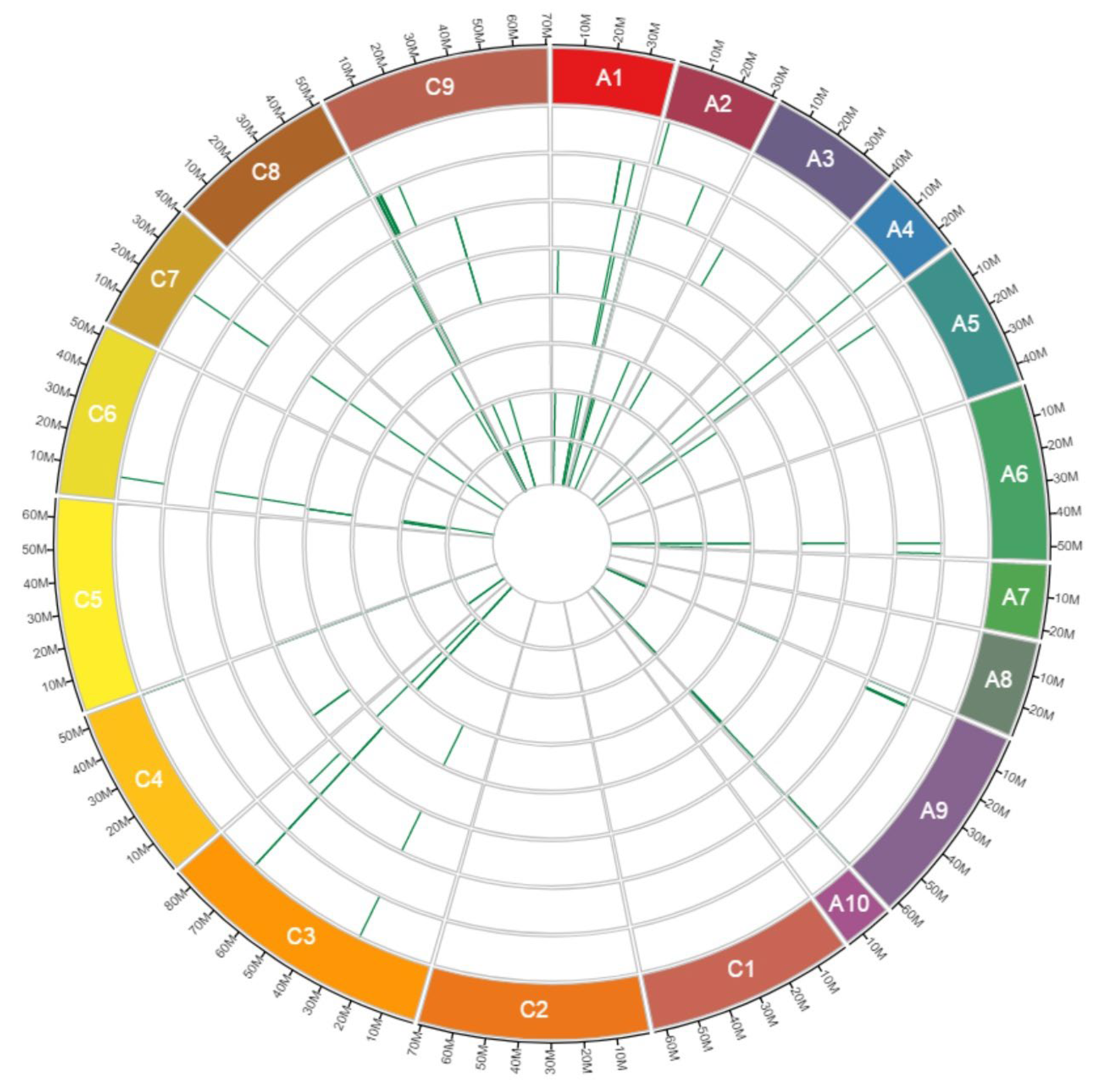
3.5. Investigating of RGAs Affected by CNV Events across Known Genomic Regions for Blackleg Resistance Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Golicz, A.A.; Bayer, P.E.; Bhalla, P.L.; Batley, J.; Edwards, D. Pangenomics comes of age: From bacteria to plant and animal applications. Trends Genet. 2020, 36, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Bayer, P.E.; Golicz, A.A.; Scheben, A.; Batley, J.; Edwards, D. Plant pan-genomes are the new reference. Nat. Plants 2020, 6, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Dolatabadian, A.; Bayer, P.E.; Tirnaz, S.; Hurgobin, B.; Edwards, D.; Batley, J. Characterization of disease resistance genes in the Brassica napus pangenome reveals significant structural variation. Plant Biotechnol. J. 2020, 18, 969–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurgobin, B.; Golicz, A.A.; Bayer, P.E.; Chan, C.K.; Tirnaz, S.; Dolatabadian, A.; Schiessl, S.V.; Samans, B.; Montenegro, J.D.; Parkin, I.A.P.; et al. Homoeologous exchange is a major cause of gene presence/absence variation in the amphidiploid Brassica napus. Plant Biotechnol. J. 2018, 16, 1265–1274. [Google Scholar] [CrossRef] [Green Version]
- Bayer, P.E.; Scheben, A.; Golicz, A.A.; Yuan, Y.; Faure, S.; Lee, H.; Chawla, H.S.; Anderson, R.; Bancroft, I.; Raman, H.; et al. Modelling of gene loss propensity in the pangenomes of three Brassica species suggests different mechanisms between polyploids and diploids. Plant Biotechnol. J. 2021, 19, 2488–2500. [Google Scholar] [CrossRef] [PubMed]
- Golicz, A.A.; Bayer, P.E.; Barker, G.C.; Edger, P.P.; Kim, H.; Martinez, P.A.; Chan, C.K.K.; Severn-Ellis, A.; McCombie, W.R.; Parkin, I.A.P.; et al. The pangenome of an agronomically important crop plant Brassica oleracea. Nat. Commun. 2016, 7, 13390. [Google Scholar] [CrossRef] [Green Version]
- Bayer, P.E.; Golicz, A.A.; Tirnaz, S.; Chan, C.K.; Edwards, D.; Batley, J. Variation in abundance of predicted resistance genes in the Brassica oleracea pangenome. Plant Biotechnol. J. 2019, 17, 789–800. [Google Scholar] [CrossRef] [Green Version]
- Montenegro, J.D.; Golicz, A.A.; Bayer, P.E.; Hurgobin, B.; Lee, H.; Chan, C.K.K.; Visendi, P.; Lai, K.; Doležel, J.; Batley, J. The pangenome of hexaploid bread wheat. Plant J. 2017, 90, 1007–1013. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Scheben, A.; Verpaalen, B.; Tirnaz, S.; Bayer, P.E.; Hodel, R.G.J.; Batley, J.; Soltis, D.E.; Soltis, P.S.; Edwards, D. Amborella gene presence/absence variation is associated with abiotic stress responses that may contribute to environmental adaptation. New Phytol. 2021, 233, 1548–1555. [Google Scholar] [CrossRef]
- Zhao, J.; Bayer, P.; Ruperao, P.; Saxena, R.; Khan, A.; Golicz, A.; Nguyen, H.; Batley, J.; Edwards, D.; Varshney, R. Trait associations in the pangenome of pigeon pea (Cajanus cajan). Plant Biotechnol. J. 2020, 18, 1946–1954. [Google Scholar] [CrossRef]
- Yu, J.; Golicz, A.A.; Lu, K.; Dossa, K.; Zhang, Y.; Chen, J.; Wang, L.; You, J.; Fan, D.; Edwards, D. Insight into the evolution and functional characteristics of the pan-genome assembly from sesame landraces and modern cultivars. Plant Biotechnol. J. 2019, 17, 881–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Q.; Feng, Q.; Lu, H.; Li, Y.; Wang, A.; Tian, Q.; Zhan, Q.; Lu, Y.; Zhang, L.; Huang, T.; et al. Pan-genome analysis highlights the extent of genomic variation in cultivated and wild rice. Nat. Genet. 2018, 50, 278–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Chebotarov, D.; Kudrna, D.; Llaca, V.; Lee, S.; Rajasekar, S.; Mohammed, N.; Al-Bader, N.; Sobel-Sorenson, C.; Parakkal, P. A platinum standard pan-genome resource that represents the population structure of Asian rice. Sci. Data 2020, 7, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Du, H.; Li, P.; Shen, Y.; Peng, H.; Liu, S.; Zhou, G.A.; Zhang, H.; Liu, Z.; Shi, M.; et al. Pan-Genome of Wild and Cultivated Soybeans. Cell 2020, 182, 162–176.e13. [Google Scholar] [CrossRef]
