
SESN3 Inhibited SMAD3 to Relieve Its Suppression for MiR-124, Thus Regulating Pre-Adipocyte Adipogenesis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Animals
2.2. Cell Culture, Transfection and Differentiation
2.3. RNA Isolation, Library Preparation, and RT-PCR
2.4. Oil Red O Staining and Triglyceride Assay
2.5. Cell Counting Kit-8 (CCK-8) Assay
2.6. Cell Division Assay by EdU Incubation
2.7. Luciferase Reporter Assay
2.8. Chromatin Immunoprecipitation Assay PCR (ChIP-PCR)
2.9. Western Blotting
2.10. Bioinformatics Analysis
2.11. Statistical Analysis
3. Results
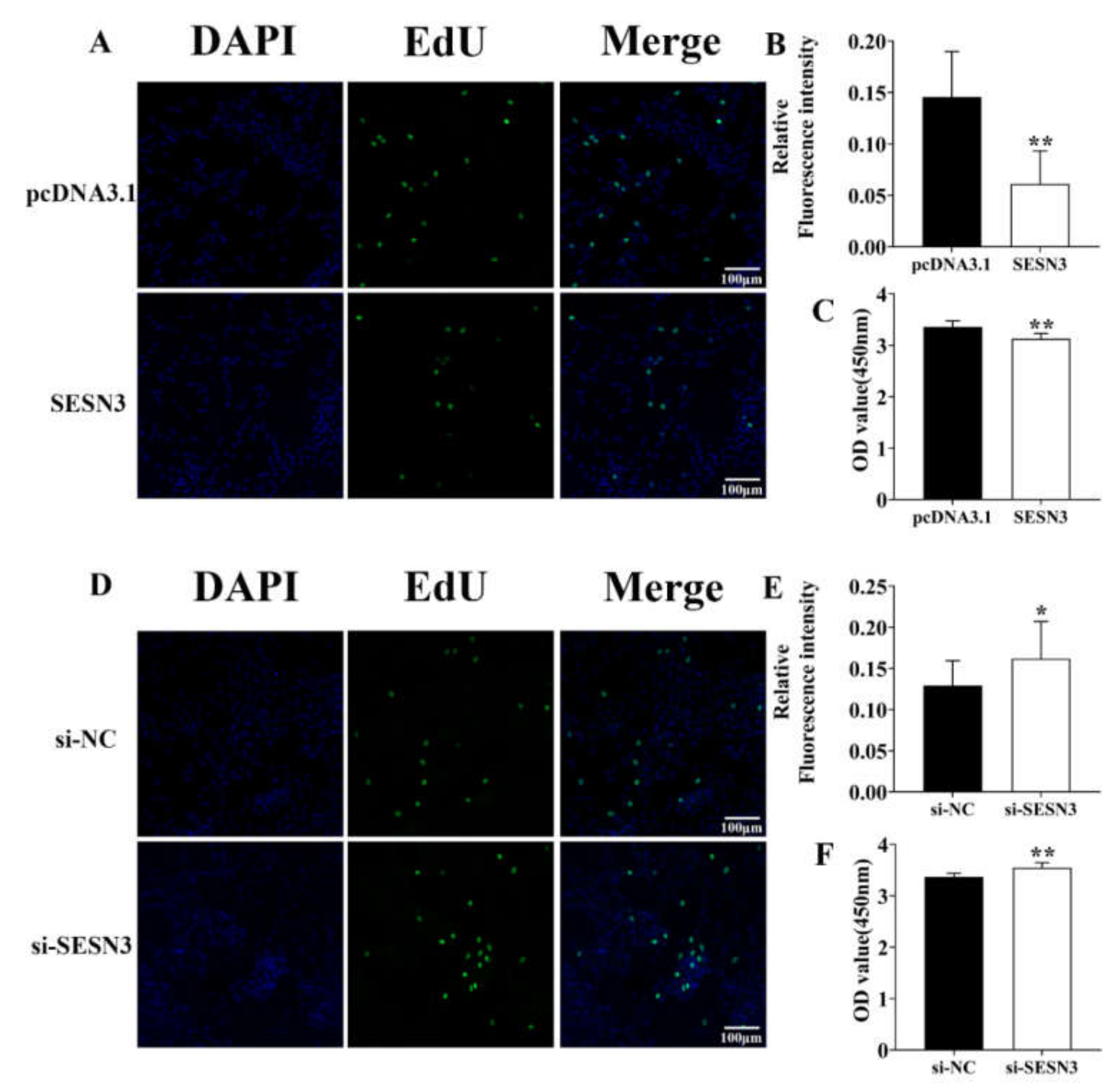
3.1. Identification of the SESN3 Proliferative Role in the Pre-Adipocyte
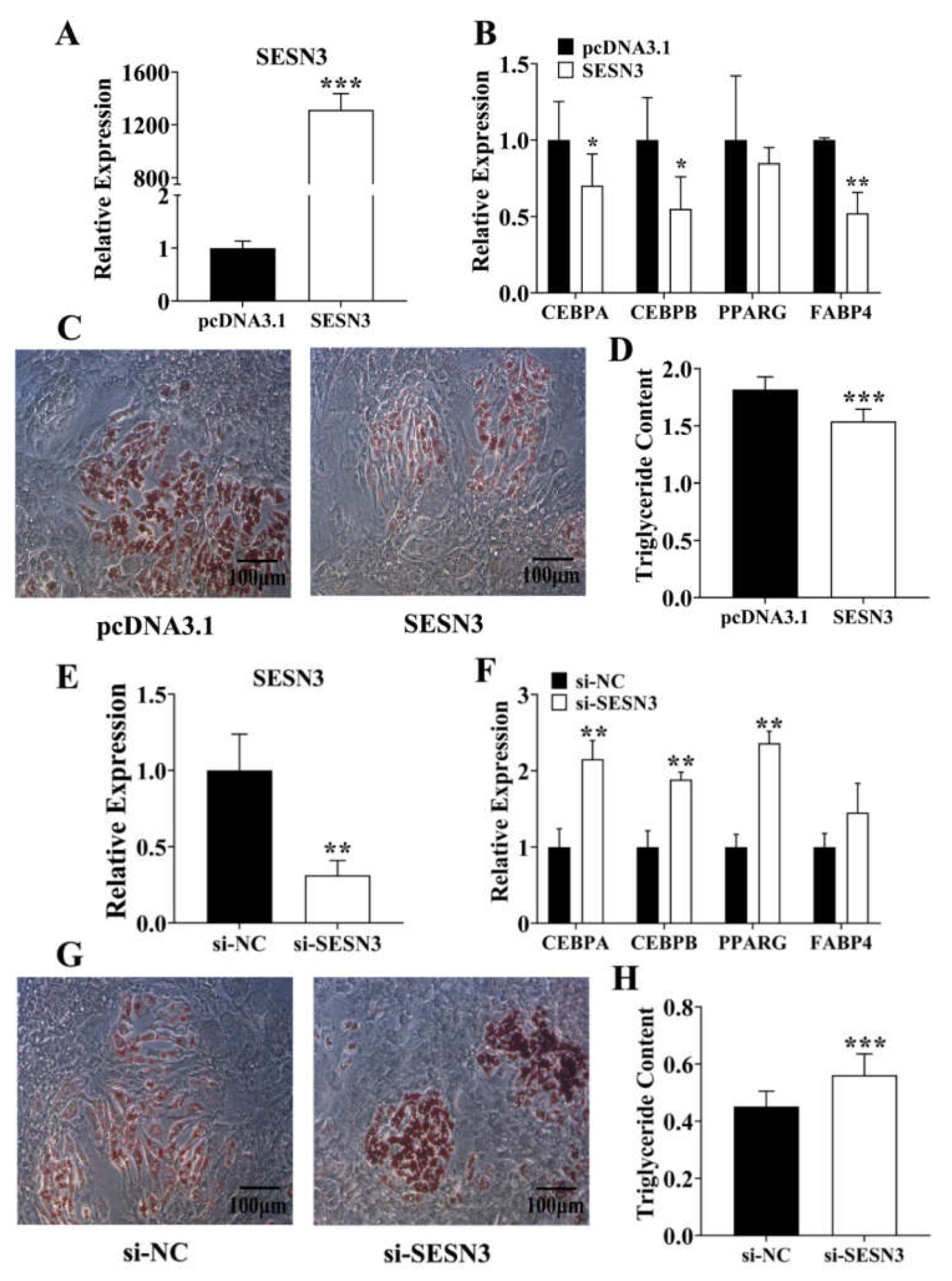
3.2. Sesn3 Inhibits Pre-Adipocyte Adipogenesis
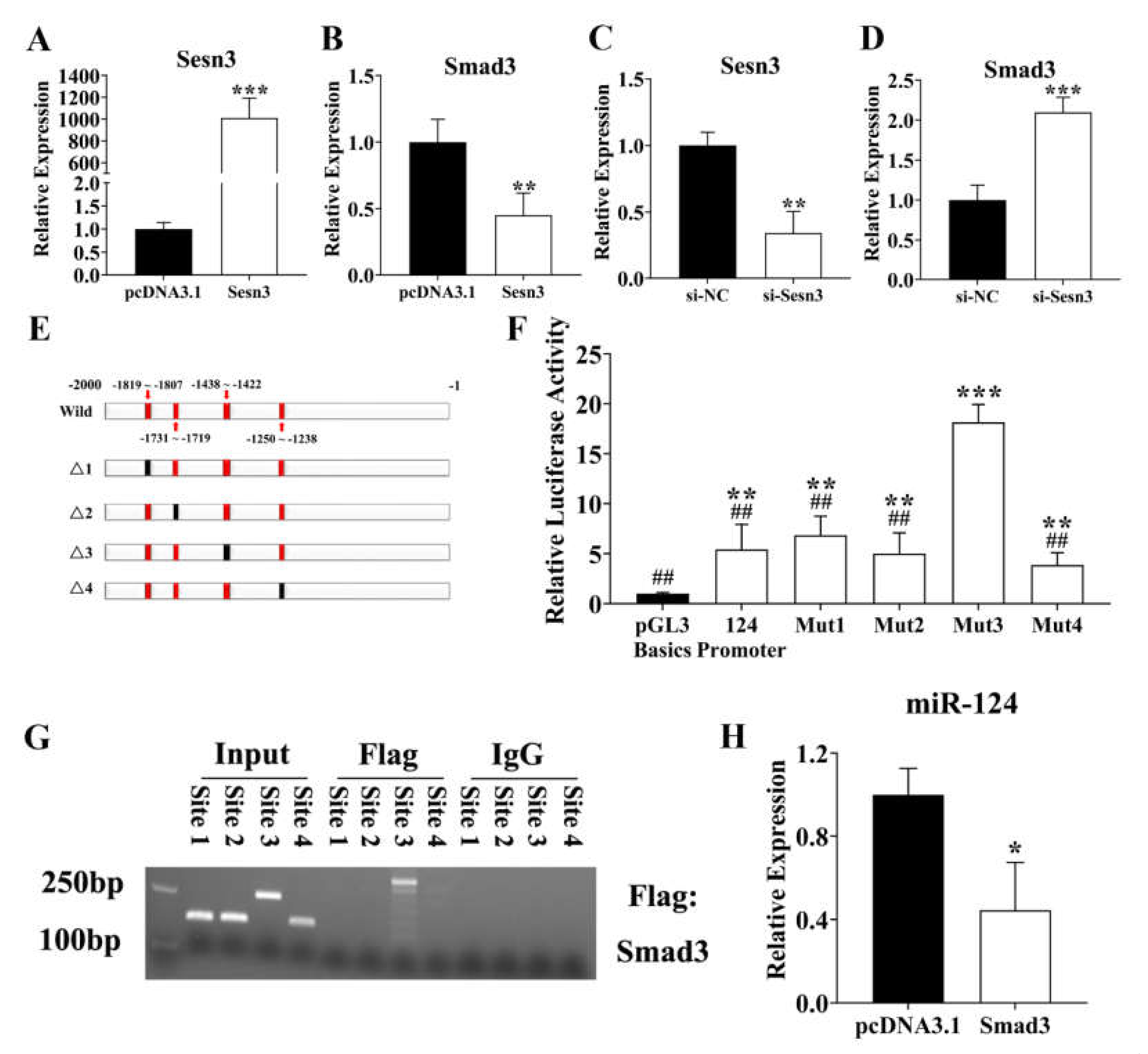
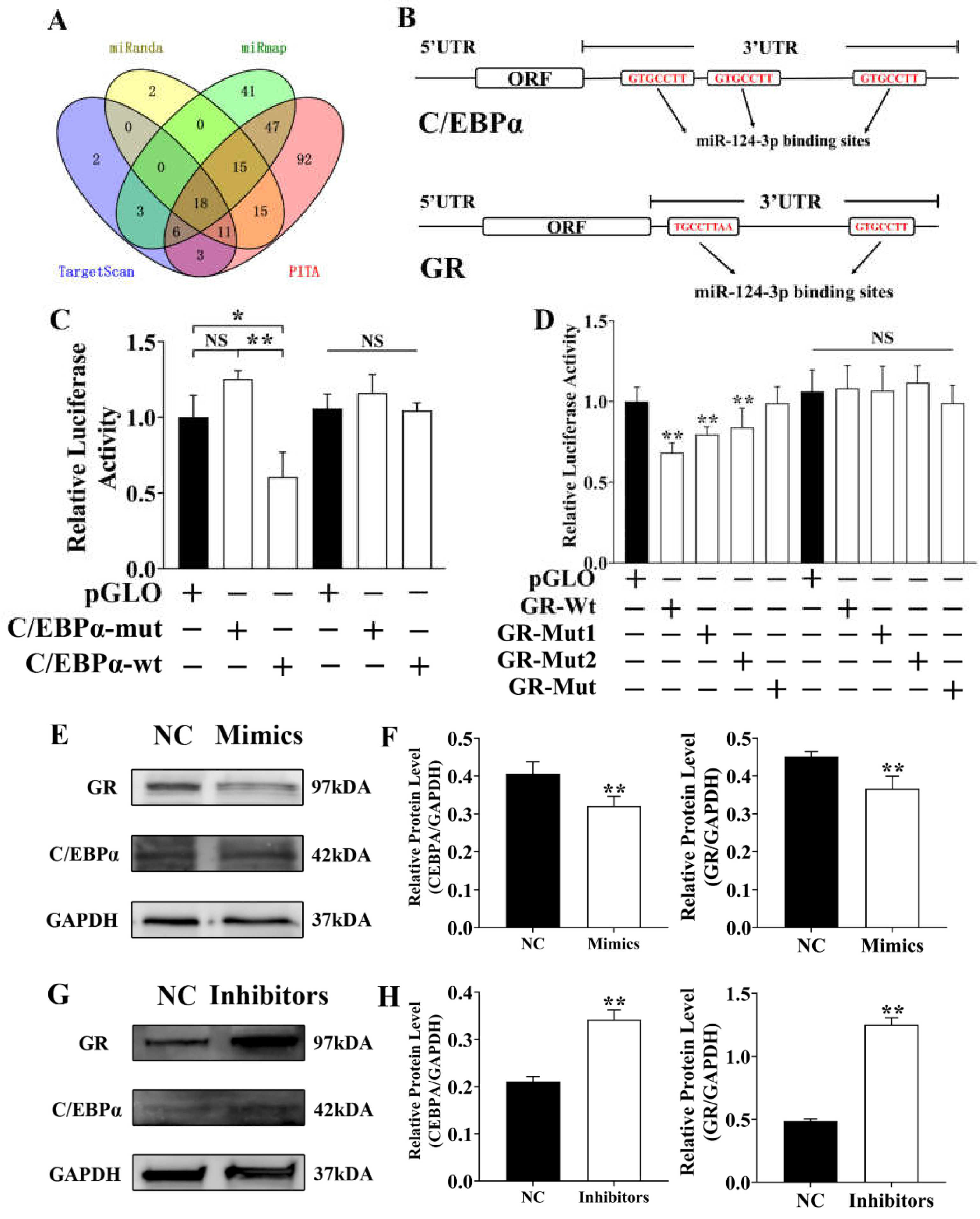
3.3. SESN3 Inhibits SMAD3 to Reduce Its Suppressing Effect on ssc-miR-124 Transcription
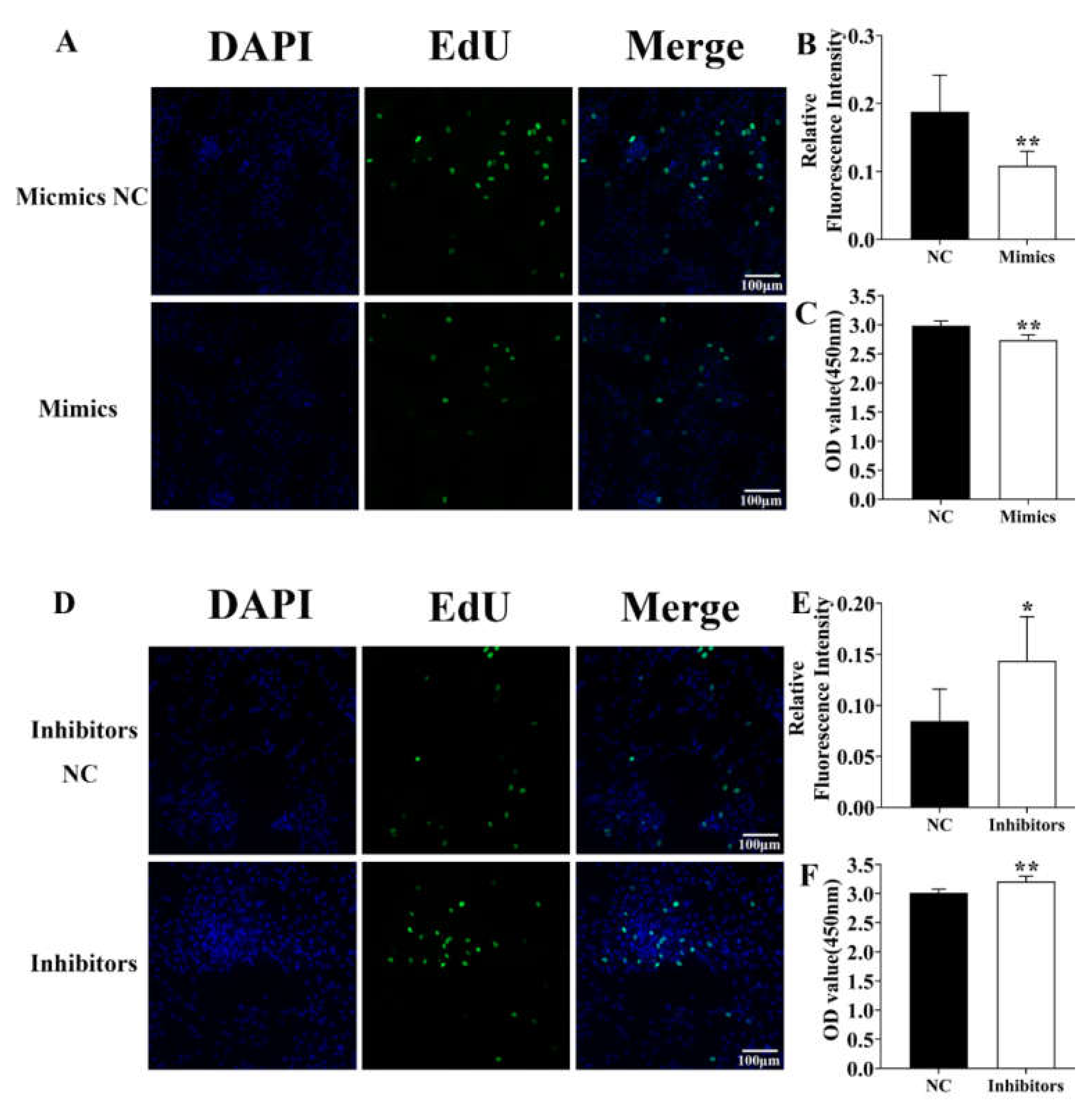
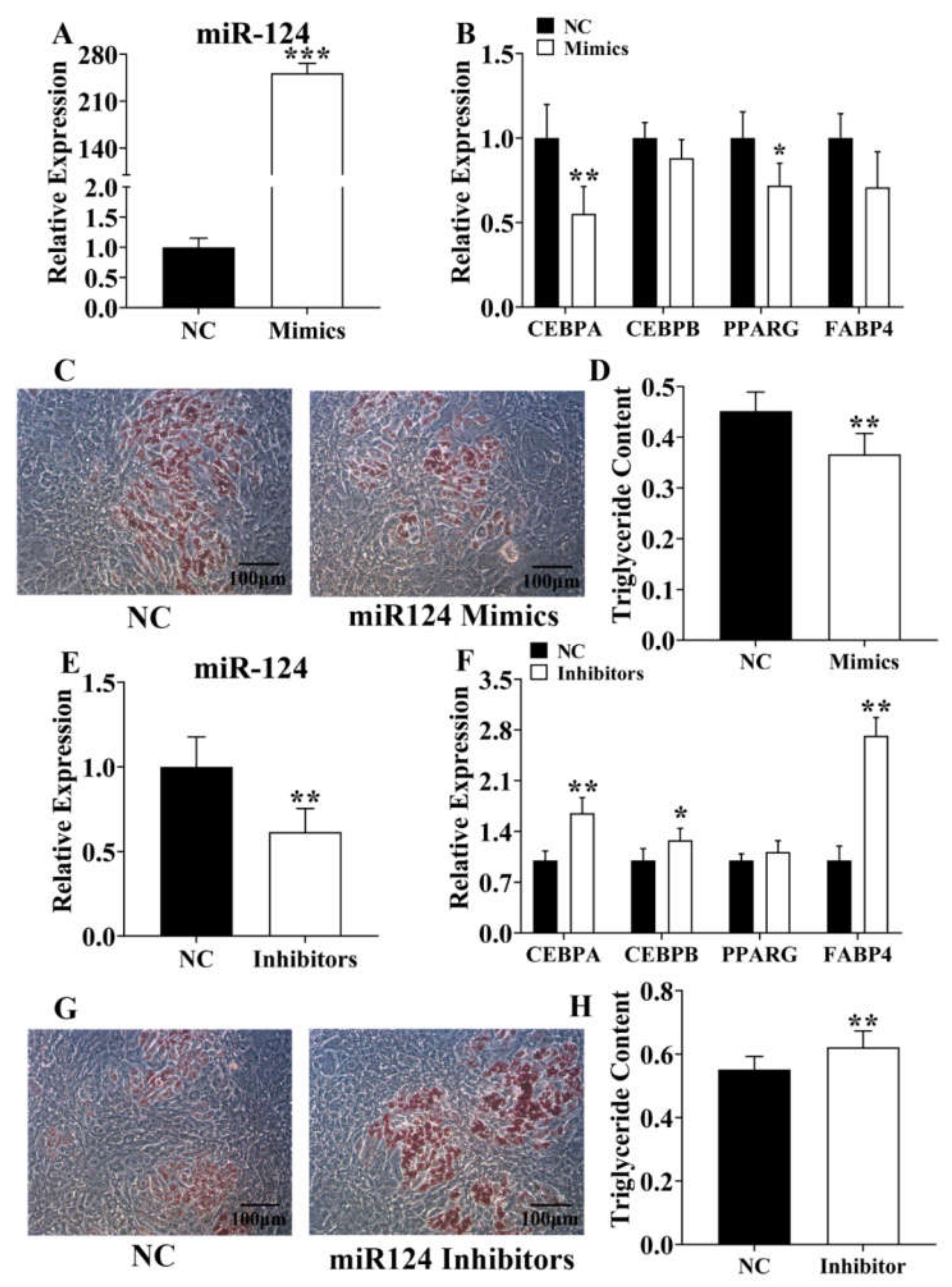
3.4. Identification of ssc-miR-124 Proliferative Role in the Pre-Adipocyte
3.5. Ssc-miR-124 Inhibits Pre-Adipocyte Adipogenesis
3.6. Ssc-miR-124 Inhibits C/EBPα and GR by Targeting Their 3′UTR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Goran, M.I.; Ball, G.D.C.; Cruz, M.L. Obesity and Risk of Type 2 Diabetes and Cardiovascular Disease in Children and Adolescents. J. Clin. Endocrinol. Metab. 