

miR-125-3p and miR-276b-3p Regulate the Spermatogenesis of Bactrocera dorsalis by Targeting the orb2 Gene

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rearing of Insects
2.2. Selection of Target Gene
2.3. miRNA Target Site Prediction
2.4. dsRNA, Agomir, and Antagomir Preparation
2.5. Dual Luciferase Reporter Assay
2.6. RNA Isolation and Quantitative Real-Time PCR
2.7. Administration of dsRNA, Agomirs, and Antagomirs to the Adult Flies
2.8. Reproductive Capacity of Male Flies
2.9. Sperm Viability Assays and Spermatozoa Counts
2.10. Data Analyses
3. Results
3.1. Gene Selection
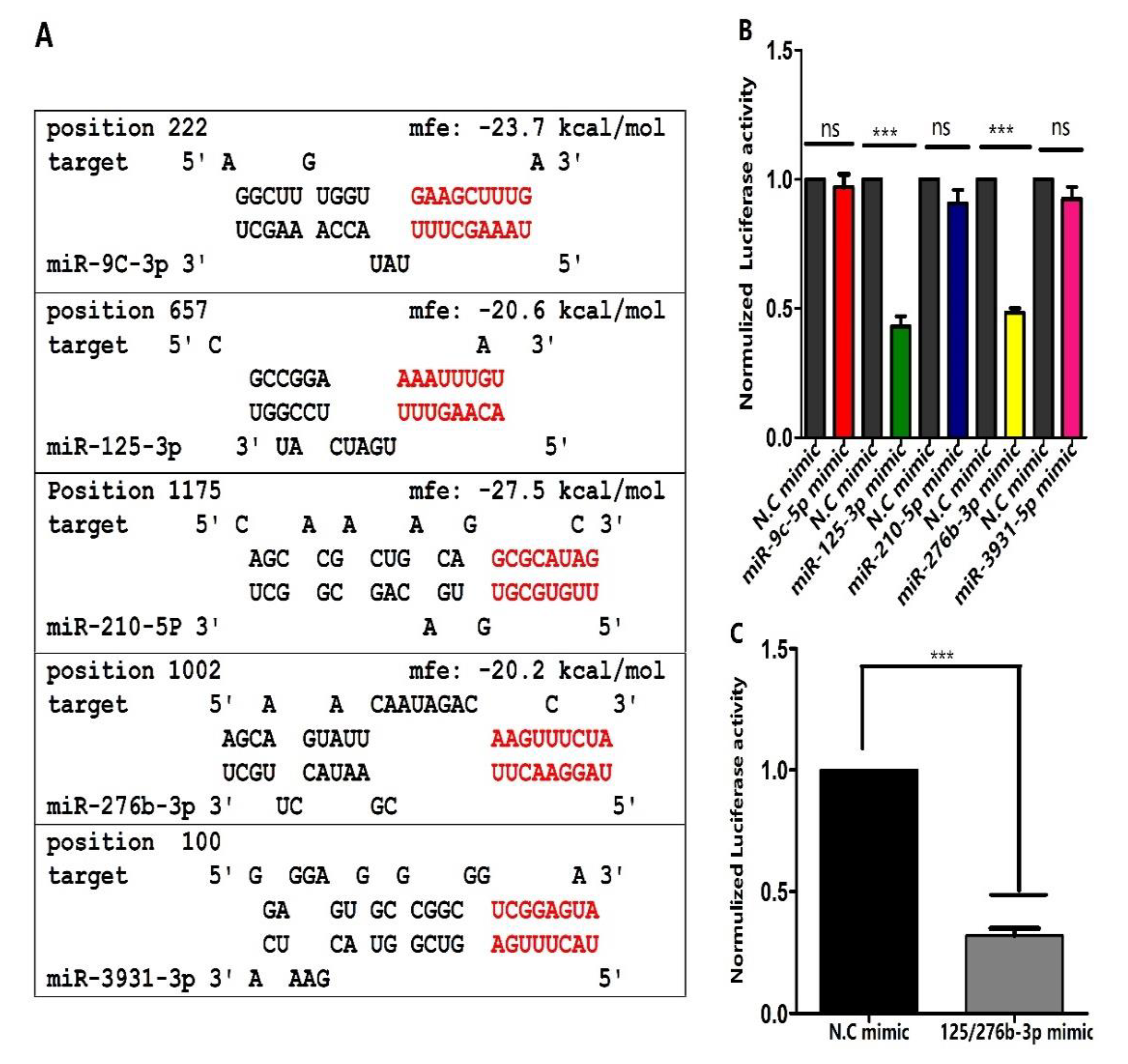
3.2. Prediction of miRNAs Targeting orb2 and Their Confirmation
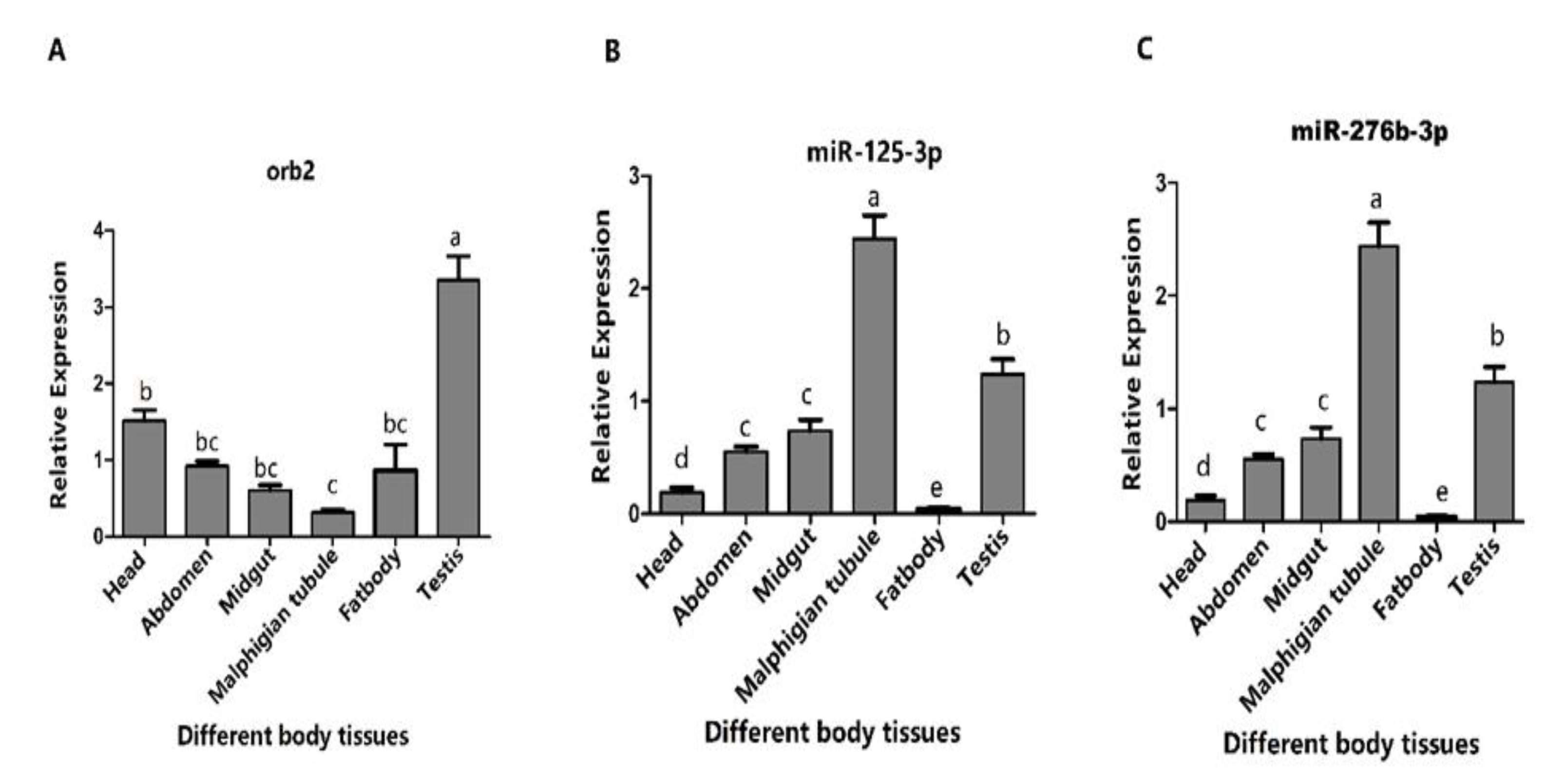
3.3. Expression Profiles of orb2 and Its Target miRNAs
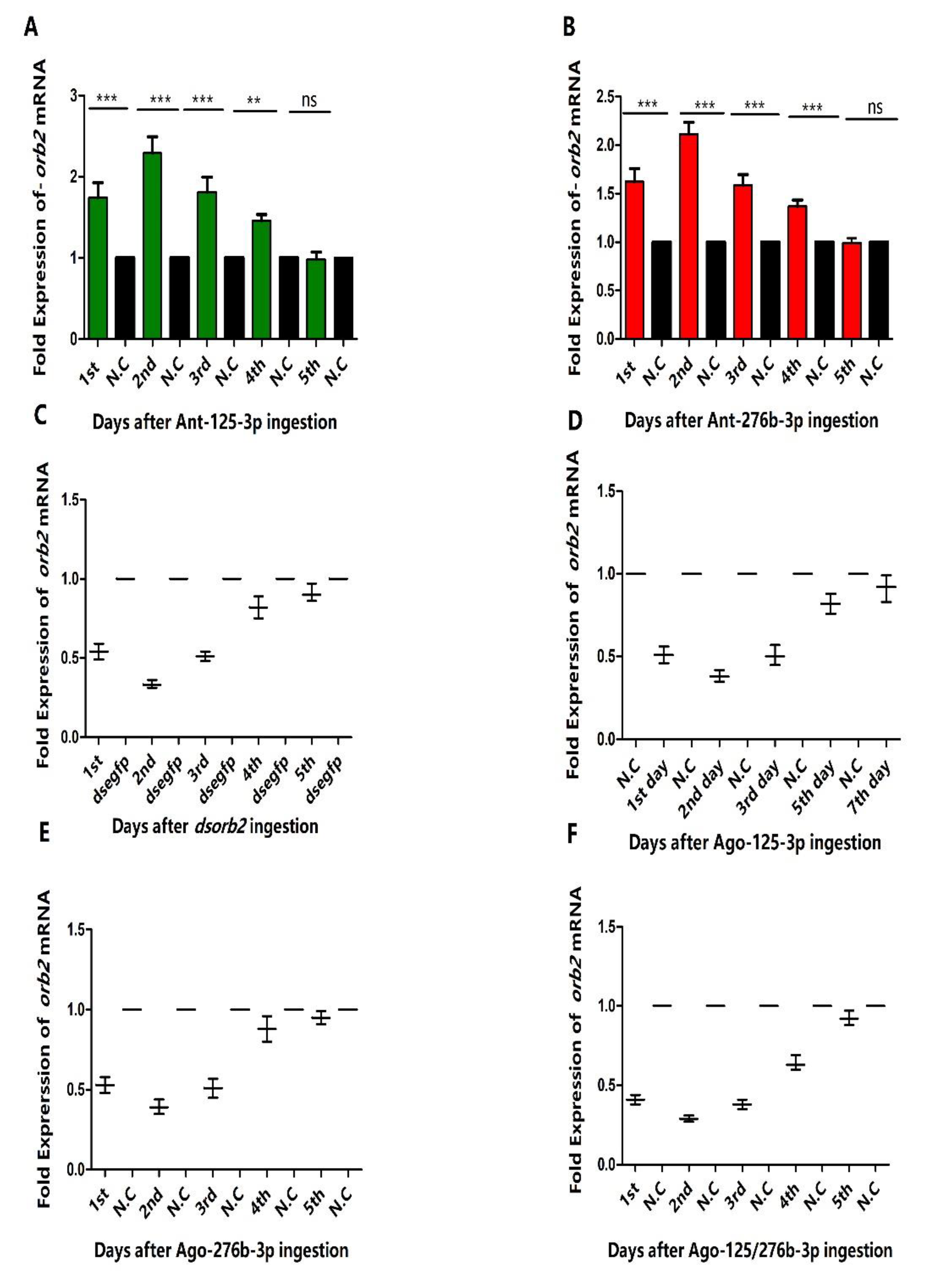
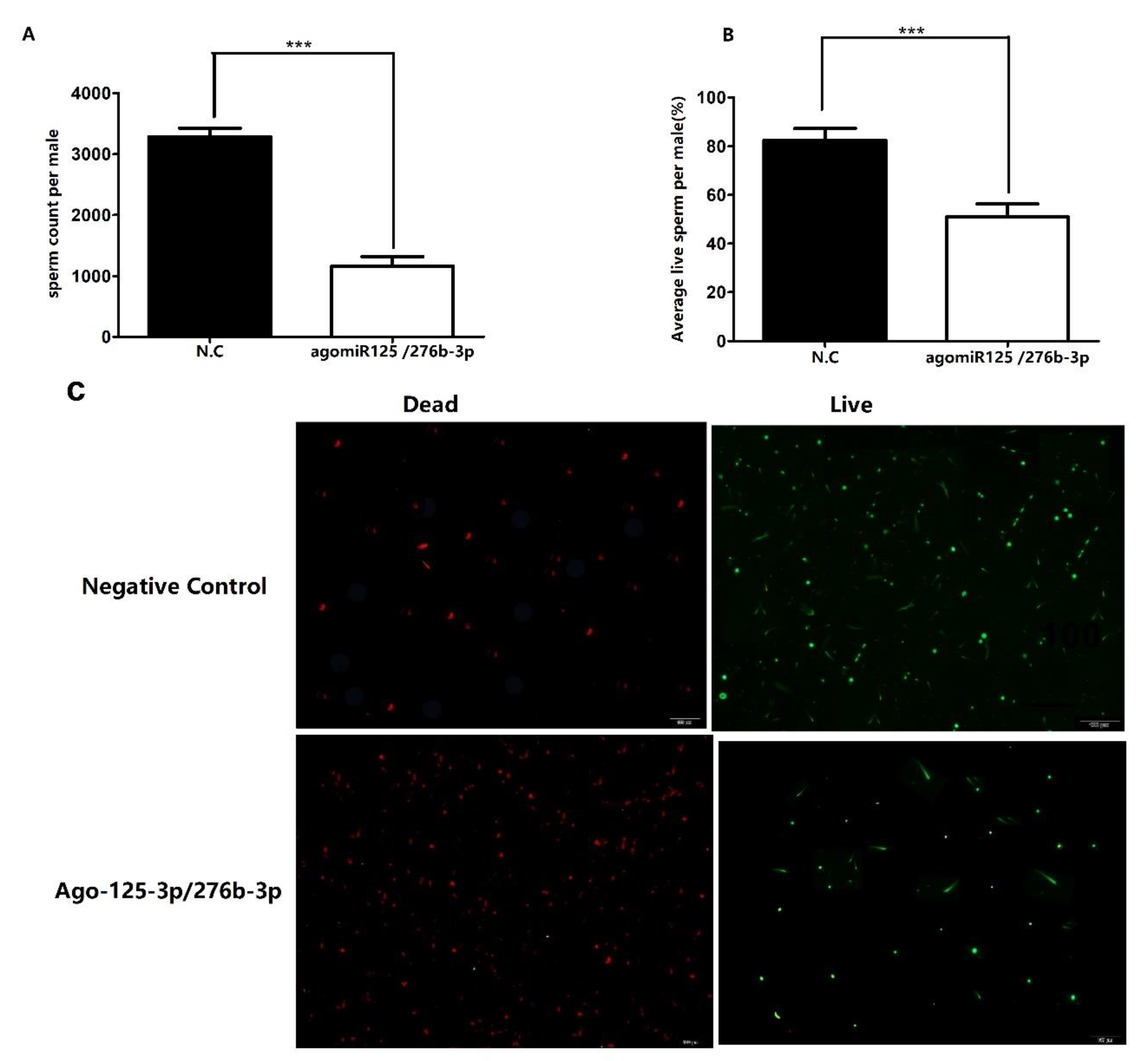
3.4. Overexpression of miR-125-3p and miR-276b-3p Impairs the Male Fertility
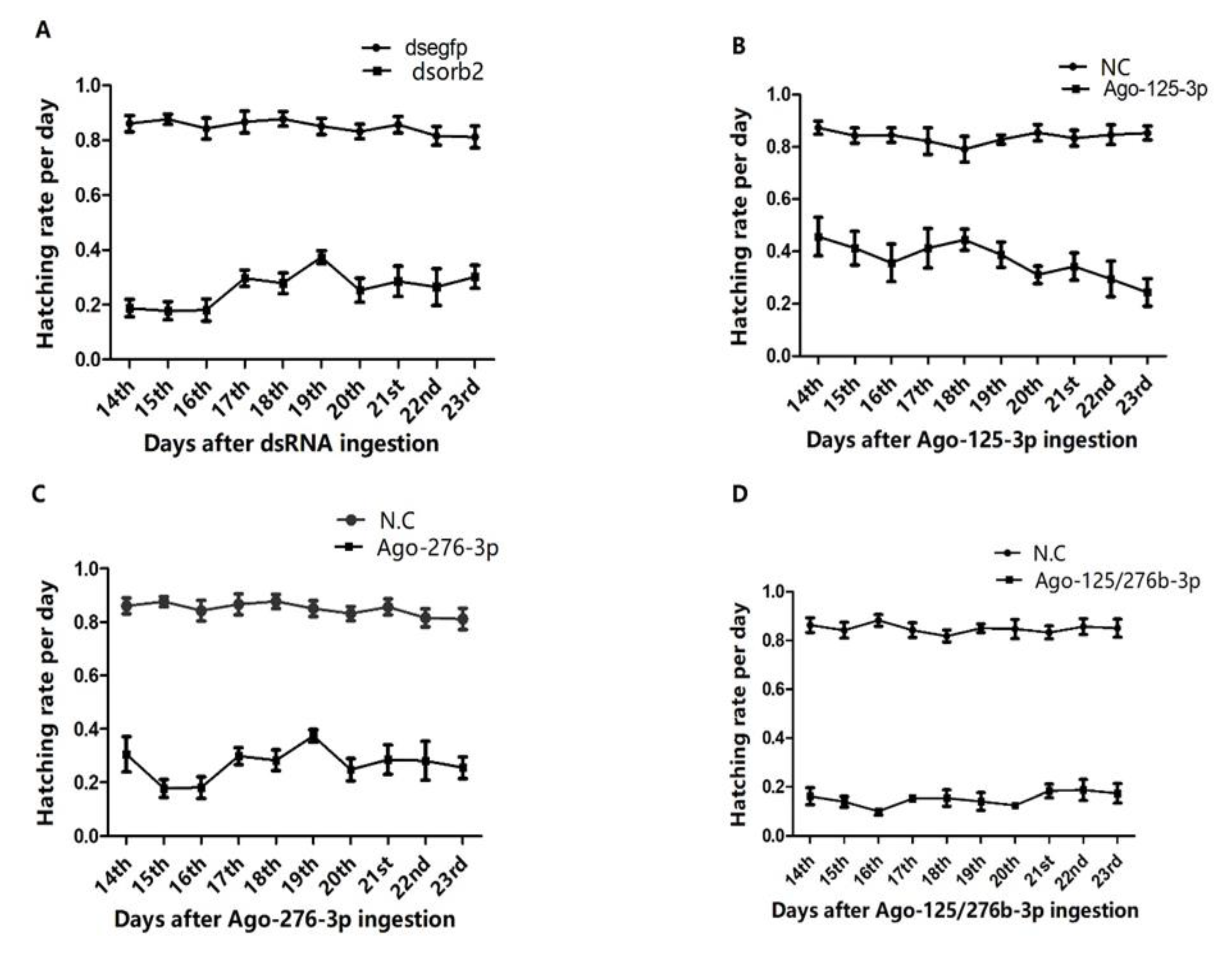
3.5. Investigation of Number of Sperms and Sperm Viability


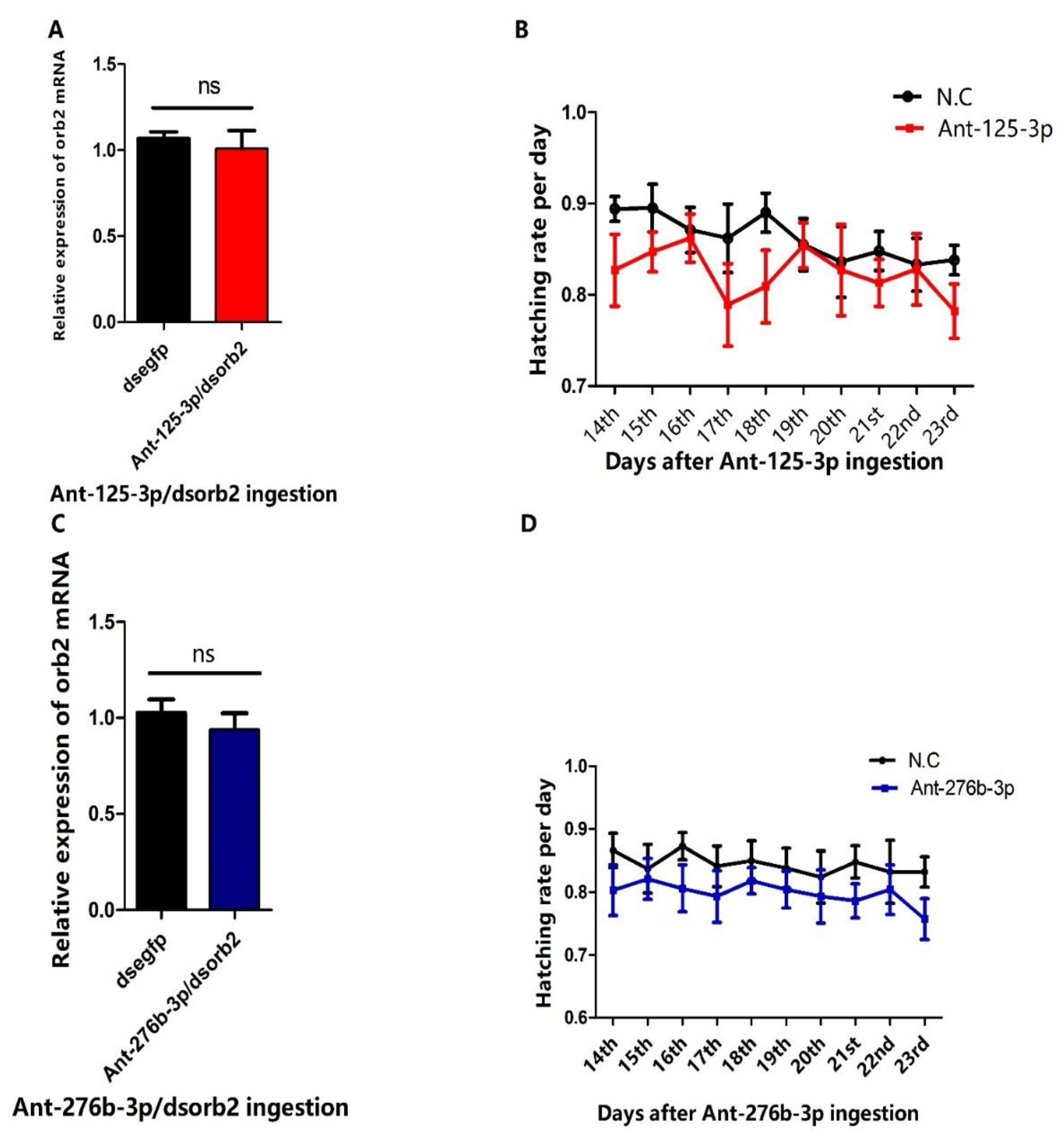
3.6. RNAi of Antagomir-125-3p and Antagomir-276b-3p Treated Individuals Rescued the Phenotype Caused by dsorb2 Treatment

4. Discussions

5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Clarke, A.R.; Armstrong, K.F.; Carmichael, A.E.; Milne, J.R.; Raghu, S.; Roderick, G.K.; Yeates, D.K. Invasive phytophagous pests arising through a recent tropical evolutionary radiation: The Bactrocera dorsalis complex of fruit flies. Annu. Rev. Entomol. 