
DNA Barcoding and Species Classification of Morchella

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. DNA Extraction, PCR Amplication and Sequencing
2.3. DNA Molecular Barcoding
2.4. Phylogenetic Relationship Analysis
2.5. Diversification Time Estimates
3. Results
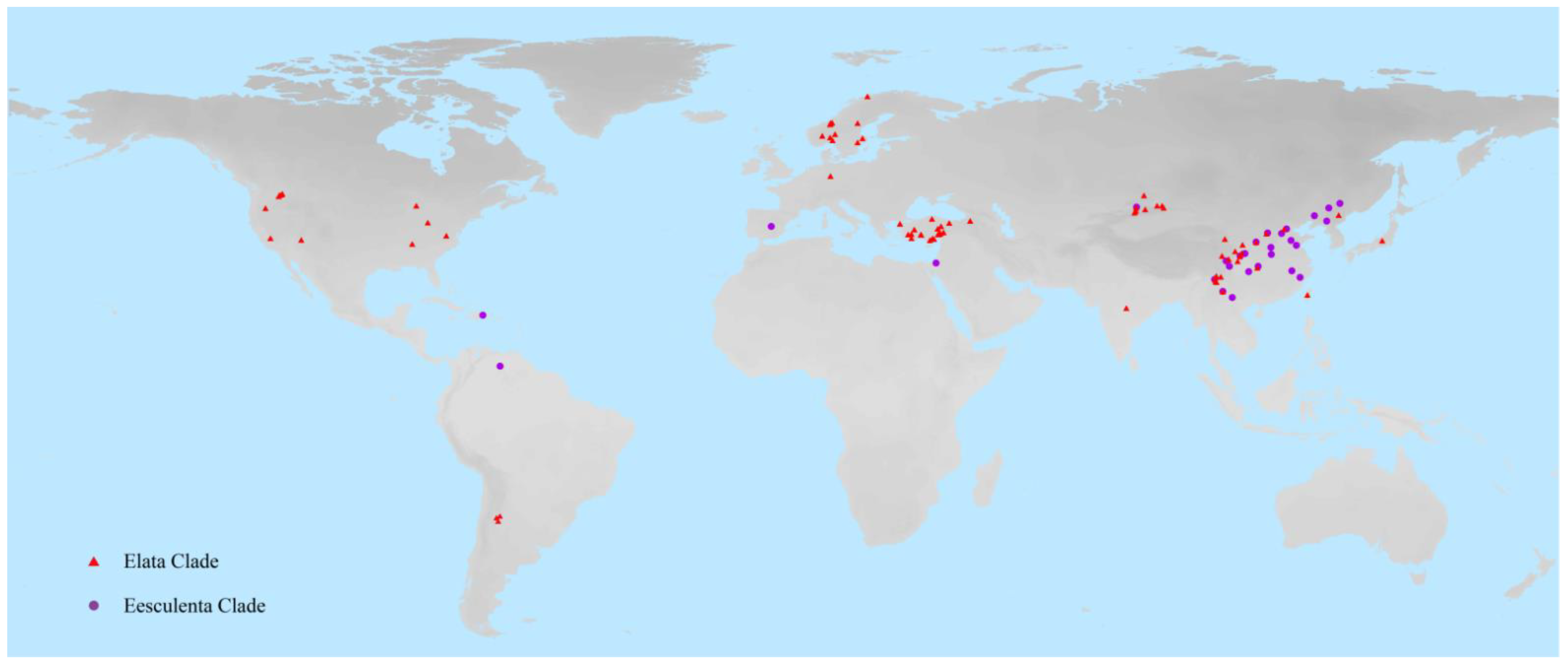
3.1. Sampling and DNA Molecular Sequences
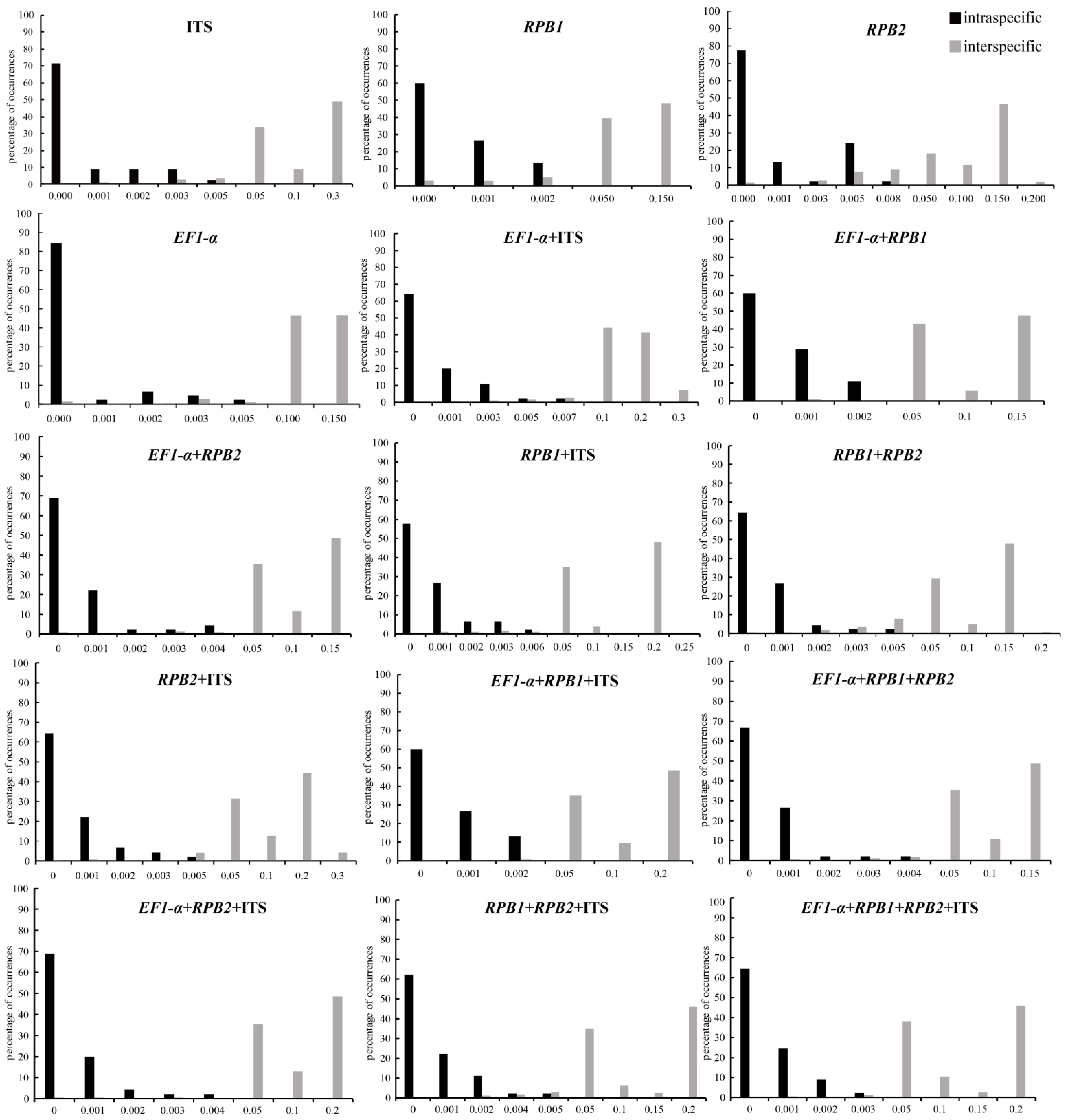
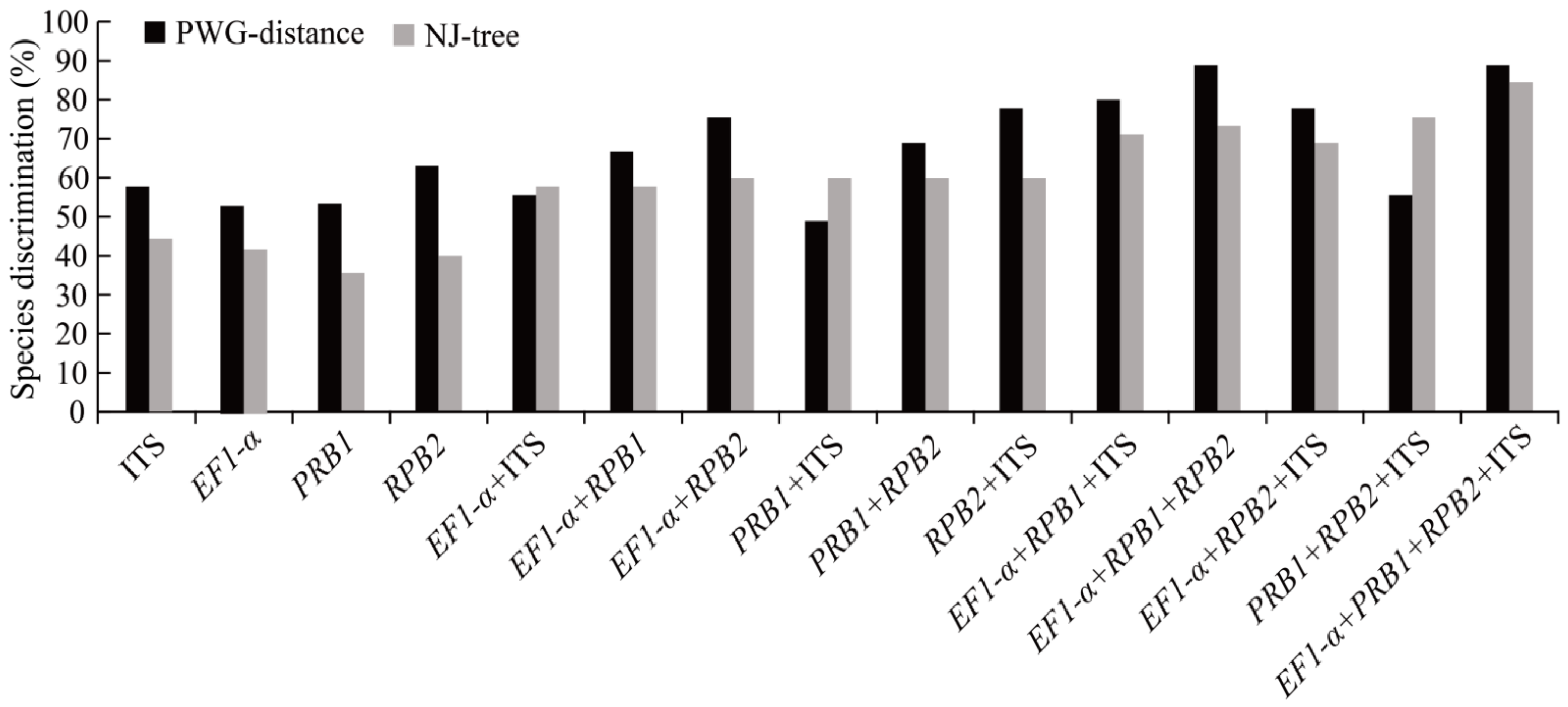
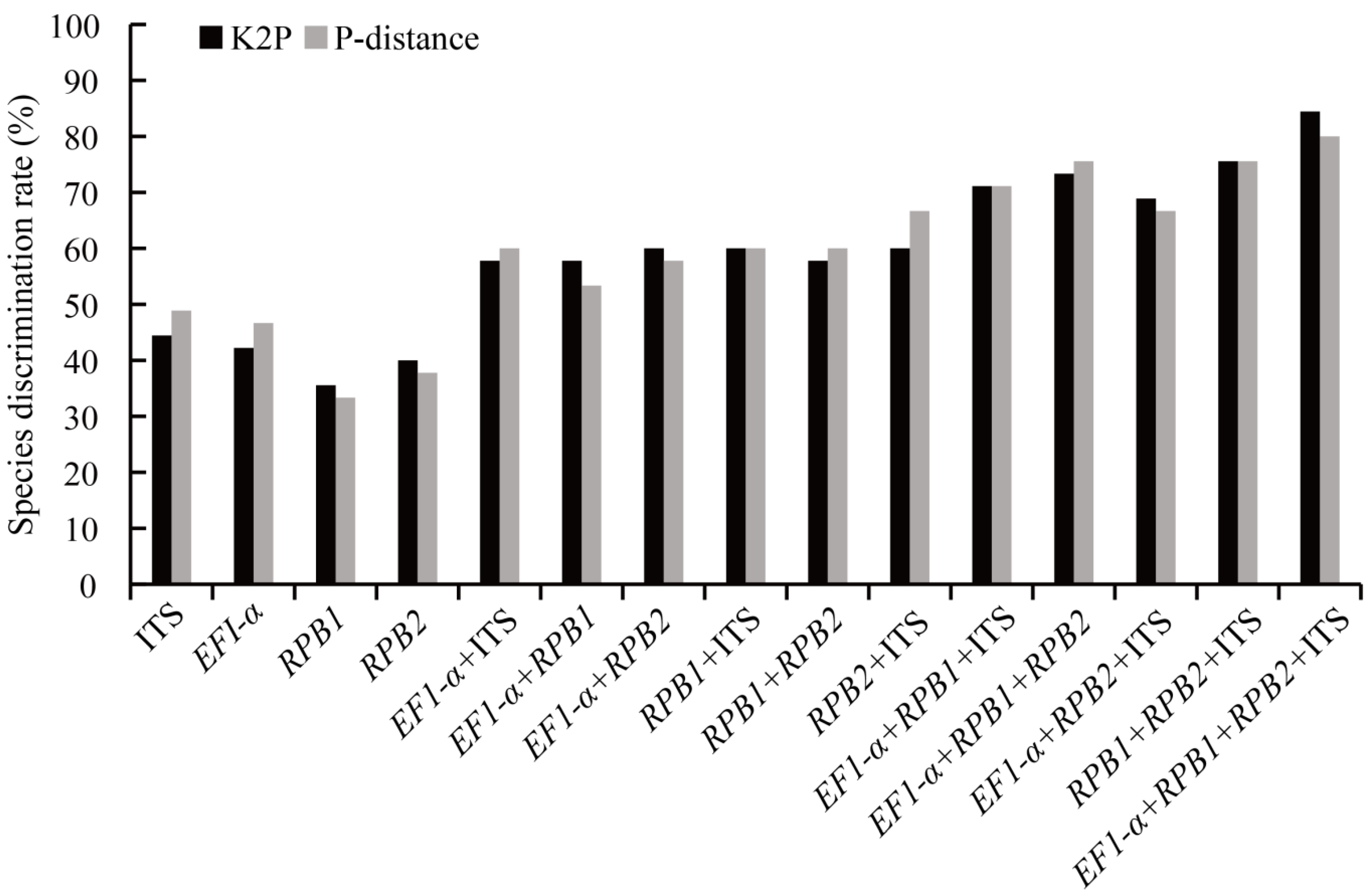
3.2. DNA Barcodes
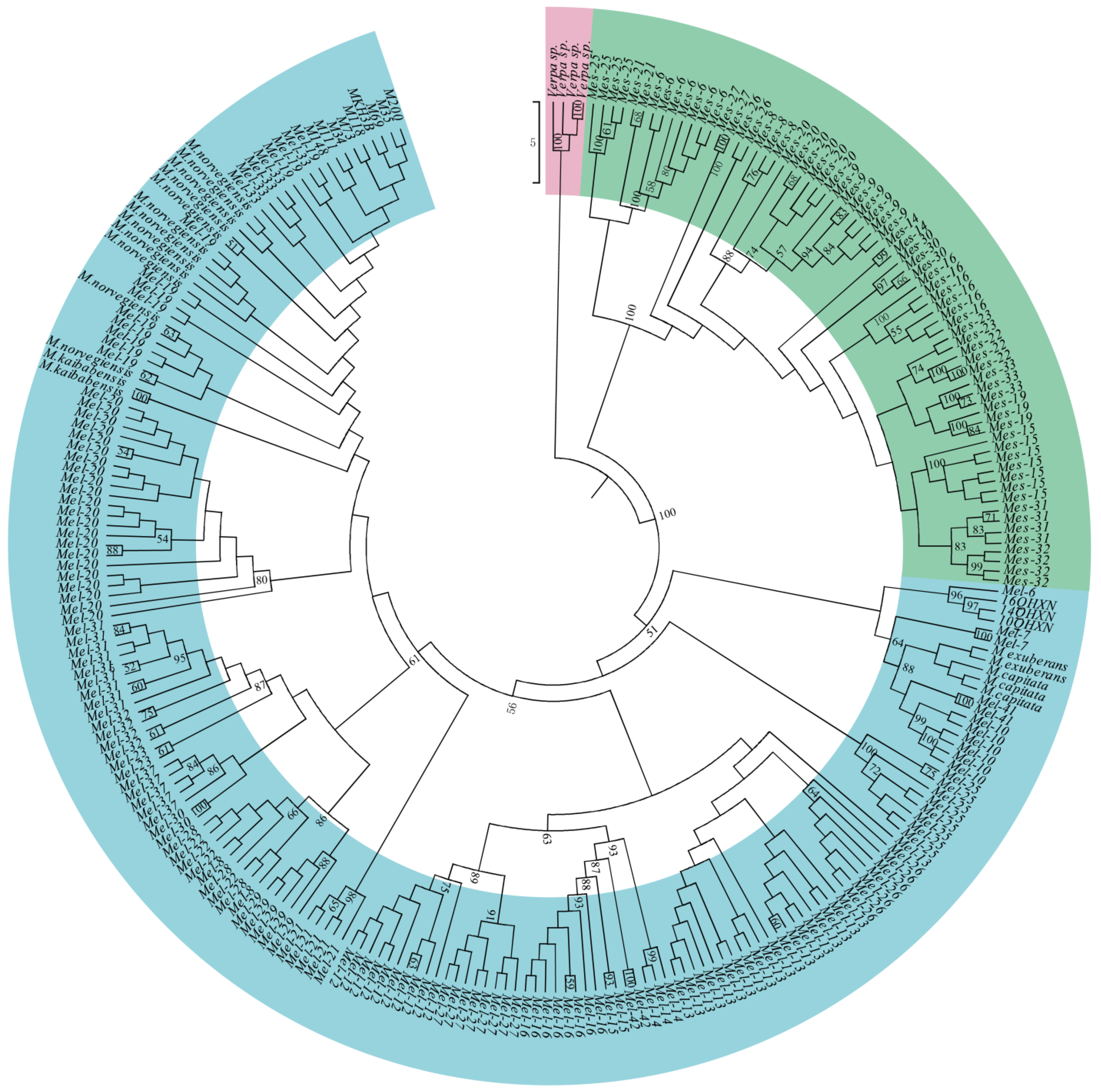
3.3. Phylogenetic Relationship
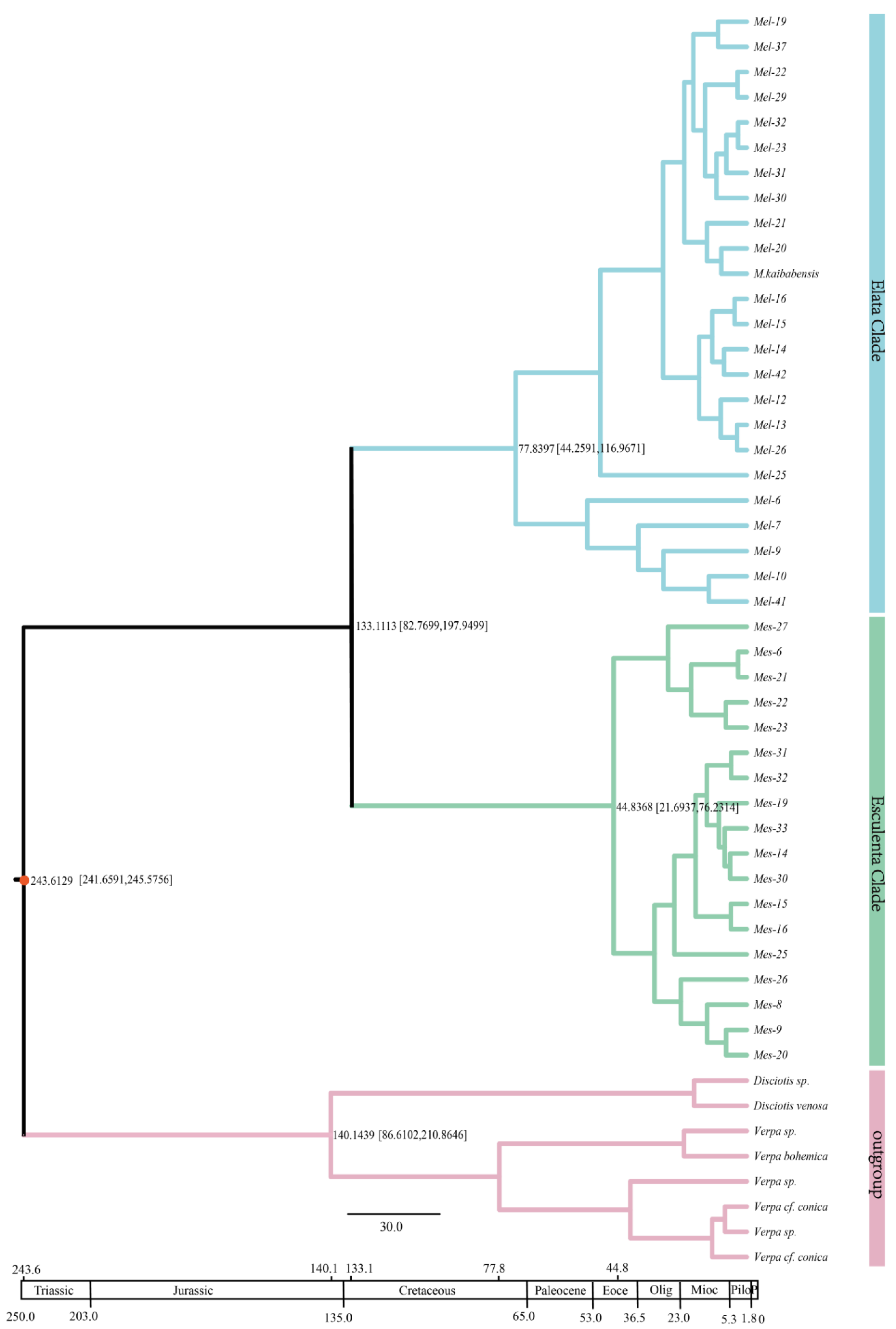
3.4. Divergence Time of Morchella
4. Discussion
4.1. Evaluation of Barcoding Efficiency of Morchella
4.2. Phylogenetic Relationships of Morchella
4.3. Divergence Time of Morchella
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hibbett, D.; Binder, M.; Bischoff, J.F.; Blackwell, M.; Cannon, P.F.; Eriksson, O.E.; Huhndorf, S.; James, T.; Kirk, P.M.; Lucking, R.; et al. A higher-level phylogenetic classification of the Fungi. Mycol. Res. 2007, 111, 509–547. [Google Scholar] [CrossRef]
- Tietel, Z.; Masaphy, S. True morels (Morchella)-nutritional and phytochemical composition, health benefits and flavor: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1888–1901. [Google Scholar] [CrossRef]