2003, 88, 1417–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopelman, P.G. Obesity as a medical problem. Nat. Cell Biol. 2000, 404, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Li, Q.; Wang, J.; Zhao, Y.; Niu, L.; Bai, L.; Shuai, S.; Li, X.; Zhang, S.; Zhu, L. miR-144-3p Promotes Adipogenesis Through Releasing C/EBPα From Klf3 and CtBP. Front. Genet. 2018, 9, 677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza-Mello, V. Peroxisome proliferator-activated receptors as targets to treat non-alcoholic fatty liver disease. World J. Hepatol. 2015, 7, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Barroso, I.; Gurnell, M.; Crowley, V.; Agostini, M.; Schwabe, J.; Soos, M.; Maslen, G.; Williams, T.; Lewis, H.; Schafer, A.; et al. Dominant negative mutations in human PPARgamma associated with severe insulin resistance, diabetes mellitus and hypertension. Nat. Commun. 1999, 402, 880–883. [Google Scholar] [CrossRef] [PubMed]
- Doney, A.; Fischer, B.; Leese, G.; Morris, A.; Palmer, C. Cardiovascular risk in type 2 diabetes is associated with variation at the PPARG locus: A Go-DARTS study. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 2403–2407. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Molina, A.; Efeyan, A.; Lopez-Guadamillas, E.; Muñoz-Martin, M.; López, G.G.; Cañamero, M.; Mulero, F.; Pastor, J.; Martinez, S.; Romanos, E.; et al. Pten Positively Regulates Brown Adipose Function, Energy Expenditure, and Longevity. Cell Metab. 2012, 15, 382–394. [Google Scholar] [CrossRef] [Green Version]