2005, 50, 293–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, I.M.; Elson-Harris, M.M. Fruit Flies of Economic Significance: Their Identification and Bionomics; CAB International, Walligford and ACIAR: Canberra, Australia, 1992. [Google Scholar]
- Allwood, A.; Chinajariyawong, A.; Kritsaneepaiboon, S.; Drew, R.; Hamacek, E.; Hancock, D.; Hengsawad, C.; Jipanin, J.; Jirasurat, M.; Krong, C.K. Host plant records for fruit Flies (Diptera: Tephritidae) in Southeast Asia. Raffles Bull. Zool. 1999, 47, 1–92. [Google Scholar]
- Alphey, N.; Bonsall, M.B. Genetics-Based Methods for Agricultural Insect Pest Management. Agric. For. Entomol. 2018, 20, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Wimmer, E.A. Eco-friendly insect management. Nat. Biotechnol. 2005, 23, 432–433. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.W.; Zheng, W.; Sohail, S.; Li, Q.; Zheng, W.; Zhang, H. A genetically enhanced sterile insect technique against the fruit fly, Bactrocera dorsalis (Hendel) by feeding adult double-stranded RNAs. Sci. Rep. 2017, 7, 4063. [Google Scholar] [CrossRef] [Green Version]
- Sohail, S.; Tariq, K.; Zheng, W.; Ali, M.W.; Peng, W.; Raza, M.F.; Zhang, H. RNAi-mediated knockdown of Tssk1 and Tektin1 genes impair male fertility in Bactrocera dorsalis. Insects 2019, 10, 164. [Google Scholar] [CrossRef] [Green Version]