- Du, X.H.; Zhao, Q.; O’Donnell, K.; Rooney, A.P.; Yang, Z.L. Multigene molecular phylogenetics reveals true morels (Morchella) are especially species-rich in China. Fungal Genet. Biol. 2012, 49, 455–469. [Google Scholar] [CrossRef]
- Bunyard, B.A.; Nicholson, M.S.; Royse, D.J. Phylogenetic resolution of Morchella, Verpa, and Disciotis [Pezizales: Morchellaceae] based on restriction enzyme analysis of the 28S ribosomal RNA gene. Exp. Mycol. 1995, 19, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Volk, T.J.; Leonard, T.J. Experimental studies on the morel. I. Heterokaryon formation between monoascosporous strains of Morchella. Mycologia 1989, 81, 523–531. [Google Scholar] [CrossRef]
- Wipf, D.; Munch, J.C.; Botton, B.; Buscot, F. DNA polymorphism in morels: Complete sequences of the internal transcribed spacer of genes coding for rRNA in Morchella esculenta (yellow morels) and Morchella conica (black morels). Appl. Environ. Microbiol. 1996, 62, 3541–3543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzmán, G.; Tapia, F. The known morels in Mexico, a description of a new blushing species, Morchella rufobrunnea, and new data on M. guatemalensis. Mycologia 1998, 90, 705–714. [Google Scholar] [CrossRef]
- Du, X.H.; Zhao, Q.; Yang, Z.L. Diversity, evolutionary history and cultivation of morels: A review. Mycosystema 2014, 33, 183–197. [Google Scholar] [CrossRef]
