
Genome Mining Reveals High Biosynthetic Potential of Biocontrol Agent Bacillus velezensis B.BV10
 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant-Pathogenic Fungi
2.2. Origin of Isolate B.BV10, Phenotypic Characterization, and Colony Architecture
2.3. Dual Culture Assay
2.4. In Vivo Biocontrol
2.5. Complete Genome Sequencing and Assembly
2.6. Phylogenomic Comparison and Tree
2.7. Representation of the Circularized Genome, Secondary Metabolite Cluster, and Colonization-Related Genes
2.8. Data Availability
3. Results
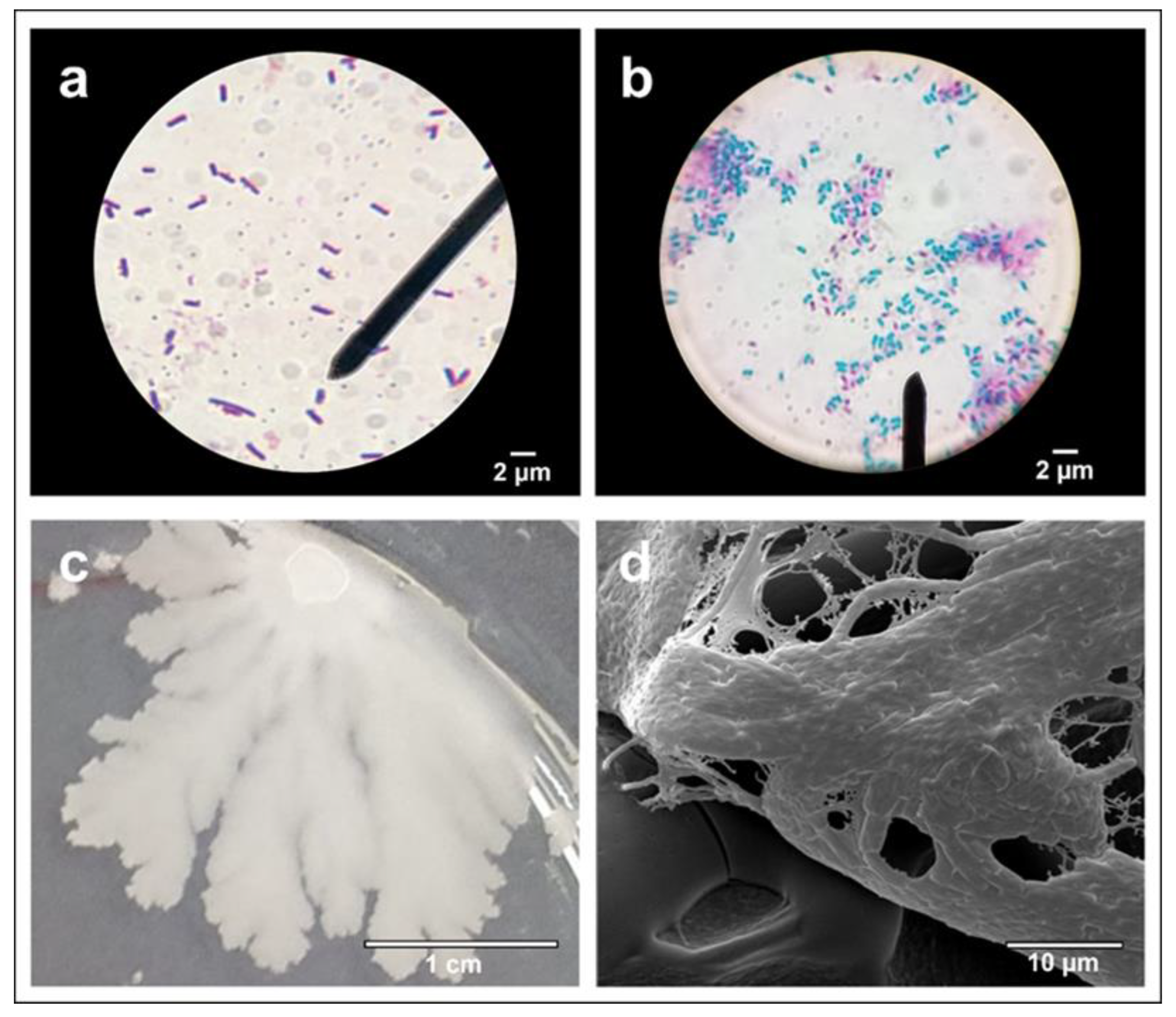
3.1. Strain Characterization
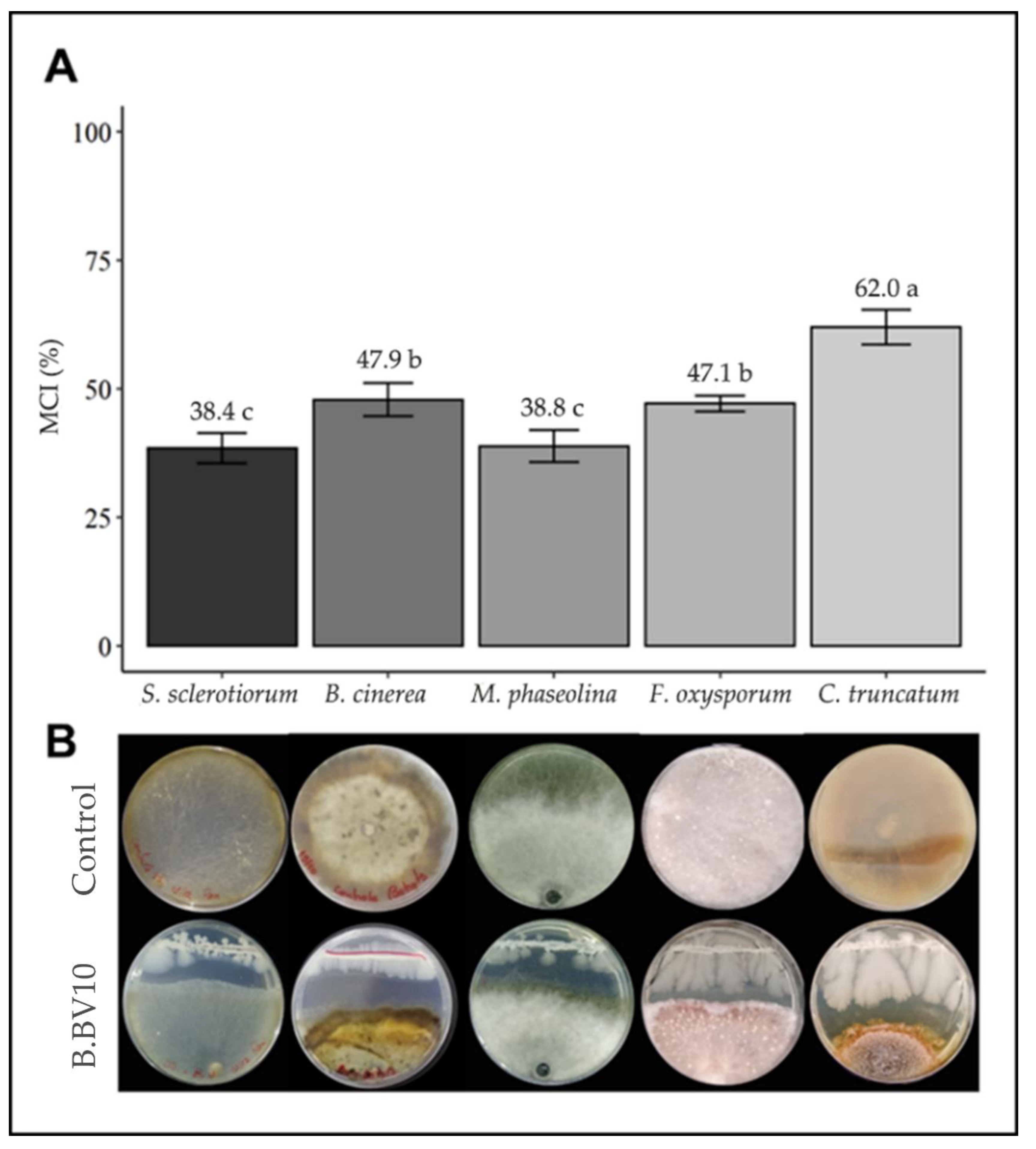
3.2. Dual Culture Assay
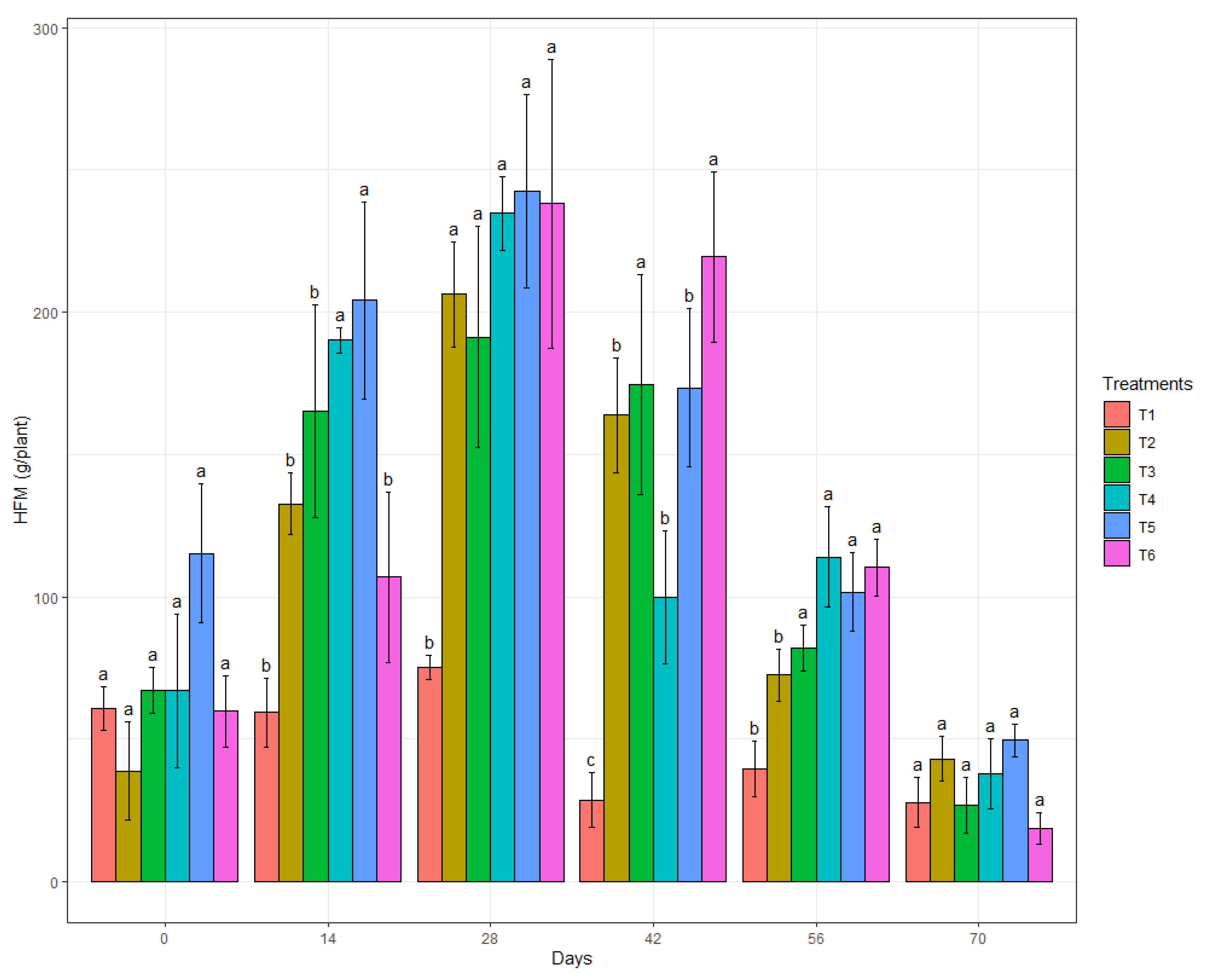
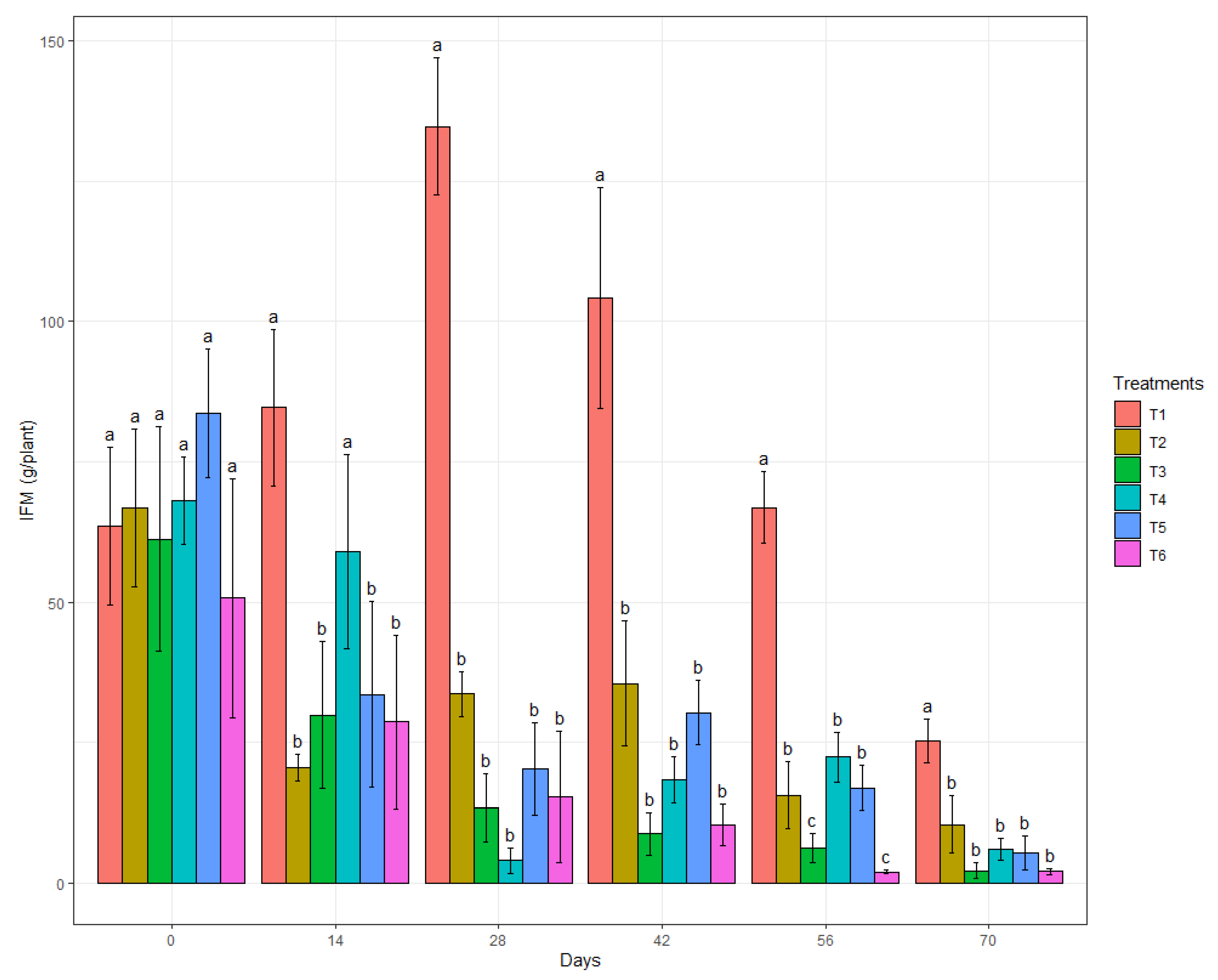
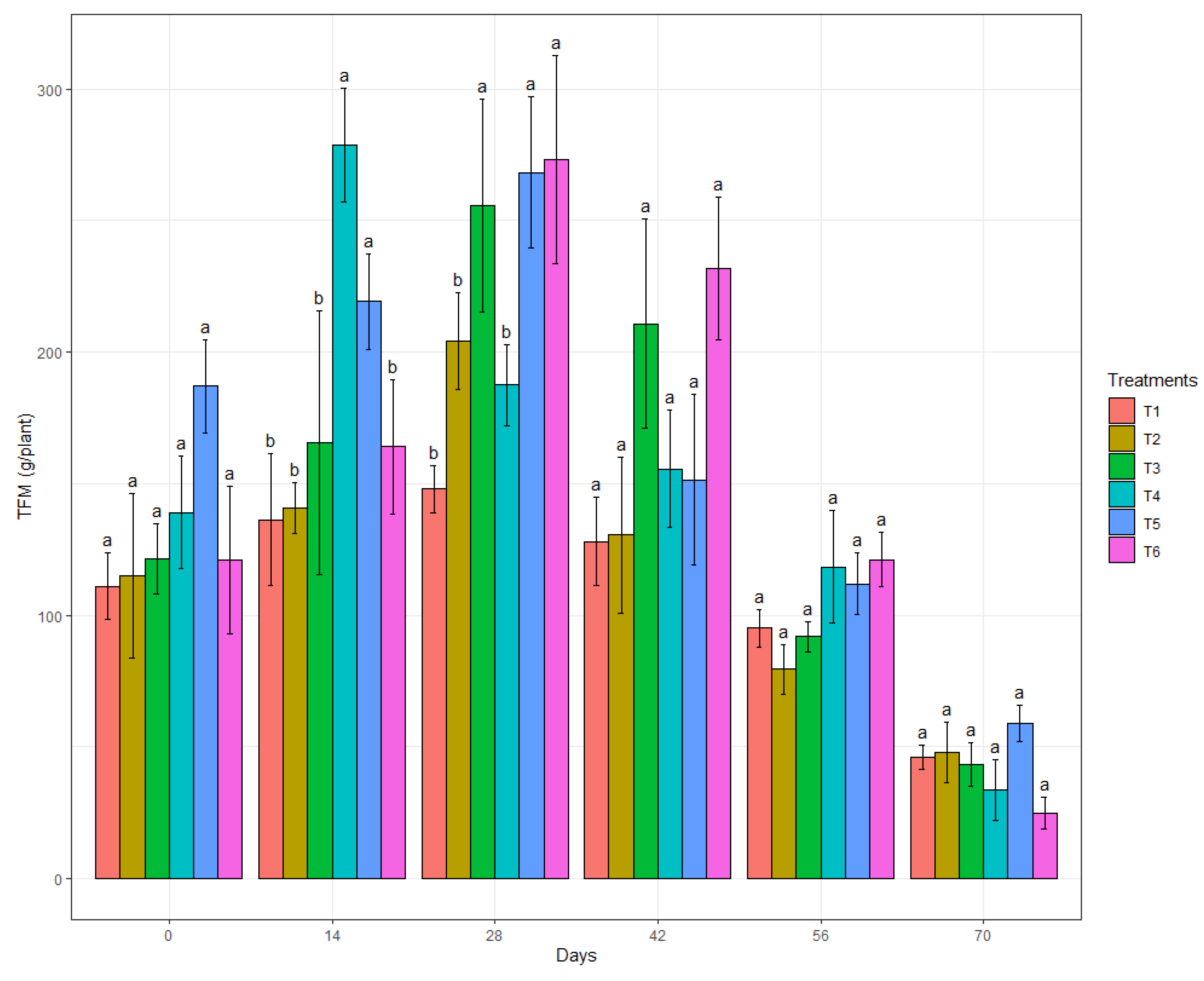
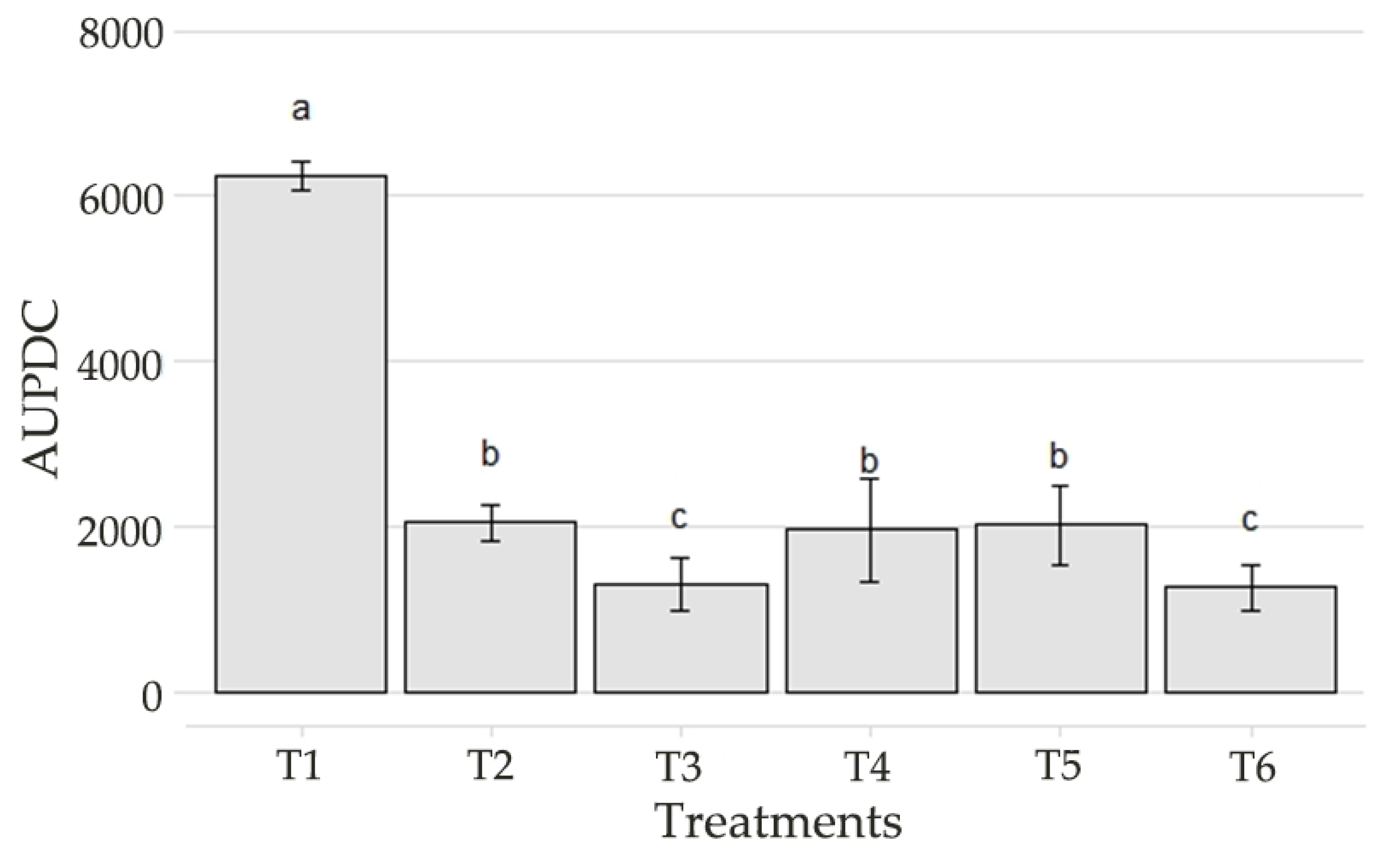
3.3. In Vivo Biocontrol
3.4. Complete Genome Sequencing and Assembly
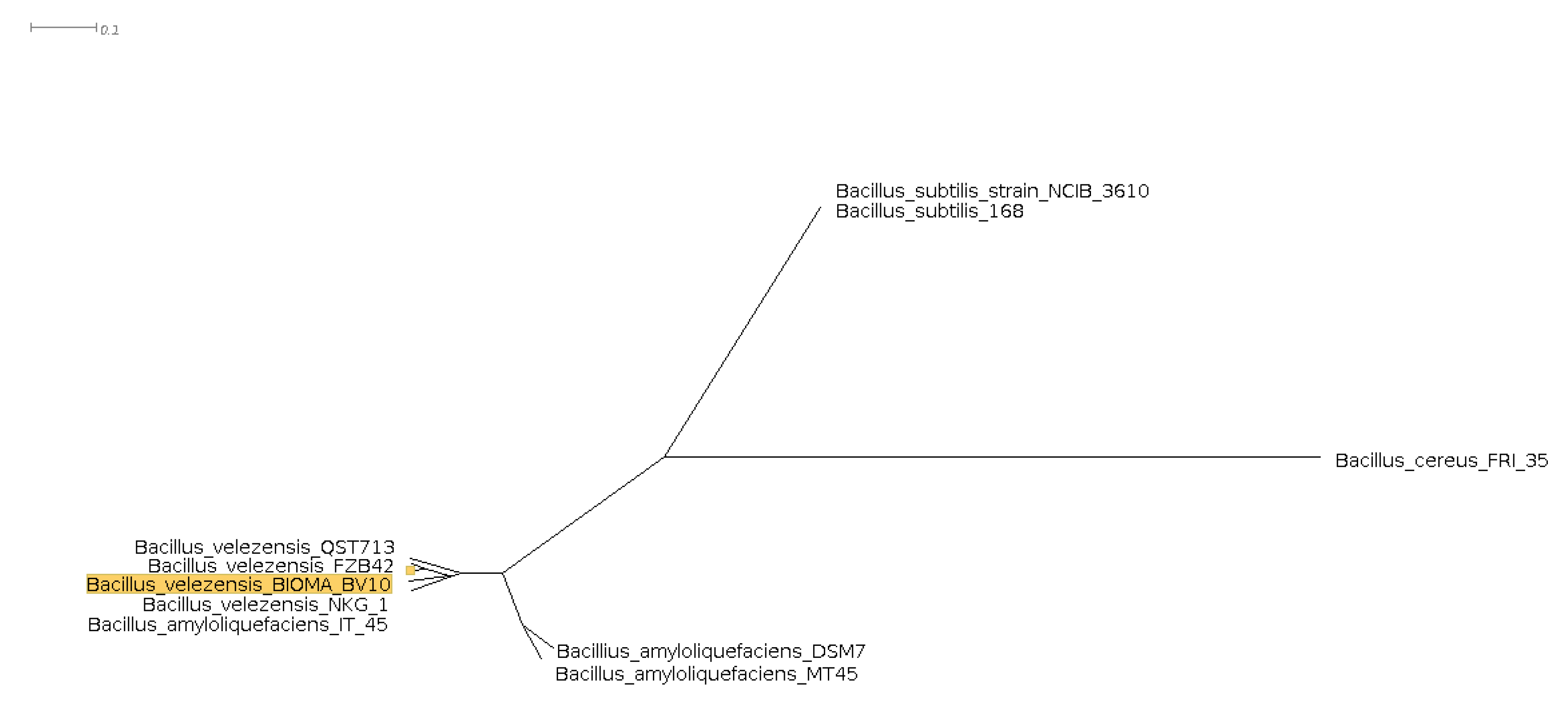
3.5. Phylogenomic Comparison and Tree
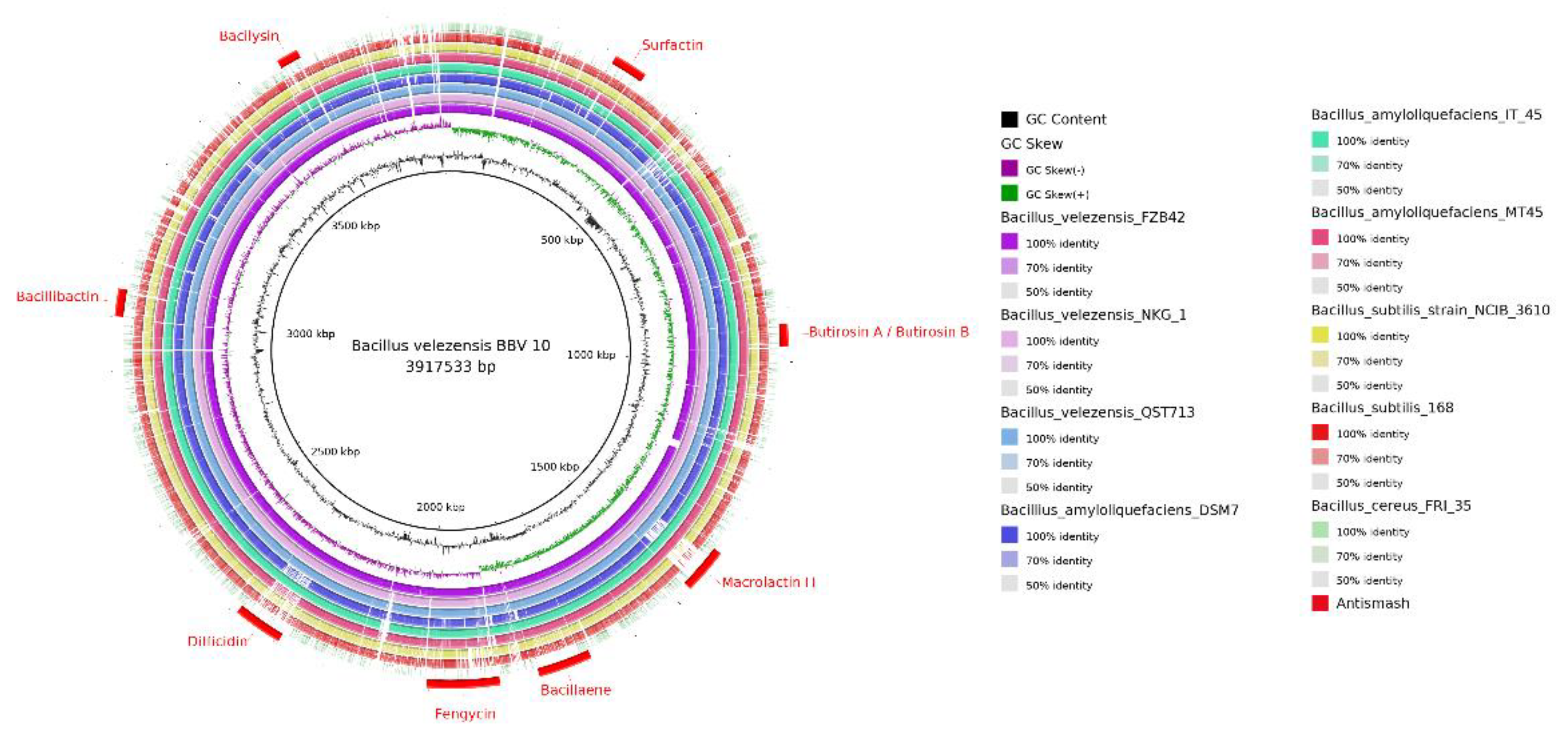
3.6. Representation of the Circularized Genome, Secondary Metabolite Cluster, and Colonization-Related Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Numan, M.; Bashir, S.; Khan, Y.; Mumtaz, R.; Shinwari, Z.K.; Khan, A.L.; Khan, A.; AL-Harrasi, A. Plant growth promoting bacteria as an alternative strategy for salt tolerance in plants: A review. Microbiol. Res. 2018, 209, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Fan, B.; Blom, J.; Klenk, H.P.; Borriss, R. Bacillus amyloliquefaciens, Bacillus velezensis, and Bacillus siamensis form an “operational group B. amyloliquefaciens” within the B. subtilis species complex. Front. Microbiol. 2017, 8, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, S.; Munir, S.; He, P.; Li, Y.; Wu, Y.; Li, X.; Kong, B.; He, P.; He, Y. Bacillus subtilis L1-21 as a biocontrol agent for postharvest gray mold of tomato caused by Botrytis cinerea. Biol. Control 2021, 157, 104568. [Google Scholar] [CrossRef]
- Qu, H.; Zhao, L.; Zhao, F.; Liu, Y.; Yang, Z. Biocontrol of gray mold decay in pear by Bacillus amyloliquefaciens strain BA3 and its effect on postharvest quality parameters. Pol. J. Microbiol. 2016, 65, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Kurniawan, O.; Wilson, K.; Mohamed, R.; Avis, T.J. Bacillus and Pseudomonas spp. provide antifungal activity against gray mold and Alternaria rot on blueberry fruit. Biol. Control 2018, 126, 136–141. [Google Scholar] [CrossRef]