- Bayer, P.E.; Valliyodan, B.; Hu, H.; Marsh, J.I.; Yuan, Y.; Vuong, T.D.; Patil, G.; Song, Q.; Batley, J.; Varshney, R.K. Sequencing the USDA core soybean collection reveals gene loss during domestication and breeding. Plant Genome 2021, 15, e20109. [Google Scholar] [CrossRef]
- Torkamaneh, D.; Lemay, M.A.; Belzile, F. The pan-genome of the cultivated soybean (PanSoy) reveals an extraordinarily conserved gene content. Plant Biotechnol. J. 2021, 19, 1852–1862. [Google Scholar] [CrossRef]
- Rijzaani, H.; Bayer, P.E.; Rouard, M.; Doležel, J.; Batley, J.; Edwards, D. The pangenome of banana highlights differences between genera and genomes. Plant Genome 2021, 15, e20100. [Google Scholar] [CrossRef]
- Varshney, R.; Nayak, S.; May, G.; Jackson, S. Next-generation sequencing technologies and their implications for crop genetics and breeding. Trends Biotechnol. 2009, 27, 522–530. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Lu, T.; Han, B. Resequencing rice genomes: An emerging new era of rice genomics. Trends Genet. 2013, 29, 225–232. [Google Scholar] [CrossRef]
- Wang, H.; Chai, Z.; Hu, D.; Ji, Q.; Xin, J.; Zhang, C.; Zhong, J. A global analysis of CNVs in diverse yak populations using whole-genome resequencing. BMC Genom. 2019, 20, 61. [Google Scholar] [CrossRef]
- Murthy, M.; Veerappa, A.M.; Seshachalam, K.; Ramachandra, N. High-resolution arrays reveal burden of copy number variations on Parkinson disease genes associated with increased disease risk in random cohorts. Neurol. Res. 2016, 38, 775–785. [Google Scholar] [CrossRef]
- Gamazon, E.; Stranger, B. The impact of human copy number variation on gene expression. Brief. Funct. Genom. 2015, 14, 352–357. [Google Scholar] [CrossRef] [Green Version]
- Hastings, P.; Lupski, J.; Rosenberg, S.; Ira, G. Mechanisms of change in gene copy number. Nat. Rev. Genet. 2009, 10, 551–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrider, D.; Hahn, M. Gene copy-number polymorphism in nature. Proc. R. Soc. B Biol. Sci. 2010, 277, 3213–3221. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Yang, N.; Cheng, H.; Song, J.; Qu, L. Genome-wide identification of copy number variations between two chicken lines that differ in genetic resistance to Marek’s disease. BMC Genom. 2015, 16, 843. [Google Scholar] [CrossRef] [Green Version]
- Hull, R.; Cruz, C.; Jack, C.; Houseley, J. Environmental change drives accelerated adaptation through stimulated copy number variation. PLoS Biol. 2017, 15, e2001333. [Google Scholar] [CrossRef] [Green Version]
- Bai, Z.; Chen, J.; Liao, Y.; Wang, M.; Liu, R.; Ge, S.; Wing, R.; Chen, M. The impact and origin of copy number variations in the Oryza species. BMC Genom. 2016, 17, 261. [Google Scholar] [CrossRef] [Green Version]
- DeBolt, S. Copy number variation shapes genome diversity in Arabidopsis over immediate family generational scales. Genome Biol. Evol. 2010, 2, 441–453. [Google Scholar] [CrossRef] [Green Version]
- Schiessl, S.; Huettel, B.; Kuehn, D.; Reinhardt, R.; Snowdon, R.J. Targeted deep sequencing of flowering regulators in Brassica napus reveals extensive copy number variation. Sci. Data 2017, 4, 170013. [Google Scholar] [CrossRef] [Green Version]
- Redon, R.; Ishikawa, S.; Fitch, K.; Feuk, L.; Perry, G.; Andrews, T.; Fiegler, H.; Shapero, M.; Carson, A.; Chen, W.; et al. Global variation in copy number in the human genome. Nature 2006, 444, 444–454. [Google Scholar] [CrossRef]