- Kanwal, H.K.; Acharya, K.; Ramesh, G.; Reddy, M.S. Molecular characterization of Morchella species from the western Himalayan region of India. Curr. Biol. 2011, 62, 1245–1252. [Google Scholar] [CrossRef]
- Clowez, P. Les morilles. Une nouvelle approche mondiale du genre Morchella. Bull. Soc. Mycol. Fr. 2012, 126, 199–376. [Google Scholar]
- Kuo, M.; Dewsburry, D.R.; O’Donnell, K.; Carter, M.C.; Rehner, S.A.; Moore, J.D.; Moncalvo, J.M.; Canfield, S.A.; Methven, A.S.; Volk, T.J. Taxonomic revision of true morels (Morchella) in Canada and the United States. Mycologia 2012, 104, 1159–1177. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, P.E.; Karunarathna, S.C.; Li, Q.H.; Gui, H.; Yang, X.Q.; Yang, X.F.; He, J.; Ye, L.; Guo, J.Y.; Li, H.L.; et al. Prized edible Asian mushrooms: Ecology, conservation and sustainability. Fungal Divers. 2012, 56, 31–47. [Google Scholar] [CrossRef]
- Richard, F.; Bellanger, J.M.; Clowez, P.; Hansen, K.; O’Donnell, K.; Urban, A.; Sauve, M.; Courtecuisse, M.; Moreau, P.A. True morels (Morchella, Pezizales) of Europe and North America: Evolutionary relationships inferred from multilocus data and a unified taxonomy. Mycologia 2015, 107, 359–382. [Google Scholar] [CrossRef] [PubMed]
