
Changes in Expression of Specific mRNA Transcripts after Single- or Re-Irradiation in Mouse Testes

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
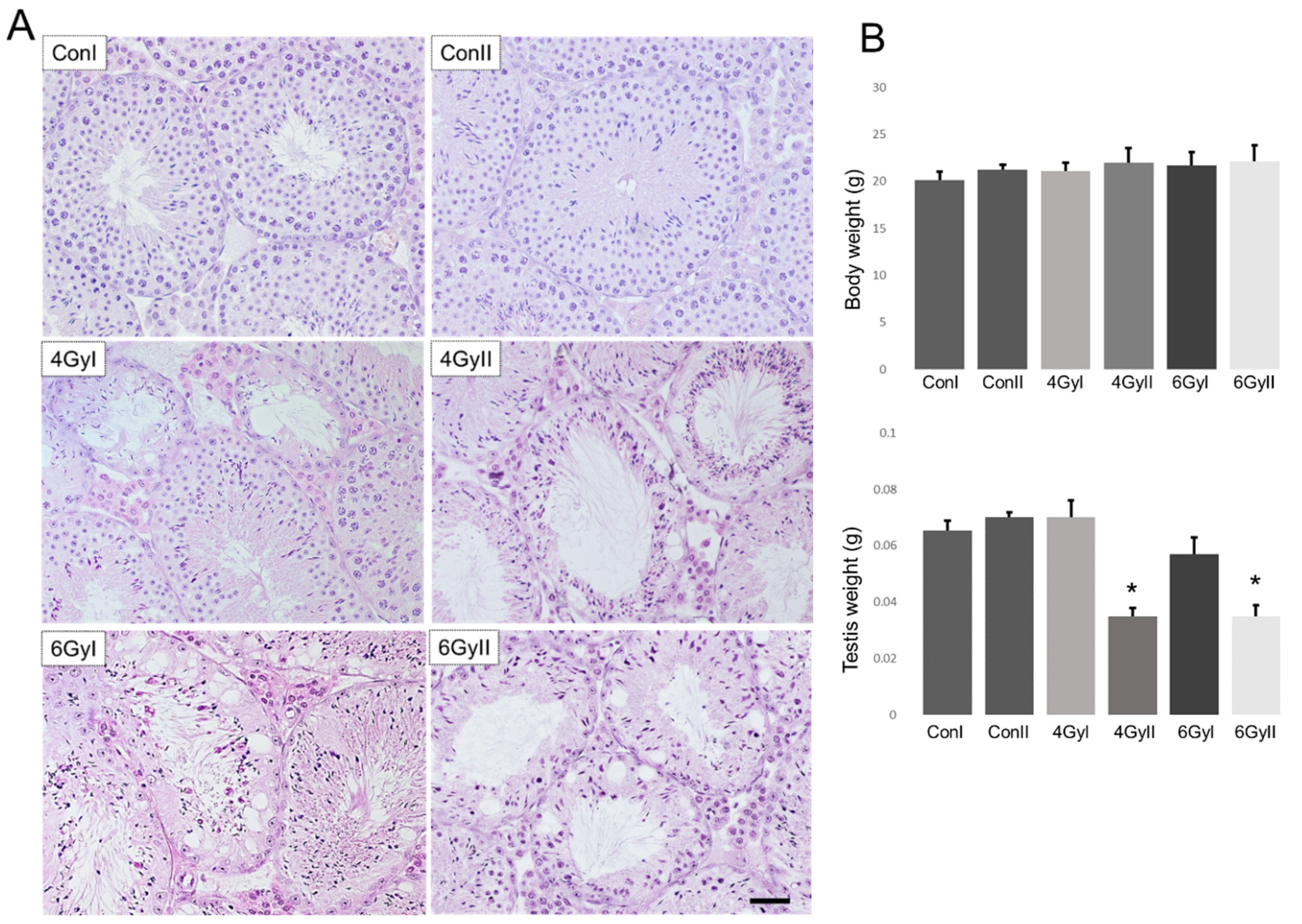
2.3. Histological Examination of the Testes
2.4. Detection of Serum Anti-Germ Cell Antibodies
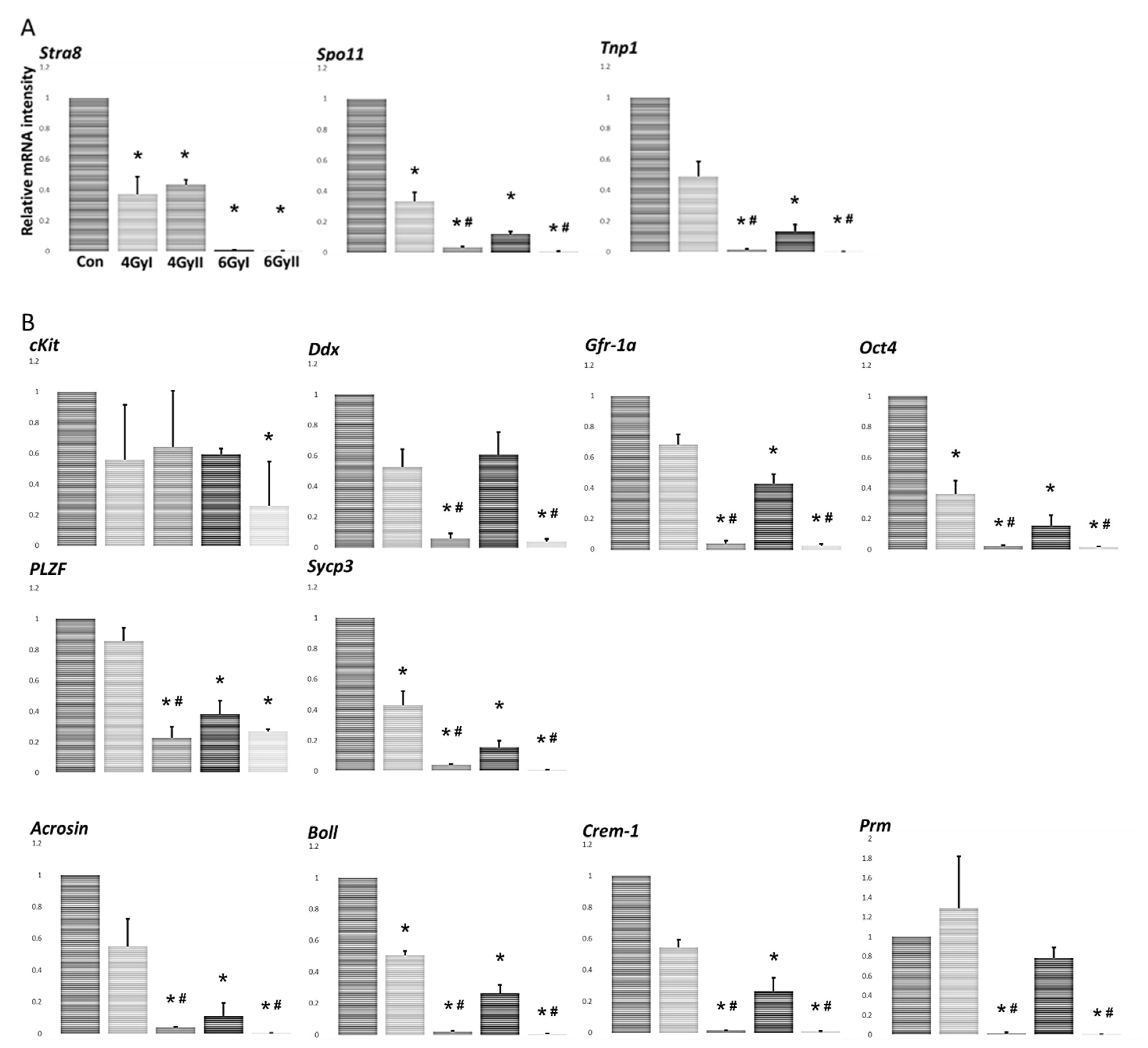
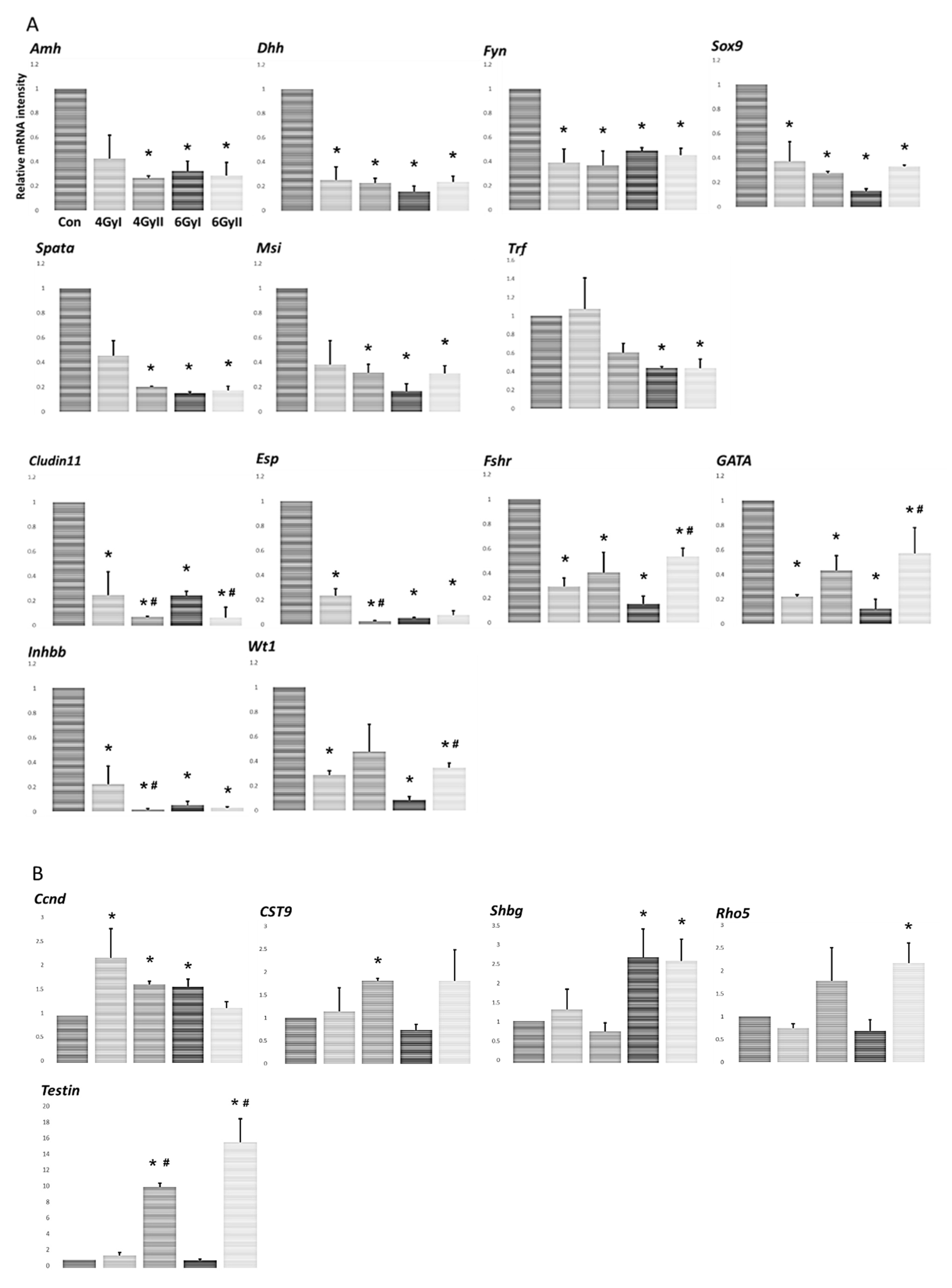
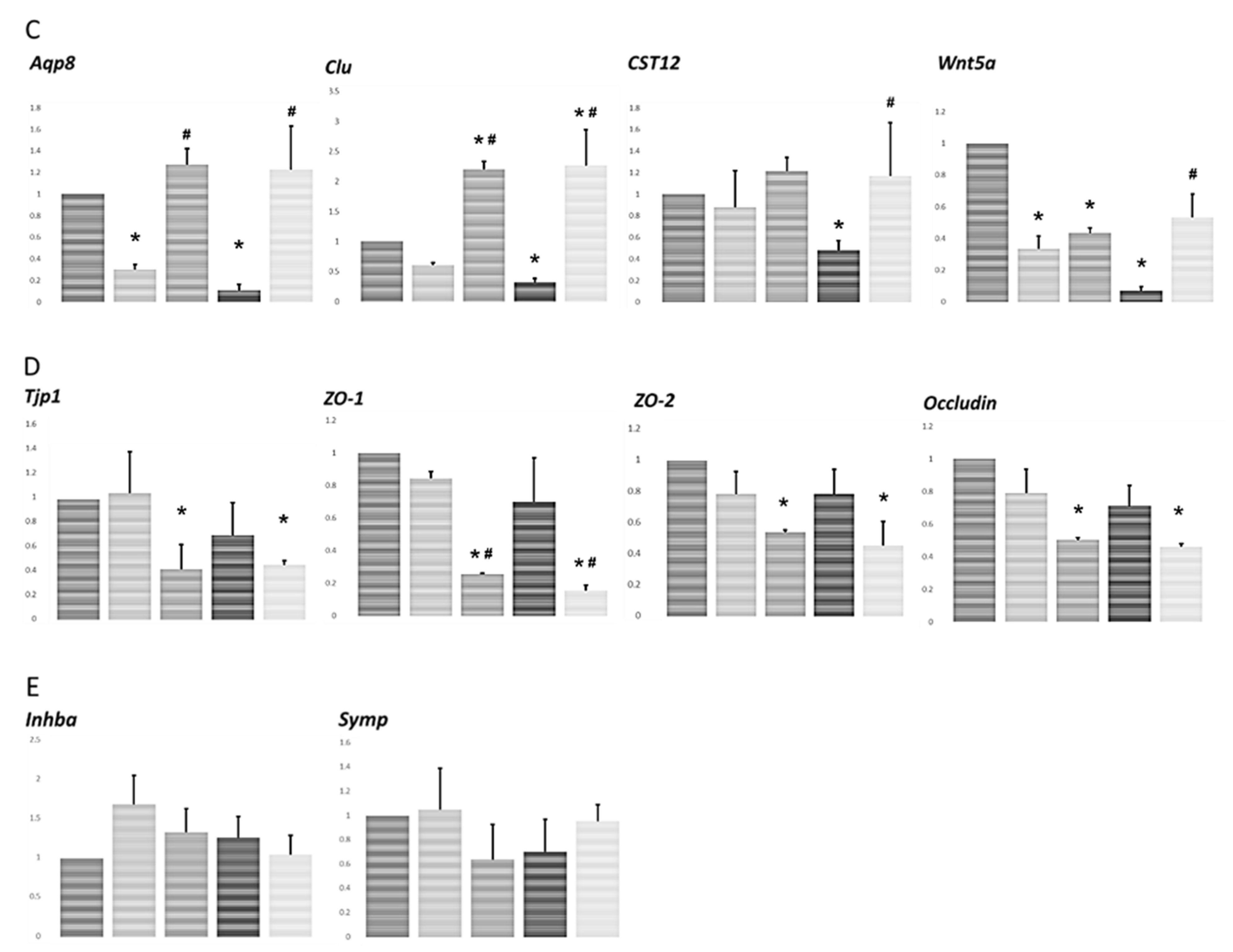
2.5. Analysis of the Specific mRNA Species in Testes Using Real-Time RT-PCR
2.6. Statistical Analysis
3. Results
3.1. Histopathological Changes after Irradiation Exposure
3.2. Effect of Irradiation Exposure on Levels of mRNA Transcripts Encoding Markers of Germ Cell Differentiation Products
3.3. Effect of Irradiation on Levels of mRNA Transcripts Encoding Markers of Sertoli Cell-Specific Products
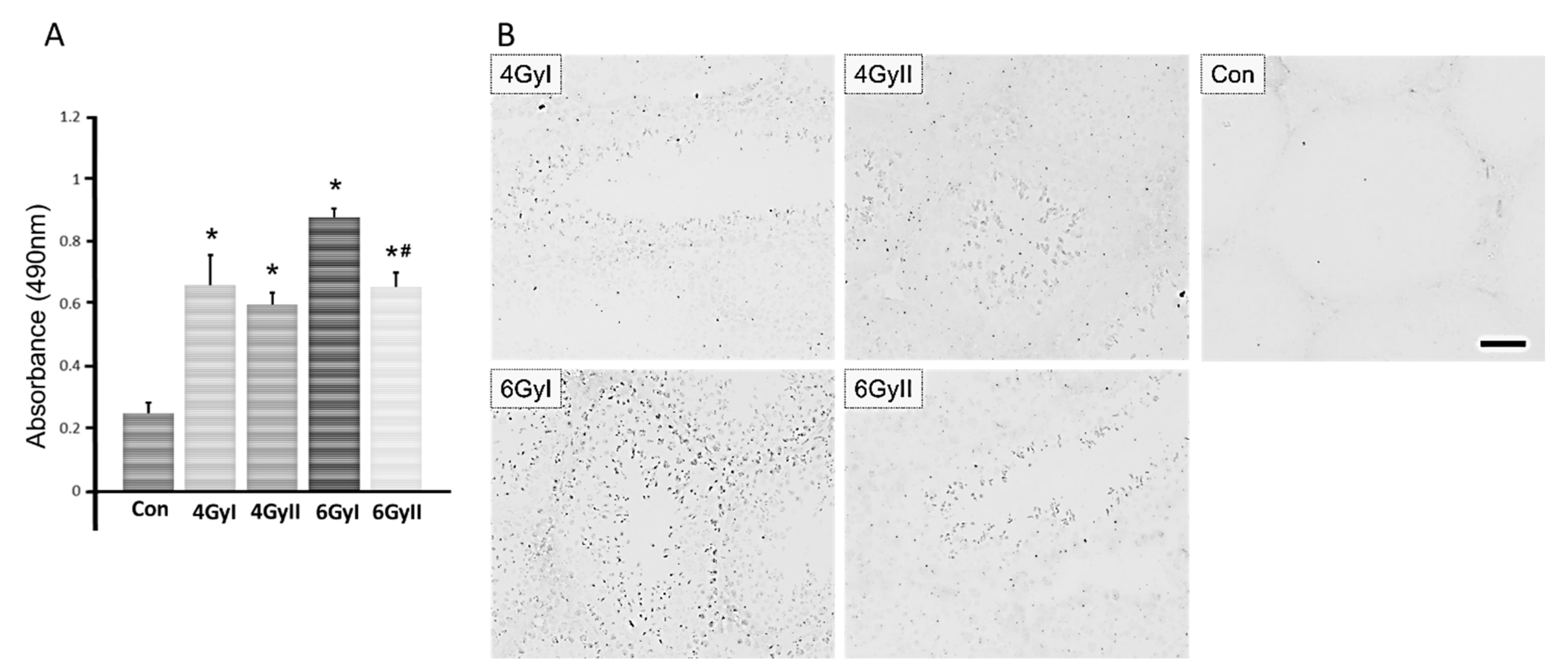
3.4. Detection of Serum Anti-Germ Cell Antibodies
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Qu, N.; Kuramasu, M.; Hirayanagi, Y.; Nagahori, K.; Hayashi, S.; Ogawa, Y.; Terayama, H.; Suyama, K.; Naito, M.; Sakabe, K.; et al. Gosha-Jinki-Gan recovers spermatogenesis in mice with busulfan-induced aspermatogenesis. Int. J. Mol. Sci. 2018, 19, 2606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Nagahori, K.; Qu, N.; Kuramasu, M.; Hirayanagi, Y.; Hayashi, S.; Ogawa, Y.; Hatayama, N.; Terayama, H.; Suyama, K.; et al. The traditional Japanese medicine Goshajinkigan makes a recovery from irradiation-induced aspermatogenesis in mice. BMC Complement. Altern. Med. 2019, 19, 362. [Google Scholar] [CrossRef]
- Kaur, G.; Thompson, L.A.; Dufour, J.M. Sertoli cells–immunological sentinels of spermatogenesis. Semin. Cell Dev. Biol. 2014, 30, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Mital, P.; Hinton, B.T.; Dufour, J.M. The blood-testis and blood-epididymis barriers are more than just their tight junctions. Biol. Reprod. 2011, 84, 851–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plöen, L.; Setchell, B.P. Blood-testis barriers revisited. A homage to Lennart Nicander. Int. J. Androl. 