- Zhao, X.; Kuipers, O.P. Identification and classification of known and putative antimicrobial compounds produced by a wide variety of Bacillales species. BMC Genom. 2016, 17, 882. [Google Scholar] [CrossRef] [Green Version]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the antimicrobial compounds produced by members of the Bacillus subtilis group. Front. Microbiol. 2019, 10, 302. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Medema, M.H.; Kazempour, D.; Fischbach, M.A.; Breitling, R.; Takano, E.; Weber, T. antiSMASH 2.0—A versatile platform for genome mining of secondary metabolite producers. Nucleic Acids Res. 2013, 41, W204–W212. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Medema, M.H.; Kottmann, R.; Lee, S.Y.; Weber, T. The antiSMASH database, a comprehensive database of microbial secondary metabolite biosynthetic gene clusters. Nucleic Acids Res. 2017, 45, D555–D559. [Google Scholar] [CrossRef]
- Fan, B.; Wang, C.; Song, X.; Ding, X.; Wu, L.; Wu, H.; Gao, X.; Borriss, R. Bacillus velezensis FZB42 in 2018: The Gram-positive model strain for plant growth promotion and biocontrol. Front. Microbiol. 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baptista, J.P.; Sanches, P.P.; Teixeira, G.M.; Morey, A.T.; Tavares, E.R.; Yamada-Ogatta, S.F.; da Rocha, S.P.D.; Hungria, M.; Ribeiro, R.A.; Balbi-Peña, M.I.; et al. Complete genome sequence of Bacillus velezensis LABIM40, an effective antagonist of fungal plant pathogens. Genome Announc. 2018, 6, e00595-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saberi Riseh, R.; Skorik, Y.A.; Thakur, V.K.; Moradi Pour, M.; Tamanadar, E.; Noghabi, S.S. Encapsulation of plant biocontrol bacteria with alginate as a main polymer material. Int. J. Mol. Sci. 2021, 22, 11165. [Google Scholar] [CrossRef]
- Teixeira, G.M.; Mosela, M.; Nicoletto, M.L.A.; Ribeiro, R.A.; Hungria, M.; Youssef, K.; Higashi, A.Y.; Mian, S.; Ferreira, A.S.; Gonçalves, L.S.A.; et al. Genomic insights into the antifungal activity and plant growth-promoting ability in Bacillus velezensis CMRP 4490. Front. Microbiol. 2021, 11, 618415. [Google Scholar] [CrossRef] [PubMed]
- Wrege, M.S.; Steinmetz, S.; Reisser Junior, C.; Almeida, I.R. Atlas Climático da Região Sul do Brasil: Estados do Paraná, Santa Catarina e Rio Grande do Sul, 2nd ed.; Wrege, M.S., Steinmetz, S., Reisser Junior, C., Almeida, I.R., Eds.; Embrapa: Brasília, Brazil, 2012; Volume 1. [Google Scholar]