- Nedunoori, A.; Turanov, S.V.; Kartavtsev, Y.P. Fish product mislabeling identified in the Russian far east using DNA barcoding. Gene Reports 2017, 8, 144–149. [Google Scholar] [CrossRef]
- Waugh, J. DNA barcoding in animal species: Progress, potential and pitfalls. Bioessays 2007, 29, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, M.A.; Baraloto, C.; Engel, J.; Mori, S.A.; Pétronelli, P.; Riéra, B.; Roger, A.; Thébaud, C.; Chave, J. Identification of Amazonian trees with DNA barcodes. PLoS ONE 2009, 4, e7483. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Cigelnik, E.; Weber, N.S.; Trappe, J.M. Phylogenetic relationships among ascomycetous trufflfles and the true and false morels inferred from 18S and 28S ribosomal DNA sequence analysis. Mycologia 1997, 89, 48–65. [Google Scholar] [CrossRef]
- O’Donnell, K.; Rooney, A.P.; Mills, G.L.; Kuo, M.; Weber, N.S.; Rehner, S.A. Phylogeny and historical biogeography of true morels (Morchella) reveals an early Cretaceous origin and high continental endemism and provincialism in the Holarctic. Fungal Genet. Biol. 2011, 48, 252–265. [Google Scholar] [CrossRef]
- Zeng, N.K.; Tang, L.P.; Li, Y.C.; Tolgor, B.; Zhu, X.T.; Zhao, Q.; Yang, Z.L. The genus Phylloporus (Boletaceae, Boletales) from China: Morphological and multilocus DNA sequence analyses. Fungal Divers. 2013, 58, 73–101. [Google Scholar] [CrossRef]