1992, 15, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Qu, N.; Itoh, M.; Sakabe, K. Effects of chemotherapy and radiotherapy on spermatogenesis: The role of testicular immunology (Review). Int. J. Mol. Sci. 2019, 20, 957. [Google Scholar] [CrossRef] [Green Version]
- Sanders, J.E.; Hawley, J.; Levy, W.; Gooley, T.; Buckner, C.D.; Deeg, H.J.; Doney, K.; Storb, R.; Sullivan, K.; Witherspoon, R.; et al. Pregnancies following high-dose cyclophosphamide with or without high-dose busulfan or total-body irradiation and bone marrow transplantation. Blood 1996, 87, 3045–3052. [Google Scholar] [CrossRef]
- Vergouwen, R.P.F.A.; Huiskamp, R.; Bas, R.J.; Roepers-Gajadien, H.L.; de Jong, F.H.; van Eerdenburg, F.J.C.M.; Davids, J.A.G.; de Rooij, D.G. Radiosensitivity of testicular cells in the prepubertal mouse. Radiat. Res. 1994, 139, 316–326. [Google Scholar] [CrossRef]
- Khan, S.; Adhikari, J.S.; Alam Rizvi, M.; Chaudhury, N.K. Radioprotective potential of melatonin against ⁶⁰Co γ-ray-induced testicular injury in male C57BL/6 mice. J. Biomed. Sci. 2015, 22, 61. [Google Scholar] [CrossRef] [Green Version]
- Gabryś, D. Re-irradiation of vertebral bodies. Phys. Medica. 2019, 64, 311–316. [Google Scholar] [CrossRef] [Green Version]
- Hirai, S.; Naito, M.; Terayama, H.; Hatayama, N.; Qu, N.; Musha, M.; Itoh, M. Serum autoantibodies in mice immunized with syngeneic testiscular germs cell alone. Am. J. Reprod. Immunol. 2013, 70, 509–517. [Google Scholar] [CrossRef]
- Yelick, P.C.; Kwon, Y.H.; Flynn, J.F.; Borzorgzadeh, A.; Kleene, K.C.; Hecht, N.B. Mouse transition protein 1 is translationally regulated during the postmeiotic stages of spermatogenesis. Mol. Reprod. Dev. 1989, 1, 193–200. [Google Scholar] [CrossRef]
- Oulad-Abdelghani, M.; Bouillet, P.; Decimo, D.; Gansmuller, A.; Heyberger, S.; Dolle, P.; Bronner, S.; Lutz, Y.; Chambon, P. Characterization of a premeiotic germ cell-specific cytoplasmic protein encoded by Stra8, a novel retinoic acid-responsive gene. J. Cell. Biol. 1996, 135, 469–477. [Google Scholar] [CrossRef]