- Springer, N.; Ying, K.; Fu, Y.; Ji, T.; Yeh, C.; Jia, Y.; Wu, W.; Richmond, T.; Kitzman, J.; Rosenbaum, H.; et al. Maize inbreds exhibit high levels of copy number variation. PLoS Genet. 2009, 5, e1000734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson-Wagner, R.; Eichten, S.; Kumari, S.; Tiffin, P.; Stein, J.; Ware, D.; Springer, N. Pervasive gene content variation and copy number variation in maize and its undomesticated progenitor. Genome Res. 2010, 20, 1689–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maron, L.; Guimaraes, C.; Kirst, M.; Albert, P.; Birchler, J.; Bradbury, P.; Buckler, E.; Coluccio, A.; Danilova, T.; Kudrna, D.; et al. Aluminum tolerance in maize is associated with higher MATE1 gene copy number. Proc. Natl. Acad. Sci. USA 2013, 110, 5241–5246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zmienko, A.; Samelak-Czajka, A.; Kozlowski, P.; Szymanska, M.; Figlerowicz, M. Arabidopsis thaliana population analysis reveals high plasticity of the genomic region spanning MSH2, AT3G18530 and AT3G18535 genes and provides evidence for NAHR-driven recurrent CNV events occurring in this location. BMC Genom. 2016, 17, 893. [Google Scholar] [CrossRef] [Green Version]
- Zmienko, A.; Marszalek-Zenczak, M.; Wojciechowski, P.; Samelak-Czajka, A.; Luczak, M.; Kozlowski, P.; Karlowski, W.M.; Figlerowicz, M. AthCNV: A Map of DNA Copy Number Variations in the Arabidopsis Genome [OPEN]. Plant Cell 2020, 32, 1797–1819. [Google Scholar] [CrossRef] [Green Version]
- Yu, P.; Wang, C.; Xu, Q.; Feng, Y.; Yuan, X.; Yu, H.; Wang, Y.; Tang, S.; Wei, X. Detection of copy number variations in rice using array-based comparative genomic hybridization. BMC Genom. 2011, 12, 372. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Wang, Y.; Zheng, J.; Wen, Y.; Qu, M.; Kang, S.; Wu, S.; Deng, X.; Hong, K.; Li, S.; et al. A genome-wide survey of copy number variations reveals an asymmetric evolution of duplicated genes in rice. BMC Biol. 2020, 18, 73. [Google Scholar] [CrossRef]
- Saintenac, C.; Jiang, D.; Akhunov, E. Targeted analysis of nucleotide and copy number variation by exon capture in allotetraploid wheat genome. Genome Biol. 2011, 12, R88. [Google Scholar] [CrossRef] [Green Version]
- Diaz, A.; Zikhali, M.; Turner, A.; Isaac, P.; Laurie, D. Copy number variation affecting the photoperiod-B1 and vernalization-A1 genes is associated with altered flowering time in wheat. PLoS ONE 2012, 7, e33234. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Saghai Maroof, M.A.; Allard, R. Effects on adaptedness of variations in ribosomal DNA copy number in populations of wild barley. Proc. Natl. Acad. Sci. USA 1990, 87, 8741–8745. [Google Scholar] [CrossRef]
- Sutton, T.; Baumann, U.; Hayes, J.; Collins, N.; Shi, B.; Schnurbusch, T.; Hay, A.; Mayo, G.; Pallotta, M.; Tester, M.; et al. Boron-toxicity tolerance in barley arising from efflux transporter amplification. Science 2007, 318, 1446–1449. [Google Scholar] [CrossRef]
- Nitcher, R.; Distelfeld, A.; Tan, C.; Yan, L.; Dubcovsky, J. Increased copy number at the HvFT1 locus is associated with accelerated flowering time in barley. Mol. Genet. Genom. 2013, 288, 261–275. [Google Scholar] [CrossRef] [Green Version]
- Datta, S.; Jankowicz-Cieslak, J.; Nielen, S.; Ingelbrecht, I.; Till, B. Induction and recovery of copy number variation in banana through gamma irradiation and low-coverage whole-genome sequencing. Plant Biotechnol. J. 2018, 16, 1644–1653. [Google Scholar] [CrossRef] [Green Version]
- Alonge, M.; Wang, X.; Benoit, M.; Soyk, S.; Pereira, L.; Zhang, L.; Suresh, H.; Ramakrishnan, S.; Maumus, F.; Ciren, D. Major Impacts of widespread structural variation on gene expression and crop improvement in tomato. Cell 2020, 182, 145–161.e123. [Google Scholar] [CrossRef]