- Okada, H.; Tajima, A.; Shichiri, K.; Tanaka, A.; Tanaka, K.; Inoue, I. Genome-wide expression of azoospermia testes demonstrates a specific profile and implicates ART3 in genetic susceptibility. PLoS Genet. 2008, 4, e26. [Google Scholar] [CrossRef] [Green Version]
- De Kretser, D. Local regulation of testicular function. Int. Rev. Cyt. 1987, 109, 89–112. [Google Scholar]
- Mukherjee, A.; Koli, S.; Reddy, K. Regulatory non-coding transcripts in spermatogenesis: Shedding light on ‘dark matter’. Andrology. 2014, 2, 360–369. [Google Scholar] [CrossRef]
- Lai, E.C. Micro RNAs are complementary to 3′ UTR sequence motifs that mediate negative post-transcriptional regulation. Nat. Genet. 2002, 30, 363–364. [Google Scholar] [CrossRef]
- Gendron, C.M.; Pletcher, S.D. Micro RNA s mir-184 and let-7 alter Drosophila metabolism and longevity. AGING C. 2017, 16, 1434–1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozano, J.; Montañez, R.; Belles, X. MiR-2 family regulates insect metamorphosis by controlling the juvenile hormone signaling pathway. Proc. Natl. Acad. Sci. USA 2015, 112, 3740–3745. [Google Scholar] [CrossRef] [PubMed]
- Perdomo, H.D.; Hussain, M.; Parry, R.; Etebari, K.; Hedges, L.M.; Zhang, G.; Schulz, B.L.; Asgari, S. Human blood microRNA hsa-miR-21-5p induces vitellogenin in the mosquito Aedes aegypti. Commun. Biol. 2021, 4, 856. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.-J.; Liu, Y.-J.; Zhu, F.; Cai, L.-M.; Liu, X.-M.; Tian, Z.-Q.; Cheng, J.; Li, Z.; Liu, X.