- Shannon, M.; Richardson, L.; Christian, A.; Handel, M.A.; Thelen, M.P. Differential gene expression of mammalian SPO11/TOP6A homologs during meiosis. FEBS Lett. 1999, 462, 329–334. [Google Scholar] [CrossRef] [Green Version]
- O’Shaughnessy, P.J.; Hu, L.; Baker, P.J. Effect of germ cell depletion on levels of specific mRNA transcripts in mouse Sertoli cells and Leydig cells. Reproduction 2008, 135, 839–850. [Google Scholar] [CrossRef] [Green Version]
- De França, L.R.; Bartke, A.; Borg, K.E.; Oecim, M.; Fadden, C.T.; Yagi, A.; Russell, L.D. Sertoli cells in testes containing or lacking germ cells: A comparative study of paracrine effects using the W (c-kit) gene mutant mouse model. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1994, 240, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Yeh, J.R.; Zhang, X.; Nagano, M.C. Wnt5a Is a Cell-Extrinsic Factor That Supports Self-Renewal of Mouse Spermatogonial Stem Cells. J. Cell Sci. 2011, 124, 2357–2366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, J.; Maheswaran, S.; Donahoe, P.K. Mullerian inhibiting substance: An instructive developmental hormone with diagnostic and possible therapeutic applications. Endocr. Rev. 2001, 22, 657–674. [Google Scholar] [CrossRef]
- Pérez, C.V.; Sobarzo, C.M.; Jacobo, P.V.; Pellizzari, E.H.; Cigorraga, S.B.; Denduchis, B.; Lustig, L. Loss of Occludin Expression and Impairment of Blood-Testis Barrier Permeability in Rats with Autoimmune Orchitis: Effect of Interleukin 6 on Sertoli Cell Tight Junctions. Biol. Reprod. 2012, 87, 122. [Google Scholar] [CrossRef] [PubMed]
- Cimino, I.; Casoni, F.; Liu, X.; Messina, A.; Parkash, J.; Jamin, S.P.; Catteau-Jonard, S.; Collier, F.; Baroncini, M.; Dewailly, D.; et al. Novel role for anti-mullerian hormone in the regulation of GnRH neuron excitability and hormone secretion. Nat. Commun. 2016, 7, 10055. [Google Scholar] [CrossRef] [Green Version]