- Cook, D.; Lee, T.; Guo, X.; Melito, S.; Wang, K.; Bayless, A.; Wang, J.; Hughes, T.; Willis, D.; Clemente, T.; et al. Copy number variation of multiple genes at Rhg1 mediates nematode resistance in soybean. Science 2012, 338, 1206–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.; Diers, B.; Hudson, M. An efficient method for measuring copy number variation applied to improvement of nematode resistance in soybean. Plant J. 2016, 88, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Bakker, E.; Toomajian, C.; Kreitman, M.; Bergelson, J. A genome-wide survey of R gene polymorphisms in Arabidopsis. Plant Cell 2006, 18, 1803–1818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.; Araki, H.; Chen, L.; Chen, J.; Tian, D. Unique evolutionary mechanism in R genes under the presence/absence polymorphism in Arabidopsis thaliana. Genetics 2006, 172, 1243–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Liu, X.; Ge, S.; Jensen, J.; Hu, F.; Li, X.; Dong, Y.; Gutenkunst, R.; Fang, L.; Huang, L.; et al. Resequencing 50 accessions of cultivated and wild rice yields markers for identifying agronomically important genes. Nat. Biotechnol. 2012, 30, 105–111. [Google Scholar] [CrossRef]
- González, V.; Aventin, N.; Centeno, E.; Puigdomenech, P. High presence/absence gene variability in defence-related gene clusters of Cucumis melo. BMC Genom. 2013, 14, 782. [Google Scholar] [CrossRef]
- Lin, X.; Zhang, Y.; Kuang, H.; Chen, J. Frequent loss of lineages and deficient duplications accounted for low copy number of disease resistance genes in Cucurbitaceae. BMC Genom. 2013, 14, 335. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Ren, J.; Peng, Z.; Umana, A.; Le, H.; Danilova, T.; Fu, J.; Wang, H.; Robertson, A.; Hulbert, S.; et al. Analysis of extreme phenotype bulk copy number variation. Front. Plant Sci. 2018, 9, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, O.; Kumar, I.; Diers, B.; Hudson, M. Evolution and selection of Rhg1, a copy-number variant nematode-resistance locus. Mol. Ecol. 2015, 24, 1774–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabur, I.; Chawla, H.S.; Lopisso, D.T.; von Tiedemann, A.; Snowdon, R.J.; Obermeier, C. Gene presence-absence variation associates with quantitative Verticillium longisporum disease resistance in Brassica napus. Sci. Rep. 2020, 10, 4131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopec, P.M.; Mikolajczyk, K.; Jajor, E.; Perek, A.; Nowakowska, J.; Obermeier, C.; Chawla, H.S.; Korbas, M.; Bartkowiak-Broda, I.; Karlowski, W.M. Local Duplication of TIR-NBS-LRR Gene Marks Clubroot Resistance in Brassica napus cv. Tosca. Front. Plant Sci. 2021, 12, 639631. [Google Scholar] [CrossRef] [PubMed]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [Green Version]
- Bhattarai, K.; Wang, W.; Cao, Z.; Deng, Z. Comparative Analysis of Impatiens Leaf Transcriptomes Reveal Candidate Genes for Resistance to Downy Mildew Caused by Plasmopara obducens. Int. J. Mol. Sci. 2018, 19, 2057. [Google Scholar] [CrossRef] [Green Version]
- Marti, F.; Saski, C.; Manganaris, G.; Gasic, K.; Crisosto, C. Genomic sequencing of Japanese plum. Front. Plant Sci. 2018, 9, 21. [Google Scholar]
- Mason, A.; Snowdon, R. Oilseed rape: Learning about ancient and recent polyploid evolution from a recent crop species. Plant Biol. 2016, 18, 883–892. [Google Scholar] [CrossRef]
- Szadkowski, E.; Eber, F.; Huteau, V.; Lode, M.; Coriton, O.; Jenczewski, E.; Chevre, A. Polyploid formation pathways have an impact on genetic rearrangements in resynthesized Brassica napus. New Phytol. 2011, 191, 884–894. [Google Scholar] [CrossRef]
- Nicolas, S.; Monod, H.; Eber, F.; Chevre, A.; Jenczewski, E. Non-random distribution of extensive chromosome rearrangements in Brassica napus depends on genome organization. Plant J. 2012, 70, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ecke, W.; Clemens, R.; Honsdorf, N.; Becker, H.C. Extent and structure of linkage disequilibrium in canola quality winter rapeseed (Brassica napus L.). Theor. Appl. Genet. Theor. Und Angew. Genet. 2010, 120, 921–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganya, S.; Svotwa, E.; Katsaruware, R.D. Performance of two Rape (Brassica napus) cultivars under different fertilizer management levels in the smallholder sector of Zimbabwe. Int. J. Agron. 2018, 2018, 2351204. [Google Scholar] [CrossRef]