- Santos, H.G.; Jacomine, P.K.T.; Anjos, L.H.C.; Oliveira, V.A.; Lumbreras, J.F.; Coelho, M.R.; Almeida, J.A.; Araujo Filho, J.C.; Oliveira, J.B.; Cunha, T.J.F. Sistema Brasileiro de Classificação de Solos, 5th ed.; Embrapa, Ed.; Embrapa: Brasília, Brazil, 2018; Volume 1. [Google Scholar]
- Alves, K.S.; del Ponte, E.M. Analysis and simulation of plant disease progress curves in R: Introducing the epifitter package. Phytopathol. Res. 2021, 3, 22. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data 2014. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Leung, H.C.M.; Yiu, S.M.; Chin, F.Y.L. IDBA-UD: A de novo assembler for single-cell and metagenomic sequencing data with highly uneven depth. Bioinformatics 2012, 28, 1420–1428. [Google Scholar] [CrossRef] [Green Version]
- Galardini, M.; Biondi, E.G.; Bazzicalupo, M.; Mengoni, A. CONTIGuator: A bacterial genomes finishing tool for structural insights on draft genomes. Source Code Biol. Med. 2011, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y.; et al. SOAPdenovo2: An empirically improved memory-efficient short-read de novo assembler. Gigascience 2012, 1, 18. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.; Ouk Kim, Y.; Park, S.C.; Chun, J. OrthoANI: An improved algorithm and software for calculating average nucleotide identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acids Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef] [PubMed]
- Ågren, J.; Sundström, A.; Håfström, T.; Segerman, B. Gegenees: Fragmented alignment of multiple genomes for determining phylogenomic distances and genetic signatures unique for specified target groups. PLoS ONE 2012, 7, e39107. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. AntiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Alikhan, N.-F.; Petty, N.K.; ben Zakour, N.L.; Beatson, S.A. BLAST ring image generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Stülke, J. SubtiWiki in 2018: From genes and proteins to functional network annotation of the model organism Bacillus subtilis. Nucleic Acids Res. 2018, 46, D743–D748. [Google Scholar] [CrossRef]
- Wu, L.; Wu, H.J.; Qiao, J.; Gao, X.; Borriss, R. Novel routes for improving biocontrol activity of Bacillus based bioinoculants. Front. Microbiol. 2015, 6, 1395. [Google Scholar] [CrossRef] [Green Version]