- Schlatt, S.; von Schönfeldt, V.; Schepers, A.G. Male germ cell transplantation: An experimental approach with a clinical perspective. Br. Med. Bull. 2000, 56, 824–836. [Google Scholar] [CrossRef] [Green Version]
- Radford, J. Restoration of fertility after treatment for cancer. Horm. Res. 2003, 59, 21–23. [Google Scholar] [CrossRef]
- Wallace, W.H.; Thompson, A.B. Preservation of fertility in children treated for cancer. Arch. Dis. Child. 2003, 88, 493–496. [Google Scholar] [CrossRef]
- Takagi, S.; Itoh, N.; Kimura, M.; Sasao, T.; Tsukamoto, T. Spermatogonial proliferation and apoptosis in hypospermatogenesis associated with nonobstructive azoospermia. Fertil. Steril. 2001, 76, 901–907. [Google Scholar] [CrossRef]
- Yakirevich, E.; Sabo, E.; Dirnfeld, M.; Sova, Y.; Spagnoli, G.C.; Resnick, M.B. Morphometrical quantification of spermatogonial germ cells with the 57B anti-MAGE-A4 antibody in the evaluation of testicular biopsies for azoospermia. Appl. Immunohistochem. Mol. Morphol. 2003, 11, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Meistrich, M.L.; Hunter, N.R.; Suzuki, N.; Trostle, P.K.; Withers, H.R. Gradual regeneration of mouse testicular stem cells after exposure to ionizing radiation. Radiat. Res. 1978, 74, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Q.; Cheng, C.Y. A seamless trespass: Germ cell migration across the seminiferous epithelium during spermatogenesis. J. Cell Biol. 2007, 178, 549–556. [Google Scholar] [CrossRef] [Green Version]
- Meinhardt, A.; Hedger, M.P. Immunological, paracrine and endocrine aspects of testicular immune privilege. Mol. Cell Endocrinol. 2011, 335, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.Y.; Mruk, D.D. The blood-testis barrier and its implications for male contraception. Pharmacol. Rev. 2012, 64, 16–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.-W.; Ding, G.-R.; Shi, C.-H.; Zeng, L.-H.; Liu, J.-Y.; Li, J.; Zhao, T.; Chen, Y.-B.; Guo, G.