- Lammerink, J.; Hart, R. ‘Tina’, a new swede cultivar with resistance to dry rot and clubroot. N. Z. J. Exp. Agric. 1985, 13, 417–420. [Google Scholar] [CrossRef]
- Hasan, M.J.; Rahman, H. Genetics and molecular mapping of resistance to Plasmodiophora brassicae pathotypes 2, 3, 5, 6, and 8 in rutabaga (Brassica napus var. napobrassica). Genome 2016, 59, 805. [Google Scholar] [CrossRef] [Green Version]
- Wilch, A. Characterisation of Genotypic and Tissue Specific Resistance in Oilseed Rape (B. napus) Against Sclerotinia sclerotiorum; Georg-August-Universität Göttingen: Göttingen, Germany, 2018. [Google Scholar]
- Lammerink, J. Six pathogenic races of Plasmodiophora brassicae Wor. in New Zealand. N. Z. J. Agric. Res. 1965, 8, 156–164. [Google Scholar] [CrossRef] [Green Version]
- Knights, B.A. Studies in the cruciferae: Sterols in pollen of Brassica napus L. Phytochemistry 1968, 7, 1707–1708. [Google Scholar] [CrossRef]
- Stonard, J.F.; Downes, K.; Pirie, E.; Fitt, B.D.L.; Evans, N. Development of phoma stem canker (Leptosphaeria maculans) and light leaf spot (Pyrenopeziza brassicae) on current and historical oilseed rape cultivars in 2003/04, 2004/05 and 2005/06 UK growing seasons. In Proceedings of the 12th International Rapeseed Congress, Wuhan, China, 26–30 March 2007; pp. 26–30. [Google Scholar]
- Bolger, A.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Chiang, C.; Layer, R.; Faust, G.; Lindberg, M.; Rose, D.; Garrison, E.; Marth, G.; Quinlan, A.R.; Hall, I.M. SpeedSeq: Ultra-fast personal genome analysis and interpretation. Nat. Methods 2015, 12, 966–968. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Tarasov, A.; Vilella, A.; Cuppen, E.; Nijman, I.; Prins, P. Sambamba: Fast processing of NGS alignment formats. Bioinformatics 2015, 31, 2032–2034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abyzov, A.; Urban, A.; Snyder, M.; Gerstein, M. CNVnator: An approach to discover, genotype, and characterize typical and atypical CNVs from family and population genome sequencing. Genome Res. 2011, 21, 974–984. [Google Scholar] [CrossRef] [Green Version]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinlan, A.; Hall, I. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Quan, X.; Jia, G.; Xiao, J.; Cloutier, S.; You, F. RGAugury: A pipeline for genome-wide prediction of resistance gene analogs. BMC Genom. 2016, 17, 852. [Google Scholar] [CrossRef] [Green Version]
- Delourme, R.; Pilet-Nayel, M.; Archipiano, M.; Horvais, R.; Tanguy, X.; Rouxel, T.; Brun, H.; Renard, M.; Balesdent, M. A Cluster of Major Specific Resistance Genes to Leptosphaeria maculans in Brassica napus. Phytopathology 2004, 94, 578–583. [Google Scholar] [CrossRef] [Green Version]
- Raman, R.; Taylor, B.; Marcroft, S.; Stiller, J.; Eckermann, P.; Coombes, N.; Rehman, A.; Lindbeck, K.; Luckett, D.; Wratten, N.; et al. Molecular mapping of qualitative and quantitative loci for resistance to Leptosphaeria maculans causing blackleg disease in canola. Crop Pasture Sci. 2012, 125, 405–418. [Google Scholar] [CrossRef]
- Raman, R.; Taylor, B.; Lindbeck, K.; Coombes, N.; Barbulescu, D.; Salisbury, P.; Raman, H. Molecular mapping and validation of Rlm1 gene for resistance to Leptosphaeria maculans in canola. Crop Pasture Sci. 2012, 63, 1007–1017. [Google Scholar] [CrossRef]