- Lemon, K.P.; Earl, A.M.; Vlamakis, H.C.; Aguilar, C.; Kolter, R. Biofilm development with an emphasis on Bacillus subtilis. Curr. Top Microbiol. Immunol. 2008, 322, 1–16. [Google Scholar] [CrossRef]
- Arnaouteli, S.; Bamford, N.C.; Stanley-Wall, N.R.; Kovács, Á.T. Bacillus subtilis biofilm formation and social interactions. Nat. Rev. Microbiol. 2021, 19, 600–614. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.B.; Bertolli, S.K.; Mougous, J.D. The central role of interbacterial antagonism in bacterial life. Curr. Biol. 2020, 30, R1203–R1214. [Google Scholar] [CrossRef] [PubMed]
- Kautsar, S.A.; Blin, K.; Shaw, S.; Navarro-Muñoz, J.C.; Terlouw, B.R.; van der Hooft, J.J.J.; van Santen, J.A.; Tracanna, V.; Suarez Duran, H.G.; Pascal Andreu, V.; et al. MIBiG 2.0: A repository for biosynthetic gene clusters of known function. Nucleic Acids Res. 2020. 48, D454–D458. [CrossRef] [Green Version]
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; van Kan, J.A.L. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef]
- Simpson, D. The economic importance of strawberry crops. In The Genomes of Rosaceous Berries and Their Wild Relatives. Compendium of Plant Genomes 2018; Hytönen, T., Graham, J., Harrison, R., Eds.; Springer: Cham, Germany, 2018. [Google Scholar] [CrossRef]
- Balderas-Ruíz, K.A.; Gómez-Guerrero, C.I.; Trujillo-Roldán, M.A.; Valdez-Cruz, N.A.; Aranda-Ocampo, S.; Juárez, A.M.; Leyva, E.; Galindo, E.; Serrano-Carreón, L. Bacillus velezensis 83 increases productivity and quality of tomato (Solanum lycopersicum L.): Pre and postharvest assessment. Curr. Res. Microb. Sci. 2021, 2, 100076. [Google Scholar] [CrossRef]
- Jiang, C.-H.; Liao, M.-J.; Wang, H.-K.; Zheng, M.-Z.; Xu, J.-J.; Guo, J.-H. Bacillus velezensis, a potential and efficient biocontrol agent in control of pepper gray mold caused by Botrytis cinerea. Biol. Control 2018, 126, 147–157. [Google Scholar] [CrossRef]
- Toral, L.; Rodríguez, M.; Béjar, V.; Sampedro, I. Crop protection against Botrytis cinerea by rhizosphere biological control agent Bacillus velezensis XT1. Microorganisms 2020, 8, 992. [Google Scholar] [CrossRef] [PubMed]
- Amarouchi, Z.; Esmaeel, Q.; Sanchez, L.; Jacquard, C.; Hafidi, M.; Vaillant-Gaveau, N.; Ait Barka, E. Beneficial microorganisms to control the gray mold of grapevine: From screening to mechanisms. Microorganisms 2021, 9, 1386. [Google Scholar] [CrossRef] [PubMed]