-Z. Mechanisms involved in the blood-testis barrier increased permeability induced by EMP. Toxicology 2010, 276, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Howell, S.J.; Shalet, S.M. Spermatogenesis after cancer treatment: Damage and recovery. J. Natl. Cancer. Inst. Monogr. 2005, 34, 12–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietze, R.; Shihan, M.; Stammler, A.; Konrad, L.; Scheiner-Bobis, G. Cardiotonic steroid ouabain stimulates expression of blood-testis barrier proteins claudin-1 and -11 and formation of tight junctions in Sertoli cells. Mol Cell Endocrinol. 2015, 405, 1–13. [Google Scholar] [CrossRef]
- Komljenovic, D.; Sandhoff, R.; Teigler, A.; Heid, H.; Just, W.W.; Gorgas, K. Disruption of blood-testis barrier dynamics in ether-lipid-deficient mice. Cell Tissue Res. 2009, 337, 281–299. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.E.; Braun, R.E. Germ cell migration across Sertoli cell tight junctions. Science 2012, 338, 798–802. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, P.; William Buaas, F.; Sharma, M.; Smith, B.E.; Greenlee, A.R.; Eacker, S.M.; Braun, R.E. Androgen-dependent sertoli cell tight junction remodeling is mediated by multiple tight junction components. Mol. Endocrinol. 2014, 28, 1055–1072. [Google Scholar] [CrossRef] [Green Version]
- Morrow, C.M.; Tyagi, G.; Simon, L.; Carnes, K.; Murphy, K.M.; Cooke, P.S.; Hofmann, M.-C.C.; Hess, R.A. Claudin 5 expression in mouse seminiferous epithelium is dependent upon the transcription factor ets variant 5 and contributes to blood-testis barrier function. Biol. Reprod. 2009, 81, 871–879. [Google Scholar] [CrossRef] [Green Version]
- Haverfield, J.T.; Meachem, S.J.; Nicholls, P.K.; Rainczuk, K.E.; Simpson, E.R.; Stanton, P.G. Differential permeability of the blood-testis barrier during reinitiation of spermatogenesis in adult male rats. Endocrinology 2014, 155, 1131–1144. [Google Scholar] [CrossRef]
- Gow, A.; Southwood, C.M.; Li, J.S.; Pariali, M.; Riordan, G.P.; Brodie, S.E.; Danias, J.; Bronstein, J.M.; Kachar, B.; Lazzarini, R. CNS myelin and sertoli cell tight junction strands are absent in Osp/claudin-11 null mice. Cell 1999, 99, 649–659. [Google Scholar] [CrossRef] [Green Version]