-X. MicroRNA-277 regulates dopa decarboxylase to control larval-pupal and pupal-adult metamorphosis of Helicoverpa armigera. Insect Biochem. Mol. Biol. 2020, 122, 103391. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Xiao, B.; Liu, Z.; Li, N.; Zhu, E.-D.; Li, B.-S.; Xie, Q.-H.; Zhuang, Y.; Zou, Q.-M.; Mao, X.-H. Identification of MyD88 as a novel target of miR-155, involved in negative regulation of Helicobacter pylori-induced inflammation. FEBS Lett. 2010, 584, 1481–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhao, B.; Roy, S.; Saha, T.T.; Kokoza, V.A.; Li, M.; Raikhel, A.S. microRNA-309 targets the Homeobox gene SIX4 and controls ovarian development in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2016, 113, E4828–E4836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambros, V.; Bartel, B.; Bartel, D.P.; Burge, C.B.; Carrington, J.C.; Chen, X.; Dreyfuss, G.; Eddy, S.R.; Griffiths-Jones, S.; Marshall, M. A uniform system for microRNA annotation. RNA 2003, 9, 277–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brennecke, J.; Hipfner, D.R.; Stark, A.; Russell, R.B.; Cohen, S.M. Bantam encodes a developmentally regulated microRNA that controls cell proliferation and regulates the proapoptotic gene hid in Drosophila. Cell 2003, 113, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.; Liu, Y.; Sun, M.; Niu, M.; Yuan, Q.; Hai, Y.; Guo, Y.; Chen, Z.; Hou, J.; Liu, Y. MicroRNAs and DNA methylation as epigenetic regulators of mitosis, meiosis and spermiogenesis. REPR 2015, 150, R25–R34. [Google Scholar] [CrossRef] [Green Version]