- Toral, L.; Rodríguez, M.; Béjar, V.; Sampedro, I. Antifungal activity of lipopeptides from Bacillus XT1 CECT 8661 against Botrytis cinerea. Front. Microbiol. 2018, 9, 1315. [Google Scholar] [CrossRef]
- Pandin, C.; le Coq, D.; Deschamps, J.; Védie, R.; Rousseau, T.; Aymerich, S.; Briandet, R. Complete genome sequence of Bacillus velezensis QST713: A biocontrol agent that protects Agaricus bisporus crops against the Green Mould Disease. J. Biotechnol. 2018, 278, 10–19. [Google Scholar] [CrossRef]
- Chen, X.H.; Koumoutsi, A.; Scholz, R.; Schneider, K.; Vater, J.; Süssmuth, R.; Piel, J.; Borriss, R. Genome analysis of Bacillus amyloliquefaciens FZB42 reveals its potential for biocontrol of plant pathogens. J. Biotechnol. 2009, 140, 27–37. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | GenBank Access Number | GC% |
|---|---|---|
| B. velezensis FZB42 | CP000560.2 | 46.50 |
| B. velezensis NKG1 | CP024203.1 | 46.30 |
| B. velezensis QST713 | CP025079 | 45.90 |
| B. amyloliquefaciens IT45 | CP004065.1 | 46.60 |
| B. amyloliquefaciens DSM7 | NC0145511 | 46.10 |
| B. amyloliquefaciens MT45 | CP0112521 | 46.10 |
| B. subtilis 168 | NC0009643 | 43.50 |
| B. subtilis NCIB 3610 | CP020102.1 | 43.50 |
| B. cereus FRI35 | CP003747.1 | 35.45 |
| Genome Size | 3,917,533 bp |
|---|---|
| GC content | 46.60% |
| Plasmids | 0 |
| Genes (Total) | 3838 |
| Genes (Coding) | 3676 |
| rRNA operons | 9 |
| tRNA operons | 7 |
| ncRNA | 5 |
| Alignment rate | 98.27% |
| GenBank access | NZ_CP059318.1 (https://www.ncbi.nlm.nih.gov/nuccore/NZ_CP059318.1, accessed on 1 August 2022) |
| BioProject | PRJNA224116 (https://www.ncbi.nlm.nih.gov/bioproject/PRJNA224116, accessed on 1 August 2022) |
| BioSample | SAMN15484393 (https://www.ncbi.nlm.nih.gov/biosample/SAMN15484393, accessed on 1 August 2022) |
| Strain | OrthoANI (%) | dDDH | Distance | GC |
|---|---|---|---|---|
| B. velezensis FZB42 | 99.26 | 94.30 | 0.03 | 0.10 |
| B. velezensis NKG-1 | 99.06 | 91.70 | 0.07 | 0.27 |
| B. velezensis QST713 | 98.43 | 87.40 | 0.09 | 0.67 |
| B. amyloliquefaciens IT45 | 97.72 | 80.10 | 0.07 | 0.04 |
| B. amyloliquefaciens MT45 | 94.15 | 55.60 | 0.15 | 0.49 |
| B. amyloliquefaciens DSM7 | 94.04 | 55.60 | 0.17 | 0.49 |
| B. subtilis 168 | 77.00 | 20.80 | 0.57 | 3.06 |
| B. subtilis NCIB3610 | 77.00 | 20.80 | 0.57 | 3.06 |
| B. cereus FRI 35 | 66.72 | 20.80 | 0.99 | 10.98 |
| Region | Type | From | To | Most Similar Know Cluster |
|---|---|---|---|---|
| 1 | lanthipeptide class II | 97,265 | 120,306 | |