- Mazaud-Guittot, S.; Meugnier, E.; Pesenti, S.; Wu, X.; Vidal, H.; Gow, A.; Le Magueresse-Battistoni, B. Claudin 11 deficiency in mice results in loss of the sertoli cell epithelial phenotype in the testis. Biol. Reprod. 2009, 109, 202–213. [Google Scholar] [CrossRef] [Green Version]
- Moroi, S.; Saitou, M.; Fujimoto, K.; Sakakibara, A.; Furuse, M.; Ypshida, O.; Tsukita, S. Occludin is concentrated at tight junctions of mouse/rat but not human/guinea pig Sertoli cells in testes. Am. J. Physiol. 1998, 274, C1708–C1717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, M.; Ando-Akatsuka, Y.; Itoh, M.; Furuse, M.; Inazawa, J.; Fujimoto, K.; Tsukita, S. Mammalian occludin in epithelial cells: Its expression and subcellular distribution. Eur. J. Cell Biol. 1997, 73, 222–231. [Google Scholar]
- Grima, J.; Zhu, L.-J.; Cheng, C.Y. Testin is tightly associated with testicular cell membrane upon its secretion by Sertoli cells whose steady-state mRNA level in the testis correlates with the turnover and integrity of inter-testicular cell junctions. J. Biol. Chem. 1997, 272, 6499–6509. [Google Scholar] [CrossRef] [Green Version]
- Calamita, G.; Mazzone, A.; Bizzoca, A.; Svelto, M. Possible involvement of aquaporin-7 and -8 in rat testis development and spermatogenesis. Biochem. Biophys. Res. Commun. 2001, 288, 619–625. [Google Scholar] [CrossRef]
- Kageyama, Y.; Ishibashi, K.; Hayashi, T.; Xia, G.; Sasaki, S.; Kihara, K. Expression of aquaporins 7 and 8 in the developing rat testis. Andrologia 2001, 33, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.; Troedsson, M.H.T.; Rutllant, J. Region-Specific Expression of Aquaporin Subtypes in Equine Testis, Epididymis, and Ductus Deferens. Anat. Evol. Biol. 2013, 296, 1115–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, A.M.; Griswold, M.D. Expression of clusterin/sulfated glycoprotein-2 under conditions of heat stress in rat Sertoli cells and a mouse Sertoli cell line. J. Androl. 1997, 18, 257–263. [Google Scholar] [PubMed]