- Taşkın, H.; Büyükalaca, S.; Doğan, H.H.; Rehner, A.A.; O’Donnell, K. A multigene molecular phylogenetic assessment of true morels (Morchella) in Turkey. Fungal Genet. Biol. 2010, 47, 672–682. [Google Scholar] [CrossRef]
- Taşkın, H.; Büyükalaca, S.; Hansen, K.; O’Donnell, K. Multilocus phylogenetic analysis of true morels (Morchella) reveals high levels of endemics in Turkey relative to other regions of Europe. Mycologia 2017, 104, 446–461. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Kasuga, T.; Geiser, D.M.; Hibbett, D.S.; Fisher, M.C. Phylogenetic species recognition and species concepts in fungi. Fungal Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauserud, H.; Schumacher, T. Outcrossing or inbreeding: DNA markers provide evidence for type of reproductive mode in Phellinus nigrolimitatus (Basidiomycota). Mycol. Res. 2001, 105, 676–683. [Google Scholar] [CrossRef] [Green Version]
- Du, X.H.; Zhao, Q.; Yang, Z.L.; Hansen, K.; Taskin, H.; Büyükalaca, S.; Dewsbury, D.; Moncalvo, J.M.; Douhan, G.W.; Robert, V.A.; et al. How well do ITS rDNA sequences differentiate species of true morels (Morchella)? Mycologia 2012, 104, 1351–1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CBOL Plant Working Group. A DNA barcode for land plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12794–12797. [Google Scholar] [CrossRef] [Green Version]