| 2 | NRPS | 318,617 | 383,476 | Surfactin |
| 3 | PKS-like | 929,684 | 970,928 | Butyrosin A/Butyrosin B |
| 4 | Terpene | 1,056,551 | 1,073,722 | |
| 5 | TransAT-PKS | 1,380,701 | 1,467,090 | Macrolactin H |
| 6 | TransAT-PKS, T3PKS, NRPS | 1,690,625 | 1,792,535 | Bacillaene |
| 7 | NRPS, TransAT-PKS, Betalactone | 1,866,712 | 2,004,072 | Fengycin |
| 8 | Terpene | 2,031,080 | 2,052,963 | |
| 9 | T3PKS | 2,123,567 | 2,164,667 | |
| 10 | TransAT-PKS | 2,292,697 | 2,386,483 | Difficidin |
| 11 | NRPS | 3,002,179 | 3,053,970 | Bacillibactin |
| 12 | Other | 3,580,766 | 3,622,184 | Bacilysin |
| Genes | Identity (%) | Genome Position in B. Velezensis B.BV10 (bp) | Described Function According to Subtwiki [30] | |
|---|---|---|---|---|
| abrB | 91.379 | 45,889 | 45,600 | Transcriptional regulator of transition-state genes. |
| comA | 80.380 | 2,996,026 | 2,995,395 | Regulation of genetic competence and quorum sensing. |
| degQ | 86.765 | 2,999,820 | 2,999,685 | Stimulates production of degradative enzymes and extracellular poly-gamma-glutamate; stimulates phosphorylation of DegU by DegS. |
| degU | 86.377 | 3,384,604 | 3,383,915 | Two-component response regulator; regulation of degradative enzyme expression, genetic competence, biofilm formation, and capsule biosynthesis (together with SwrA); non-phosphorylated DegU is required for swarming motility. |
| epsA | 99.294 | 3,289,519 | 3,288,812 | Extracellular polysaccharide synthesis; putative transmembrane modulator of EpsB activity; might activate EpsB autophosphorylation and substrate phosphorylation. |
| epsB | 99.853 | 3,288,806 | 3,288,126 | Extracellular polysaccharide synthesis; protein tyrosine kinase; phosphorylation of EpsE. |
| epsC | 98.941 | 3,287,880 | 3,286,087 | UDP-N-acetylglucosamine 4,6-dehydratase; required for extracellular polysaccharide synthesis; this gene is inactive in B. subtilis 168. |
| epsD | 99.649 | 3,286,071 | 3,284,932 | Extracellular polysaccharide synthesis. |
| epsE | 100.000 | 3,284,935 | 3,284,093 | Inhibitor of motility and glycosyltransferase required for EPS biosynthesis. |
| epsF | 99.296 | 3,284,100 | 3,282,964 | Similar to glycosyltransferase. |
| epsG | 99.547 | 3,282,960 | 3,281,857 | Extracellular polysaccharide synthesis. |
| epsH | 99.037 | 3,281,838 | 3,280,801 | Undecaprenyl (UnDP) priming UDP-N-acetyl-glucosamine transferase; synthesis of extracellular poly-N-acetylglucosamine. |