- Sylvester, S.R.; Morales, C.; Oko, R.; Griswold, M.D. Localization of sulfated glycoprotein-2 (clusterin) on spermatozoa and in the reproductive tract of the male rat. Biol. Reprod. 1991, 45, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Qu, N.; Nagahori, K.; Kuramasu, M.; Ogawa, Y.; Suyama, K.; Hayashi, S.; Sakabe, K.; Itoh, M. Effect of Gosha-Jinki-Gan on levels of specific mRNA transcripts in mouse testes after busulfan treatment. Biomedicines 2020, 8, 432. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| GAPDH | TGTGTCCGTCGTGGATCTGA | TTGCTGTTGAAGTCGCAGGAG |
| Sertoli cell | ||
| Amh | TCCTACATCTGGCTGAAGTGATATGGG | AGGTTCTGTGTGCCCCGCAG |
| Aqp8 | GCTGGCAGTCACAGTGATCGGA | CCTGGACGATGGCAAAGGCTG |
| Ccnd2 | GGAACCTGGCCGCAGTCACC | AATCATCGACGGCGGGTACATG |
| Claudin-11 | TCACAACGTCCACCAATGACTG | GGCACATACAGGAAACCAGATG |
| Clu | CCACGCCATGAAGATTCTCCTGC | CTCCCTGGACGGCGTTCTGA |
| Cst12 | GGATGACGATTTTGCCTACAAGTTCCT | TTCTCTCTCCTGGACCTTCCTGCA |
| Cst9 | GATATTTGCCCCTTTCAGGAGAGCC | AGAGAAGTACGTGACCAGTCCATGGG |
| Dhh | GGCGCAGACCGCCTGATG | AAGGCACGGCCTTCGTAGTGG |
| Espn | GCTTCTGGTCGGGCATTACCCT | GTGTCATGCCGTCTTGGGCG |
| Fshr | GGCCAGGTCAACATACCGCTTG | TGCCTTGAAATAGACTTGTTGCAAATTG |
| Fyn | GAAGCGGCCCTGTATGGAAGGTT | TGTGGGCAGGGCATCCTATAGC |
| GATA1 | ATGGTCAGAACCGGCCTCTCATC | GAGCTTGAAATAGAGGCCGCAGG |
| Inhba | CATGGAGCAGACCTCGGAGATCA | TGGTCCTGGTTCTGTTAGCCTTGG |
| Inhbb | GAGCGCGTCTCCGAGATCATCA | CGTACCTTCCTCCTGCTGCCCTT |
| Msi1 | TCACTTTCATGGACCAGGCGG | GTTCACAGACAGCCCCCCCA |
| Occlidin | CTTCTGCTTCATCGCTTCC | CTTGCCCTTTCCTGCTTTC |
| Rhox5 | AGGTTCGCCCAGCATCGACTG | GCCGCAGCCCTCCTGATCTT |
| Shbg | GACATTCCCCAGCCTCATGCA | TGCCTCGGAAGACAGAACCACG |
| Sox9 | CGCGGAGCTCAGCAAGACTCTG | TGTCCGTTCTTCACCGACTTCCTC |
| Spata2 | GCCGTGTGGGCCTGTGCTT | TTCCCCAAATCAAACCCAAGGG |
| sympk | CAAGAAGAAGGGCCAAGCATCGA | AGGAAGTTGTCAAGCAGGGTGGG |
| Testin | AAAGACAATGGCGGCCTCGC | GGCCCCACTTTAGCCACTGCC |
| Tjp1 | GCGGAGAGAGACAAGATGTCCGC | CTCTGAAAATGAGGAT- TATCTCTTCCACCA |