- Pisano, F.; Altomare, C.; Cervio, E.; Barile, L.; Rocchetti, M.; Ciuffreda, M.C.; Malpasso, G.; Copes, F.; Mura, M.; Danieli, P. Combination of miRNA-499 and miRNA-133 Exerts a Synergic Effect on Cardiac Differentiation. Stem Cells 2015, 33, 1187–1199. [Google Scholar] [CrossRef]
- Zheng, Z.; Guan, M.; Jia, Y.; Wang, D.; Pang, R.; Lv, F.; Xiao, Z.; Wang, L.; Zhang, H.; Xue, Y. The coordinated roles of miR-26a and miR-30c in regulating TGFβ1-induced epithelial-to-mesenchymal transition in diabetic nephropathy. Sci. Rep. 2016, 6, 37492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Wu, H.; Wen, Y.; Liu, Y.; Zhou, T.; Ni, B.; Lin, Y.; Dong, J.; Zhou, Z.; Hu, Z. Identification of seven genes essential for male fertility through a genome-wide association study of non-obstructive azoospermia and RNA interference-mediated large-scale functional screening in Drosophila. Hum. Mol. Genet. 2015, 24, 1493–1503. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Zhang, Y.; Yang, Q.; Liu, B.; Wu, J.; Zhang, Y.; Yang, C.; Jiang, Y. miR-497 and miR-34a retard lung cancer growth by co-inhibiting cyclin E1 (CCNE1). Oncotarget 2015, 6, 13149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tariq, K.; Peng, W.; Saccone, G.; Zhang, H. Identification, characterization and target gene analysis of testicular microRNAs in