- Hamm, J.; Park, B.; Farmer, S. A role for C/EBPbeta in regulating peroxisome proliferator-activated receptor Gamma activity during adipogenesis in 3T3-L1 preadipocytes. J. Biol. Chem. 2001, 276, 18464–18471. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; He, K.; Wang, L.; Hu, J.; Gu, J.; Zhou, C.; Lu, R.; Jin, Y. Stk40 represses adipogenesis through translational control of CCAAT/enhancer-binding proteins. J. Cell Sci. 2015, 128, 2881–2890. [Google Scholar] [CrossRef] [Green Version]
- Lefterova, M.; Lazar, M. New developments in adipogenesis. Trends Endocrinol. Metab. TEM 2009, 20, 107–114. [Google Scholar] [CrossRef]
- Student, A.; Hsu, R.; Lane, M. Induction of fatty acid synthetase synthesis in differentiating 3T3-L1 preadipocytes. J. Biol. Chem. 1980, 255, 4745–4750. [Google Scholar] [CrossRef]
- Vu, D.; Ong, J.M.; Clemens, T.L.; Kern, P.A. 1,25-Dihydroxyvitamin D induces lipoprotein lipase expression in 3T3-L1 cells in association with adipocyte differentiation. Endocrinology 1996, 137, 1540–1544. [Google Scholar] [CrossRef]
- Hamilton, A.J.; Baulcombe, D.C. A Species of Small Antisense RNA in Posttranscriptional Gene Silencing in Plants. Science 1999, 286, 950–952. [Google Scholar] [CrossRef] [Green Version]
- Ketting, R.F.; Fischer, S.E.; Bernstein, E.; Sijen, T.; Hannon, G.J.; Plasterk, R.H. Dicer functions in RNA interference and in synthesis of small RNA involved in developmental timing in C. elegans. Genes Dev. 2001, 15, 2654–2659. [Google Scholar] [CrossRef] [Green Version]