| epsI | 98.236 | 3,280,796 | 3,279,720 | Glycosyltransferase; synthesis of extracellular poly-N-acetylglucosamine. |
| epsJ | 98.454 | 3,279,723 | 3,278,689 | UDP-N-acetyl-glucosamine transferase; synthesis of extracellular poly-N-acetylglucosamine. |
| epsK | 99.605 | 3,278,692 | 3,277,175 | Export of extracellular poly-N-acetylglucosamine. |
| epsL | 100.000 | 3,277,178 | 3,276,570 | Similar to UDP-galactose phosphate transferase; extracellular polysaccharide synthesis. |
| epsM | 99.691 | 3,276,573 | 3,275,926 | UDP-2,4,6-trideoxy-2-acetamido-4-amino glucose acetyltransferase; extracellular polysaccharide synthesis. |
| epsN | 99.062 | 3,275,921 | 3,274,749 | UDP-2,6-dideoxy 2-acetamido 4-keto glucose aminotransferase; required for extracellular polysaccharide synthesis. |
| epsO | 99.793 | 3,274,770 | 3,273,805 | Similar to pyruvyltransferase; extracellular polysaccharide synthesis. |
| galE1 | 99.899 | 1,173,111 | 1,172,119 | UDP glucose 4-epimerase. |
| kinA | 77.669 | 1,349,359 | 1,351,179 | Two-component sensor kinase; phosphorylates Spo0F; part of the phosphorelay. |
| motA | 79.044 | 1,317,745 | 1,316,930 | H+-coupled MotA-MotB flagellar stator. |
| motB | 76.305 | 1,316,958 | 1,316,216 | H+-coupled MotA-MotB flagellar stator. |
| remA | 97.778 | 1,565,578 | 1,565,847 | Transcriptional regulator of the extracellular matrix genes; acts in parallel with SinR, AbrB, and DegU. |
| sigD | 87.451 | 1,640,430 | 1,641,194 | RNA polymerase sigma fator SigD. |
| sigH | 87.931 | 123,748 | 124,385 | RNA polymerase sigma factor SigH; not fully active in laboratory strains due to a mutation (V117A). |
| sigW | 83.156 | 200,648 | 201,211 | RNA polymerase ECF-type sigma factor SigW; required for the adaptation to membrane-active agents; activated by alkaline shock and polymyxin B, vancomycin, cephalosporin C, D-cycloserine, and triton X-100. |
| sinl | 79.096 | 2,446,052 | 2,446,225 | Antagonist of SinR; drives SlrR from the SlrR (LOW) to the SlrR (HIGH) state. |
| sinR | 97.321 | 2,446,259 | 2,446,594 | Transcriptional regulator (Xre family) of post-exponential-phase response genes. |
| sipW | 100.000 | 2,448,076 | 2,447,492 | Bifunctional signal peptidase I that control surface-adhered biofilm formation and processes TasA and TapA. |
| spoOA | 99.251 | 2,412,896 | 2,412,096 | Phosphorelay regulator; initiation of sporulation; coordinates DNA replication and initiation of sporulation by binding to sites close to the oriC. |