- Lahaye, R.; Van der Bank, M.; Bogarin, D.; Warner, J.; Pupulin, F.; Gigot, G.; Maurin, O.; Duthoit, S.; Barraclough, T.G.; Savolainen, V. DNA barcoding the floras of biodiversity hotspots. Proc. Natl. Acad. Sci. USA 2008, 105, 2923–2928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollingsworth, M.L.; Clark, A.A.; Forrest, L.L.; Richardson, J.; Pennington, R.T.; Long, D.G.; Cowan, R.; Chase, M.W.; Gaudeul, M.; Hollingsworth, P.M. Selecting barcoding loci for plants: Evaluation of seven candidate loci with species-level sampling in three divergent groups of land plants. Mol. Ecol. Resour. 2009, 9, 439–457. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BioMed. Cen. Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berbee, M.L.; Taylor, J.W. Dating the molecular clock in Fungi-how close are we? Fung. Biol. Rev. 2010, 24, 1–15. [Google Scholar] [CrossRef]
- Drummond, A.J.; Ho, S.Y.; Rambaut, A. Relaxed phylogenetics and dating with confidence. PLoS Biol. 2006, 4, e88. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J. Tracer: MCMC Trace Analysis Tool Version v1.5.0. 2009. Available online: http://tree.bio.ed.ac.uk/software/tracer (accessed on 1 August 2017).