- Grishok, A.; Pasquinelli, A.E.; Conte, D.; Li, N.; Parrish, S.; Ha, I.; Baillie, D.L.; Fire, A.; Ruvkun, G.; Mello, C.C. Genes and mechanisms related to RNA interference regulate expression of the small temporal RNAs that control C. elegans developmental timing. Cell 2001, 106, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Wu, J.; Chen, W.; Tang, S.; Mo, Z.; Tang, Y.; Li, Y.; Wang, J.; Liu, X.; Peng, J.; et al. MicroRNA-27a/b regulates cellular cholesterol efflux, in-flux and esterification/hydrolysis in THP-1 macrophages. Atherosclerosis 2014, 234, 54–64. [Google Scholar] [CrossRef]
- Pan, S.; Yang, X.; Jia, Y.; Li, R.; Zhao, R. Microvesicle-shuttled miR-130b reduces fat deposition in recipient primary cul-tured porcine adipocytes by inhibiting PPAR-γ expression. J. Cell. Physiol. 2014, 229, 631–639. [Google Scholar] [CrossRef]
- Li, M.; Liu, Z.; Zhang, Z.; Liu, G.; Sun, S.; Sun, C. miR-103 promotes 3T3-L1 cell adipogenesis through AKT/mTOR signal pathway with its target being MEF2D. Biol. Chem. 2015, 396, 235–244. [Google Scholar] [CrossRef]
- Fu, T.; Seok, S.; Choi, S.; Huang, Z.; Suino-Powell, K.; Xu, H.E.; Kemper, B.; Kemper, J.K. MicroRNA 34a Inhibits Beige and Brown Fat Formation in Obesity in Part by Suppressing Adipocyte Fibroblast Growth Factor 21 Signaling and SIRT1 Function. Mol. Cell. Biol. 2014, 34, 4130–4142. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Gu, S.; Chu, W.; Sun, W.; Wei, W.; Dang, X.; Tian, Y.; Liu, K.; Chen, J. miR-17-5p Regulates Differential Expression of NCOA3 in Pig Intramuscular and Subcutaneous Adipose Tissue. Lipids 2017, 52, 939–949. [Google Scholar] [CrossRef]