- Leflon, M.; Brun, H.; Eber, F.; Delourme, R.; Lucas, M.; Vallée, P.; Ermel, M.; Balesdent, M.; Chèvre, A. Detection, introgression and localization of genes conferring specific resistance to Leptosphaeria maculans from Brassica rapa into B. TAG Theor. Appl. Genet. Theor. Und Angew. Genet. 2007, 115, 897–906. [Google Scholar] [CrossRef]
- Larkan, N.; Raman, H.; Lydiate, D.; Robinson, S.; Yu, F.; Barbulescu, D.; Raman, R.; Luckett, D.; Burton, W.; Wratten, N.; et al. Multi-environment QTL studies suggest a role for cysteine-rich protein kinase genes in quantitative resistance to blackleg disease in Brassica napus. BMC Plant Biol. 2016, 16, 183. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wurschum, T.; Longin, C.; Hahn, V.; Tucker, M.; Leiser, W. Copy number variations of CBF genes at the Fr-A2 locus are essential components of winter hardiness in wheat. Plant J. 2017, 89, 764–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francia, E.; Morcia, C.; Pasquariello, M.; Mazzamurro, V.; Milc, J.; Rizza, F.; Terzi, V.; Pecchioni, N. Copy number variation at the HvCBF4-HvCBF2 genomic segment is a major component of frost resistance in barley. Plant Mol. Biol. 2016, 92, 161–175. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Feng, Y.; Kumar, D.; Zhang, W.; Zhu, T.; Luo, M.; Messing, J. Analysis of tandem gene copies in maize chromosomal regions reconstructed from long sequence reads. Proc. Natl. Acad. Sci. USA 2016, 113, 7949–7956. [Google Scholar] [CrossRef] [Green Version]
- Demirci, S.; Fuentes, R.R.; Dooijeweert, W.V.; Aflitos, S.; Schijlen, E.; Hesselink, T.; de Ridder, D.; Dijk, A.D.J.V.; Peters, S. Chasing breeding footprints through structural variations in Cucumis melo and wild relatives. G3 Genes|Genomes|Genet. 2021, 11, jkaa038. [Google Scholar] [CrossRef]
- McKernan, K.J.; Helbert, Y.; Kane, L.T.; Ebling, H.; Zhang, L.; Liu, B.; Eaton, Z.; McLaughlin, S.; Kingan, S.; Baybayan, P.; et al. Sequence and annotation of 42 cannabis genomes reveals extensive copy number variation in cannabinoid synthesis and pathogen resistance genes. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Sankoff, D.; Zheng, C.; Zhu, Q. The collapse of gene complement following whole genome duplication. BMC Genom. 2010, 11, 313. [Google Scholar] [CrossRef] [Green Version]
- Lovene, M.; Zhang, T.; Lou, Q.; Buell, C.; Jiang, J. Copy number variation in potato-an asexually propagated autotetraploid species. Plant J. 2013, 75, 80–89. [Google Scholar]
- Schiessl, S.; Katche, E.; Ihien, E.; Chawla, H.; Mason, A. The role of genomic structural variation in the genetic improvement of polyploid crops. Crop J. 2018, 7, 127–140. [Google Scholar] [CrossRef]
- Schiessl, S.; Huettel, B.; Kuehn, D.; Reinhardt, R.; Snowdon, R. Post-polyploidisation morphotype diversification associates with gene copy number variation. Sci. Rep. 2017, 7, 41845. [Google Scholar] [CrossRef] [PubMed]
- Parkin, I.; Koh, C.; Tang, H.; Robinson, S.; Kagale, S.; Clarke, W.; Town, C.; Nixon, J.; Krishnakumar, V.; Bidwell, S.; et al. Transcriptome and methylome profiling reveals relics of genome dominance in the mesopolyploid Brassica oleracea. Genome Biol. 2014, 15, R77. [Google Scholar] [CrossRef] [PubMed]
- Zmienko, A.; Samelak, A.; Kozlowski, P.; Figlerowicz, M. Copy number polymorphism in plant genomes. Theor. Appl. Genet. 2014, 127, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beló, A.; Beatty, M.; Hondred, D.; Fengler, K.; Li, B.; Rafalski, A. Allelic genome structural variations in maize detected by array comparative genome hybridization. Theor. Appl. Genet. 2010, 120, 355–367. [Google Scholar] [CrossRef]
- Demeke, T.; Eng, M. Effect of endogenous reference genes on digital PCR assessment of genetically engineered canola events. Biomol. Detect. Quantif. 2018, 15, 24–29. [Google Scholar] [CrossRef]
- Sekhwal, M.; Li, P.; Lam, I.; Wang, X.; Cloutier, S.; You, F. Disease resistance gene analogs. Int. J. Mol. Sci. 2015, 16, 19248–19290. [Google Scholar] [CrossRef] [Green Version]