- Rambaut, A.; Suchard, M.A.; Xie, D.; Drummond, A.J. TreeAnnotator v1.8.0. 2014. Available online: http://beast.bio.ed.ac.uk/TreeAnnotator (accessed on 11 July 2015).
- Rambaut, A. FigTree, Version 1.3.1. 2009. Available online: http://tree.bio.ed.ac.uk/software/figtree (accessed on 17 May 2013).
- Meyer, C.P.; Paulay, G. DNA barcoding: Error rates based on comprehensive sampling. PLoS Biol. 2005, 3, 2229–2238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrželová, I.; Sochor, M. How useful is the current species recognition concept for the determination of true morels? Insights from the Czech Republic. MycoKeys 2019, 52, 17–43. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Ward, T.J.; Geiser, D.M.; Kistler, H.C.; Aoki, T. Genealogical concordance between the mating type locus and seven other nuclear genes supports formal recognition of nine phylogenetically distinct species within the Fusarium graminearum clade. Fungal Genet. Biol. 2004, 41, 600–623. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.H.; Wingfield, B.D.; Wingfield, M.J.; Steenkamp, E.T. Structure and evolution of the Fusarium mating type locus: New insights from the Gibberella fujikuroi complex. Fungal Genet. Biol. 2011, 48, 731–740. [Google Scholar] [CrossRef] [Green Version]
- Chai, H.; Chen, W.; Zhang, X.; Su, K.; Zhao, Y. Structural variation and phylogenetic analysis of the mating-type locus in the genus Morchella. Mycologia 2019, 111, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Du, X.H.; Zhao, Q.; Xia, E.H.; Gao, L.Z.; Richard, F.; Yang, Z.L. Mixed-reproductive strategies, competitive mating-type distribution and life cycle of fourteen black morel species. Sci. Rep. 2017, 7, 1493. [Google Scholar] [CrossRef] [PubMed]
- Loizides, M.; Bellanger, J.M.; Clowez, P.; Richard, F.; Moreau, P.A. Combined phylogenetic and morphological studies of true morels (Pezizales, Ascomycota) in Cyprus reveal significant diversity, including Morchella arbutiphila and M. disparilis spp. nov. Mycol. Progress 2016, 15, 39. [Google Scholar] [CrossRef]
- Brower, A.V. A new mimetic species of Heliconius (Lepidoptera:Nymphalidae), from southeastern Colombia, revealed by cladistic analysis of mitochondrial DNA sequence. Zool. J. Linn. Soc. 1996, 116, 317–332. [Google Scholar] [CrossRef]
- Phanpadith, P.; Yu, Z.; Li, T. High diversity of Morchella and a novel lineage of the Esculenta clade from the north Qinling Mountains revealed by GCPSR-based study. Sci. Rep. 2019, 9, 19856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baroni, T.J.; Beug, M.W.; Cantrell, S.A.; Clements, T.A.; Iturriaga, T.; Læssøe, T.; Holgado Rojas, M.E.; Aguilar, F.M.; Quispe, M.O.; Lodge, D.J.; et al. Four new species of Morchella from the Americas. Mycologia 2018, 110, 1205–1221. [Google Scholar] [CrossRef]
- Du, X.H.; Zhao, Q.; Yang, Z.L. A review on research advances, issues, and perspectives of morels. Mycology 2015, 6, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Du, X.H.; Wu, D.M.; He, G.Q.; Wei, W.; Xu, N.; Li, T.L. Six new species and two new records of Morchella in China using phylogenetic and morphological analyses. Mycologia 2019, 111, 857–870. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes:evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.X.; Fu, C.X.; Comes, H.P. Plant molecular phylogeography in China and adjacent regions: Tracing the genetic imprints of quaternary climate and environmental change in the world’s most diverse temperate flora. Mol. Phylogenet. Evol. 2011, 59, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Du, X.H.; Zhao, Q.; Xu, J.; Yang, Z.L. High inbreeding, limited recombination and divergent evolutionary patterns between two sympatric morel species in China. Sci. Rep. 2016, 6, 22434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matheny, P.B. Improving phylogenetic inference of mushrooms with RPB1 and RPB2 nucleotide sequences (Inocybe; Agaricales). Mol. Phylogenet. Evol. 2005, 35, 1–20. [Google Scholar] [CrossRef]