- Liu, K.; Zhang, X.; Wei, W.; Liu, X.; Tian, Y.; Han, H.; Zhang, L.; Wu, W.; Chen, J. Myostatin/SMAD4 signaling-mediated reg-ulation of miR-124-3p represses glucocorticoid receptor expression and inhibits adipocyte differentiation. Am. J. Physiol.-Endocrinol. Metab. 2019, 316, E635–E645. [Google Scholar]
- Lin, W.; Tang, Y.; Zhao, Y.; Zhao, J.; Zhang, L.; Wei, W.; Chen, J. MiR-144-3p targets FoxO1 to reduce its regulation of adi-ponectin and promote adipogenesis. Front. Genet. 2020, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ho, A.; Cho, C.-S.; Namkoong, S.; Cho, U.-S.; Lee, J.H. Biochemical Basis of Sestrin Physiological Activities. Trends Biochem. Sci. 2016, 41, 621–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, M.; Kim, H.; Zhong, X.; Dong, C.; Zhang, B.; Fang, Z.; Zhang, Y.; Lu, X.; Saxena, R.; Liu, Y.; et al. Sestrin 3 Protects Against Diet-Induced Nonalcoholic Steatohepatitis in Mice Through Suppression of Transform-ing Growth Factor β Signal Transduction. Hepatology 2020, 71, 76–92. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Shimkus, K.; Lacko, H.; Kutzler, L.; Jefferson, L.; Kimball, S. Evidence for a role for Sestrin1 in mediating leu-cine-induced activation of mTORC1 in skeletal muscle. Am. J. Physiol.-Endocrinol. Metab. 2019, 316, E817–E828. [Google Scholar]