- Chisholm, S.; Coaker, G.; Day, B.; Staskawicz, B. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.; Trotochaud, A.E.; Clark, S.E. The Arabidopsis CLAVATA2 gene encodes a receptor-like protein required for the stability of the CLAVATA1 receptor-like kinase. Plant Cell 1999, 11, 1925–1933. [Google Scholar] [CrossRef] [Green Version]
- Nadeau, J.A.; Sack, F.D. Control of stomatal distribution on the Arabidopsis leaf surface. Science 2002, 296, 1697–1700. [Google Scholar] [CrossRef]
- Yang, S.; Feng, Z.; Zhang, X.; Jiang, K.; Jin, X.; Hang, Y.; Chen, J.; Tian, D. Genome-wide investigation on the genetic variations of rice disease resistance genes. Plant Mol. Biol. 2006, 62, 181–193. [Google Scholar] [CrossRef]
- Zhang, M.; Wu, Y.; Lee, M.; Liu, Y.; Rong, Y.; Santos, T.; Wu, C.; Xie, F.; Nelson, R.; Zhang, H. Numbers of genes in the NBS and RLK families vary by more than four-fold within a plant species and are regulated by multiple factors. Nucleic Acids Res. 2010, 38, 6513–6525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Lim, C.J.; Lee, B.; Choi, J.; Oh, S.; Ahmad, R.; Kwon, S.; Ahn, J.; Hur, C. A Genome-wide comparison of NB-LRR type of resistance gene analogs. Mol. Cells 2012, 33, 385–392. [Google Scholar] [CrossRef] [PubMed]
- McHale, L.; Haun, W.; Xu, W.; Bhaskar, P.; Anderson, J.; Hyten, D.; Gerhardt, D.; Jeddeloh, J.; Stupar, R. Structural variants in the soybean genome localize to clusters of biotic stress response genes. Plant Physiol. 2012, 159, 1295–1308. [Google Scholar] [CrossRef] [Green Version]
- McHale, L.; Tan, X.; Koehl, P.; Michelmore, R. Plant NBS-LRR proteins: Adaptable guards. Genome Biol. 2006, 7, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyers, B.; Kozik, A.; Griego, A.; Kuang, H.; Michelmore, R. Genome-wide analysis of NBS-LRR-encoding genes in Arabidopsis. Plant Cell 2003, 15, 809–834. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.; Zhang, Y.; Hu, Q.; Chen, J.; Li, K.; Lu, C.; Liu, H.; Wang, W.; Kuang, H. Dynamic nucleotide-binding site and leucine-rich repeat-encoding genes in the grass family. Plant Physiol. 2012, 159, 197–210. [Google Scholar] [CrossRef] [Green Version]
- Marchal, C.; Zhang, J.; Zhang, P.; Fenwick, P.; Steuernagel, B.; Adamski, N.; Boyd, L.; McIntosh, R.; Wulff, B.; Berry, S. BED-domain containing immune receptors confer diverse resistance spectra to yellow rust. Nat. Plants 2018, 4, 662–668. [Google Scholar] [CrossRef]
- Ashfield, T.; Egan, A.; Pfeil, B.; Chen, N.; Podicheti, R.; Ratnaparkhe, M.; Ameline-Torregrosa, C.; Denny, R.; Cannon, S.; Doyle, J.; et al. Evolution of a complex disease resistance gene cluster in diploid Phaseolus and tetraploid Glycine. Plant Physiol. 2012, 159, 336–354. [Google Scholar] [CrossRef] [Green Version]
- Alamery, S.; Tirnaz, S.; Bayer, P.; Tollenaere, R.; Chalhoub, B.; Edwards, D.; Batley, J. Genome-wide identification and comparative analysis of NBS-LRR resistance genes in Brassica napus. Crop Pasture Sci. 2017, 69, 79–93. [Google Scholar] [CrossRef]
- Kuang, H.; Woo, S.; Meyers, B.; Nevo, E.; Michelmore, R. Multiple genetic processes result in heterogeneous rates of evolution within the major cluster disease resistance genes in lettuce. Plant Cell 2004, 16, 2870–2894. [Google Scholar] [CrossRef] [Green Version]
- Jestin, C.; Lodé, M.; Vallée, P.; Domin, C.; Falentin, C.; Horvais, R.; Coedel, S.; Manzanares-Dauleux, M.; Delourme, R. Association mapping of quantitative resistance for Leptosphaeria maculans in oilseed rape. Mol. Breed. 2011, 27, 271–287. [Google Scholar] [CrossRef]