the oriental fruit fly Bactrocera dorsalis. Insect Mol. Biol. 2016, 25, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Hafer, N.; Agunwamba, B.; Schedl, P. The CPEB protein Orb2 has multiple functions during spermatogenesis in Drosophila melanogaster. PLoS Genet. 2012, 8, e1003079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Lucas, K.J.; Roy, S.; Ha, J.; Raikhel, A.S. Mosquito-specific microRNA-1174 targets serine hydroxymethyltransferase to control key functions in the gut. Proc. Natl. Acad. Sci. USA 2014, 111, 14460–14465. [Google Scholar] [CrossRef] [Green Version]
- Krüger, J.; Rehmsmeier, M. RNAhybrid: microRNA target prediction easy, fast and flexible. Nucleic Acids Res. 2006, 34, W451–W454. [Google Scholar] [CrossRef]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, M.; Zhang, H. RNA interference of four genes in adult Bactrocera dorsalis by feeding their dsRNAs. PLoS ONE. 2011, 6, e17788. [Google Scholar] [CrossRef]
- Den Boer, S.P.; Baer, B.; Boomsma, J.J. Seminal fluid mediates ejaculate competition in social insects. Science. 2010, 327, 1506–1509. [Google Scholar] [CrossRef] [Green Version]
- Den Boer, S.P.; Boomsma, J.J.; Baer, B. Seminal fluid enhances sperm viability in the leafcutter ant Atta colombica. Behav. Ecol. Sociobiol. 2008, 62, 1843–1849. [Google Scholar] [CrossRef] [Green Version]
- Koyama, J.; Kakinohana, H.; Miyatake, T. Eradication of the melon fly, Bactrocera cucurbitae, in Japan: Importance of behavior, ecology, genetics, and evolution. Annu. Rev. Entomol. 2004, 49, 331–349. [Google Scholar] [CrossRef]