- Wolfson, R.L.; Chantranupong, L.; Saxton, R.A.; Shen, K.; Scaria, S.M.; Cantor, J.R.; Sabatini, D.M. Sestrin2 is a leucine sensor for the mTORC1 pathway. Science 2015, 351, 43–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.; Li, M.; Zhang, K.; Chen, L.; Jiang, A.-A.; Wang, J.; Lv, X.; Li, X. Identification of suitable endogenous control microRNA genes in normal pig tissues. Anim. Sci. J. 2011, 82, 722–728. [Google Scholar] [CrossRef]
- Timoneda, O.; Balcells, I.; Córdoba, S.; Castelló, A.; Sánchez, A. Determination of reference microRNAs for relative quantification in porcine tissues. PLoS ONE 2012, 7, e44413. [Google Scholar]
- Bae, I.-S.; Seo, K.-S.; Kim, S.H. Identification of endogenous microRNA references in porcine serum for quantitative real-time PCR normalization. Mol. Biol. Rep. 2018, 45, 943–949. [Google Scholar] [CrossRef]
- Harrison, D.E.; Strong, R.; Sharp, Z.D.; Nelson, J.F.; Astle, C.M.; Flurkey, K.; Nadon, N.L.; Wilkinson, J.E.; Frenkel, K.; Carter, C.S.; et al. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature 2009, 460, 392–395. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.; Rabinovitch, P.S.; Kaeberlein, M. mTOR is a key modulator of ageing and age-related disease. Nat. Cell Biol. 2013, 493, 338–345. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Budanov, A.V.; Karin, M. Sestrins Orchestrate Cellular Metabolism to Attenuate Aging. Cell Metab. 2013, 18, 792–801. [Google Scholar] [CrossRef] [Green Version]