- Huang, Y.; Jestin, C.; Welham, S.; King, G.; Manzanares-Dauleux, M.; Fitt, B.; Delourme, R. Identification of environmentally stable QTL for resistance against Leptosphaeria maculans in oilseed rape. TAG Theor. Appl. Genet. Theor. Und Angew. Genet. 2016, 129, 169–180. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivars | Deletion | Duplication | Total | Both |
|---|---|---|---|---|
| Ascona | 48 (56.47%) | 37 (43.53%) | 85 | 11 (12.94%) |
| English Giant | 87 (55.77%) | 69 (44.23%) | 156 | 29 (18.59%) |
| Hansen × Gaspard | 33 (52.38%) | 30 (47.62%) | 63 | 17 (26.98%) |
| Milena | 67 (63.81%) | 38 (36.19%) | 105 | 21 (20.00%) |
| Pacific | 58 (50%) | 58 (50%) | 116 | 19 (16.38%) |
| Pirola | 77 (65.25%) | 41 (34.75%) | 118 | 14 (11.86%) |
| Tina | 163 (68.20%) | 76 (31.80%) | 239 | 34 (14.23%) |
| Wilhelmsburger | 171 (67.06%) | 84 (32.94%) | 255 | 43 (16.86%) |
| Total | 704 (61.92%) | 433 (38.08%) | 1137 | 188 (16.53%) |
| Ascona | English Giant | Hansen × Gaspard | Milena | Pacific | Pirola | Tina | Wilhelmsburger | |
|---|---|---|---|---|---|---|---|---|
| Ascona | - | |||||||
| English Giant | 24 | - | ||||||
| Hansen × Gaspard | 17 | 11 | - | |||||
| Milena | 30 | 36 | 27 | - | ||||
| Pacific | 36 | 25 | 22 | 42 | - | |||
| Pirola | 37 | 28 | 23 | 42 | 47 | - | ||
| Tina | 31 | 52 | 24 | 42 | 47 | 38 | - | |
| Wilhelmsburger | 37 | 65 | 26 | 51 | 43 | 54 | 126 | - |
| Shared between | 1 Cultivar | 2 Cultivars | 3 Cultivars | 4 Cultivars | 5 Cultivars | 6 Cultivars | 7 Cultivars | 8 Cultivars | Total |
|---|---|---|---|---|---|---|---|---|---|
| RGAs | 262 | 157 | 81 | 27 | 15 | 14 | 5 | 2 | 563 |
| 46.54% | 27.89% | 14.39% | 4.80% | 2.66% | 2.49% | 0.89% | 0.36% | 100% |
| Locus | Marker | Reference | Chromosome | Start (Mbp) | End (Mbp) | Length (Mbp) | RGA Candidates | RGA Affected by CNV |
|---|---|---|---|---|---|---|---|---|
| Rlm1 | Na12A02 | - | 22.35 | 27.29 | 4.94 | TN 1, OTHER 1, TX 1, RLK 8, NL 1, CNL 1 | RLK 1 (1 del) | |
| Ol12-E03A | Delourme et al., 2004 [79] | |||||||
| CB10544A | Raman, Taylor, Lindbeck et al., 2012 [80] | A7 | ||||||
| Ra2-A05b | - | |||||||
| BSR + KASP | ||||||||
| Rlm3 | BnGMS147b | Delourme et al., 2004 [79] | A7 | 8.97 | 25.75 | 16.79 | RLK 44, RLP 5, NL 4, CNL 1, TNL 3, TN 1, OTHER 1, TX 1 | RLK 14 (12 del, 2 dup), TNL 2 (1 del, 1 dup) |
| IGF0504f_F | Leflon et al., 2007 [82] | |||||||
| Rlm4 | BRMS040 | Raman, Taylor, Marcroft et al., 2012) [80] | A7 | 11.49 | 15.20 | 3.71 | RLK 15, RLP 2 | RLK 9 (8 del, 1 dup) |
| Na12-E11b | ||||||||
| Rlm7 | sR7018 | Larkan et al., 2016 [83] | A7 | 12.16 | 28.19 | 16.02 | RLK 34, RLP 5, NL 4, CNL 2, TNL 3, TN 1, OTHER 1, TX 1 | RLK 8 (7 del, 1 dup), TNL 2 (1 del, 1dup) |
| Rlm9 | BnGMS665 | Delourme et al., 2004 [79] | A7 | 13.76 | 19.11 | 5.35 | RLK 16, RLP 3, NL 2 | RLK 4 (4 del) |
| BnGMS147b | ||||||||
| LepR1 | FlankingMarkers | Larkan et al., 2016 [83] | A02 | 10.02 | 20.43 | 10.41 | RLK 7 | RLK 2 (2 del) |
| LepR2 | FlankingMarkers | Larkan et al., 2016 [83] | A10 | 0.20 | 14.15 | 13.95 | RLK 17, RLP 6, TX 1, NL 1, CN 2, CNL 1, NBS 1 | RLK 4 (3 del, 1dup) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dolatabadian, A.; Yuan, Y.; Bayer, P.E.; Petereit, J.; Severn-Ellis, A.; Tirnaz, S.; Patel, D.; Edwards, D.; Batley, J. Copy Number Variation among Resistance Genes Analogues in Brassica napus. Genes 2022, 13, 2037. https://doi.org/10.3390/genes13112037
Dolatabadian A, Yuan Y, Bayer PE, Petereit J, Severn-Ellis A, Tirnaz S, Patel D, Edwards D, Batley J. Copy Number Variation among Resistance Genes Analogues in Brassica napus. Genes. 2022; 13(11):2037. https://doi.org/10.3390/genes13112037
Chicago/Turabian StyleDolatabadian, Aria, Yuxuan Yuan, Philipp Emanuel Bayer, Jakob Petereit, Anita Severn-Ellis, Soodeh Tirnaz, Dhwani Patel, David Edwards, and Jacqueline Batley. 2022. "Copy Number Variation among Resistance Genes Analogues in Brassica napus" Genes 13, no. 11: 2037. https://doi.org/10.3390/genes13112037