- Oliva, C.F.; Jacquet, M.; Gilles, J.; Lempérière, G.; Maquart, P.-O.; Quilici, S.; Schooneman, F.; Vreysen, M.J.; Boyer, S. The sterile insect technique for controlling populations of Aedes albopictus (Diptera: Culicidae) on Reunion Island: Mating vigour of sterilized males. PLoS ONE 2012, 7, e49414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, G. Genetic sexing strains in Mediterranean fruit fly, an example for other species amenable to large-scale rearing for the sterile insect technique. In The Sterile Insect Technique; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 427–451. [Google Scholar]
- Robinson, A.S. Genetic basis of the sterile insect technique. In Sterile Insect Technique; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 95–114. [Google Scholar]
- Burt, A. Site-specific selfish genes as tools for the control and genetic engineering of natural populations. Proc. R. Soc. Lond. B 2003, 270, 921–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef]
- Friedman, R.C.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Tyagi, S.; Schedl, P. Spermatid cyst polarization in Drosophila depends upon apkc and the CPEB family translational regulator orb2. PLoS Genet. 2014, 10, e1004380. [Google Scholar] [CrossRef]
- Esau, C.C. Inhibition of microRNA with antisense oligonucleotides. Methods 2008, 44, 55–60. [Google Scholar] [CrossRef]