| srfAA | 78.807 | 356,838 | 358,849 | Surfactin synthetase/competence. |
| srfAB | 79.825 | 1,892,212 | 1,892,099 | Surfactin synthetase/competence. |
| srfAC | 87.099 | 360,085 | 363,881 | Surfactin synthetase/competence. |
| swrA | 83.481 | 3,360,036 | 3,359,700 | Master activator of flagellar biosynthesis; modulator of DegU activity; converts DegU-P from a repressor to an activator of the fla-che operon; enhances sigD transcription; controls the number of flagellar basal bodies; inactive pseudogene in strain 168. |
| swrB | 81.006 | 1,641,222 | 1,641,400 | Control of SigD activity; required for full SigD activity; activates the flagellar type-III secretion export apparatus by the membrane protein FliP. |
| swrC | 79.906 | 693,419 | 696,562 | Similar to acriflavin resistance protein. |
| tasA | 99.491 | 2,447,427 | 2,446,642 | Major component of the biofilm matrix; forms amyloid fibers. |
| yhxB | 99.541 | 924,407 | 926,149 | Alpha-phosphoglucomutase; required for UDP-glucose synthesis; inhibits FtsZ ring assembly (indirect effect due to a defect in UDP-glucose synthesis); possesses secondary phosphoglucosamine mutase activity. |
| yqxM | 99.554 | 2,448,719 | 2,448,048 | TasA anchoring/assembly protein. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertê, R.; Teixeira, G.M.; de Oliveira, J.P.; Nicoletto, M.L.A.; da Silva, D.V.; de Godoy, G.G.; Sanches, D.S.; de Resende, J.T.V.; Pereira, U.d.P.; Nunes da Rocha, U.; et al. Genome Mining Reveals High Biosynthetic Potential of Biocontrol Agent Bacillus velezensis B.BV10. Genes 2022, 13, 1984. https://doi.org/10.3390/genes13111984
Bertê R, Teixeira GM, de Oliveira JP, Nicoletto MLA, da Silva DV, de Godoy GG, Sanches DS, de Resende JTV, Pereira UdP, Nunes da Rocha U, et al. Genome Mining Reveals High Biosynthetic Potential of Biocontrol Agent Bacillus velezensis B.BV10. Genes. 2022; 13(11):1984. https://doi.org/10.3390/genes13111984
Chicago/Turabian StyleBertê, Rosiana, Gustavo Manoel Teixeira, João Paulo de Oliveira, Maria Luiza Abreu Nicoletto, Daniel Vieira da Silva, Guilherme Gonçalves de Godoy, Danilo Sipoli Sanches, Juliano Tadeu Vilela de Resende, Ulisses de Padua Pereira, Ulisses Nunes da Rocha, and et al. 2022. "Genome Mining Reveals High Biosynthetic Potential of Biocontrol Agent Bacillus velezensis B.BV10" Genes 13, no. 11: 1984. https://doi.org/10.3390/genes13111984