| Trf | CAAATGCATCAGCTTCCGTGACC | CGGCATCGTACACCCAACCC |
| Wnt5a | CTGCTTCTACCATGCGTTTGCTGG | GCCATGGGACAGTGCGGC |
| Wt1 | GCTCCAGCTCAGTGAAATGGACAGAA | GGCCACTCCAGATACACGCCG |
| ZO-1 | ACAAACAGCCCTACCAACC | CCATCCTCATCTTCATCTTCTTC |
| ZO-2 | GTTTTTCTTCGTCCTAGTCCC | CATCCATCCCTTCCATCTTTC |
| Germ Cell | ||
| Acrosin | TGTCCGTGGTTGCCAAGGATAACA | AATCCGGGTACCTGCTTGTGAGTT |
| Boule | AACCCAACAAGTGGCCCAAGATAC | CTTTGGACACTCCAGCTCTGTCAT |
| c-Kit | GCATCACCATCAAAAACGTG | GATAGTCAGCGTCTCCTGGC |
| CREM-1 | TTCTTTCACGAAGACCCTCA | TGTTAGGTGGTGTCCCTTCT |
| Ddx4 | AAGCAGAGGGTTTTCCAAGC | GCCTGATGCTTCTGAATCG |
| Gfr-α1 | CAGTTTTCGTCTGCTGAGGTTG | TCTGCTCAAAGTGGCTCCAT |
| OCT4 | TGCGGAGGGATGGCATACTG | GCACAGGGCTCAGAGGAGGTTC |
| Protamine | TCCATCAAAACTCCTGCGTGA | AGGTGGCATTGTTCCTTAGCA |
| Spo11 | CGCGTGGCCTCTAGTTCTGAGG | GGTATCATCCGAAGGCCGACAGAAT |
| Stra8 | GAAGGTGCATGGTTCACCGTGG | GCTCGATCGGCGGGCCTGTG |
| Sycp3 | TGGAGCTGACATCAACAAAGC | CCCACTGCTGCAACACATTC |
| Tnp1 | GGCGATGATGCAAGTCGCAA | CCACTCTGATAGGATCTTTGGCTTTTGG |
| Zbtb16 | AACGGTTCCTGGACAGTTTG | CCACACAGCAGACAGAAGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagahori, K.; Qu, N.; Kuramasu, M.; Ogawa, Y.; Kiyoshima, D.; Suyama, K.; Hayashi, S.; Sakabe, K.; Yoshimoto, T.; Itoh, M. Changes in Expression of Specific mRNA Transcripts after Single- or Re-Irradiation in Mouse Testes. Genes 2022, 13, 151. https://doi.org/10.3390/genes13010151
Nagahori K, Qu N, Kuramasu M, Ogawa Y, Kiyoshima D, Suyama K, Hayashi S, Sakabe K, Yoshimoto T, Itoh M. Changes in Expression of Specific mRNA Transcripts after Single- or Re-Irradiation in Mouse Testes. Genes. 2022; 13(1):151. https://doi.org/10.3390/genes13010151
Chicago/Turabian StyleNagahori, Kenta, Ning Qu, Miyuki Kuramasu, Yuki Ogawa, Daisuke Kiyoshima, Kaori Suyama, Shogo Hayashi, Kou Sakabe, Takayuki Yoshimoto, and Masahiro Itoh. 2022. "Changes in Expression of Specific mRNA Transcripts after Single- or Re-Irradiation in Mouse Testes" Genes 13, no. 1: 151. https://doi.org/10.3390/genes13010151