- Meister, G.; Landthaler, M.; Patkaniowska, A.; Dorsett, Y.; Teng, G.; Tuschl, T. Human Argonaute2 mediates RNA cleavage targeted by miRNAs and siRNAs. Mol. Cell 2004, 15, 185–197. [Google Scholar] [CrossRef]
- Krützfeldt, J.; Kuwajima, S.; Braich, R.; Rajeev, K.G.; Pena, J.; Tuschl, T.; Manoharan, M.; Stoffel, M. Specificity, duplex degradation and subcellular localization of antagomirs. Nucleic Acids Res. 2007, 35, 2885–2892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, B.; Dalby, A.B.; Karpilow, J.; Khvorova, A.; Leake, D.; Vermeulen, A. Specificity and functionality of microRNA inhibitors. Silence 2010, 1, 10. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sohail, S.; Tariq, K.; Sajid, M.; Ali, M.W.; Peng, W.; Zhang, H. miR-125-3p and miR-276b-3p Regulate the Spermatogenesis of Bactrocera dorsalis by Targeting the orb2 Gene. Genes 2022, 13, 1861. https://doi.org/10.3390/genes13101861
Sohail S, Tariq K, Sajid M, Ali MW, Peng W, Zhang H. miR-125-3p and miR-276b-3p Regulate the Spermatogenesis of Bactrocera dorsalis by Targeting the orb2 Gene. Genes. 2022; 13(10):1861. https://doi.org/10.3390/genes13101861
Chicago/Turabian StyleSohail, Summar, Kaleem Tariq, Muhammad Sajid, Muhammad Waqar Ali, Wei Peng, and Hongyu Zhang. 2022. "miR-125-3p and miR-276b-3p Regulate the Spermatogenesis of Bactrocera dorsalis by Targeting the orb2 Gene" Genes 13, no. 10: 1861. https://doi.org/10.3390/genes13101861