- Sanmartín, I.; Enghoff, H.; Ronquist, F. Patterns of animal dispersal, vicariance and diversifification in the Holarctic. Biol. J. Linn. Soc. 2001, 73, 345–390. [Google Scholar] [CrossRef]
- Bai, W.N.; Wang, W.T.; Zhang, D.Y. Phylogeographic breaks within Asian butternuts indicate the existence of a phytogeographic divide in East Asia. New Phytol. 2016, 209, 1757–1772. [Google Scholar] [CrossRef] [Green Version]
- Dansgaard, W.; Johnsen, S.J.; Clausen, H.B.; Dahl-Jensen, D.; Gundestrup, N.S.; Hammer, C.U.; Hvidberg, C.S.; Steffensen, J.P.; Sveinbjörnsdottir, A.E.; Jouzel, J.; et al. Evidence for general instability of past climate from a 250-kyr ice-core record. Nature 1993, 364, 218–220. [Google Scholar] [CrossRef]
- Van Andel, T.H.; Tzedakis, P.C. Palaeolithic landscapes of Europe and environs, 150,000-25,000 years ago: An overview. Quat. Sci. Rev. 1996, 15, 481–500. [Google Scholar] [CrossRef]
- Hewitt, G.M. The genetic legacy of the quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Harrison, S.P.; Yu, G.; Takahara, H.; Prentice, I.C. Palaeovegetation (communications arising): Diversity of temperate plants in East Asia. Nature 2001, 413, 129–130. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.L. Diversity and biogeography of higher fungi in China. In Evolutionary Genetics of Fungi; Xu, J.P., Ed.; Horizon Bioscience: Norfolk, VA, USA, 2005; pp. 35–62. [Google Scholar]
- Soltis, D.E.; Morris, A.B.; McLachlan, J.S.; Manos, P.S.; Soltis, P.S. Comparative phylogeography of unglaciated eastern North America. Mol. Ecol. 2006, 15, 4261–4293. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DNA Region | Primer Pairs | Primer Sequences (5′-3′) | Thermal Cycling Conditions | References |
|---|---|---|---|---|
| EF1-α | EF1-α-F | ACTCCTAAGTACTATGTCACCGTCATT | 94 °C 2 min, 35 cycles (94 °C 1 min, 55 °C 40 s, 72 °C 1 min), 72 °C 10 min | [24] |
| EF1-α-R | TGGAGAGGAAGACGGAGAGGCTT | |||
| RPB1 | RPB1-F | TATATCACGTCGGTATGTATCCACTC | 94 °C 2 min, 35 cycles (94 °C 1 min, 55 °C 40 s, 72 °C 1 min), 72 °C 10 min | [25] |
| RPB1-R | ATTTGCTCGGATGATCTCAG | |||
| RPB2 | RPB2-F | TAGGTAGGTCCCAAGAACACC | 94 °C 2 min, 35 cycles (94 °C 1 min, 55 °C 40 s, 72 °C 1 min), 72 °C 10 min | [25] |
| RPB2-R | GATACCATGGCGAACATTCTG | |||
| ITS | ITS-1 | TCCGTAGGTGAACCTGCGG | 94 °C 2 min, 35 cycles (94 °C 1 min, 55 °C 40 s, 72 °C 1 min), 72 °C 10 min | [26] |
| ITS-4 | TCCTCCGCTTATTGATATGC |
| Gene | Length (bp) | No. SNPs | % SNP | No. inDels | % No. Variable Sites | Intraspecific Distance (Mean) | Interspecific Distance (Mean) | NJ Rate (%) P-Distance | NJ Rate (%) K2P | PWG-Distance Rate (%) K2P |
|---|---|---|---|---|---|---|---|---|---|---|
| RPB1 | 420 | 94 | 22.38 | 2 | 22.85 | 0.00056 | 0.0673 | 33.33 | 35.56 | 53.33 |
| RPB2 | 391 | 86 | 21.99 | 0 | 21.99 | 0.00057 | 0.0026 | 37.78 | 40.00 | 63.04 |
| ITS | 452 | 125 | 27.65 | 78 | 44.91 | 0.00062 | 0.1347 | 48.89 | 44.44 | 57.78 |
| EF1-α | 312 | 76 | 24.36 | 18 | 30.13 | 0.00040 | 0.077 | 46.67 | 42.22 | 53.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sa, W.; Qiao, J.; Gao, Q.; Li, Z.; Shang, Q. DNA Barcoding and Species Classification of Morchella. Genes 2022, 13, 1806. https://doi.org/10.3390/genes13101806
Sa W, Qiao J, Gao Q, Li Z, Shang Q. DNA Barcoding and Species Classification of Morchella. Genes. 2022; 13(10):1806. https://doi.org/10.3390/genes13101806
Chicago/Turabian StyleSa, Wei, Jinxia Qiao, Qiyuan Gao, Zhonghu Li, and Qianhan Shang. 2022. "DNA Barcoding and Species Classification of Morchella" Genes 13, no. 10: 1806. https://doi.org/10.3390/genes13101806