- Budanov, A.V.; Karin, M. p53 Target Genes Sestrin1 and Sestrin2 Connect Genotoxic Stress and mTOR Signaling. Cell 2008, 134, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.-L.; Loh, K.-S.; Liou, B.-Y.; Chu, I.-H.; Kuo, C.-J.; Chen, H.-D.; Chen, C.-S. SESN-1 is a positive regulator of lifespan in Caenorhabditis elegans. Exp. Gerontol. 2013, 48, 371–379. [Google Scholar] [CrossRef]
- Lee, J.H.; Budanov, A.V.; Park, E.J.; Birse, R.; Kim, T.E.; Perkins, G.A.; Ocorr, K.; Ellisman, M.H.; Bodmer, R.; Bier, E.; et al. Sestrin as a Feedback Inhibitor of TOR That Prevents Age-Related Pathologies. Science 2010, 327, 1223–1228. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Budanov, A.V.; Talukdar, S.; Park, E.J.; Park, H.L.; Park, H.-W.; Bandyopadhyay, G.; Li, N.; Aghajan, M.; Jang, I.; et al. Maintenance of Metabolic Homeostasis by Sestrin2 and Sestrin3. Cell Metab. 2012, 16, 311–321. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.C. The potential of sestrins as therapeutic targets for diabetes. Expert Opin. Ther. Targets 2015, 19, 1011–1015. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Kowalsky, A.H.; Lee, J.H. Sestrins in Physiological Stress Responses. Annu. Rev. Physiol. 2021, 83, 381–403. [Google Scholar] [CrossRef]
- Brier, A.-S.B.; Loft, A.; Madsen, J.G.S.; Rosengren, T.; Nielsen, R.; Schmidt, S.F.; Liu, Z.; Yan, Q.; Gronemeyer, H.; Mandrup, S. The KDM5 family is required for activation of pro-proliferative cell cycle genes during adipocyte differentiation. Nucleic Acids Res. 2017, 45, 1743–1759. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.-M.; Ho, C.-K.; Gao, Y.; Chong, C.-H.; Zheng, D.-N.; Zhang, Y.-F.; Yu, L. Salvianolic acid-B improves fat graft survival by promoting proliferation and adipogenesis. Stem Cell Res. Ther. 2021, 12, 1–16. [Google Scholar] [CrossRef]
- Chen, K.; Zhang, J.; Liang, F.; Zhu, Q.; Cai, S.; Tong, X.; He, Z.; Liu, X.; Chen, Y.; Mo, D. HMGB2 orchestrates mitotic clonal expansion by binding to the promoter of C/EBPβ to facilitate adipogenesis. Cell Death Dis. 2021, 12, 666. [Google Scholar] [CrossRef]
- Qadir, A.S.; Woo, K.M.; Ryoo, H.-M.; Baek, J.-H. Insulin suppresses distal-less homeobox 5 expression through the up-regulation of microRNA-124 in 3T3-L1 cells. Exp. Cell Res. 2013, 319, 2125–2134. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Umek, R.M.; McKnight, S.L. Regulated expression of three C/EBP isoforms during adipose conversion of 3T3-L1 cells. Genes Dev. 1991, 5, 1538–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, N.Z.; Cidlowski, J.A. Translational Regulatory Mechanisms Generate N-terminal Glucocorticoid Receptor Isoforms with Unique Transcriptional Target Genes. Mol. Cell 2005, 18, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Infante, M.; Armani, A.; Mammi, C.; Fabbri, A.; Caprio, M. Impact of Adrenal Steroids on Regulation of Adipose Tissue. Compr. Physiol. 2011, 7, 1425–1447. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, W.; Zhao, J.; Yan, M.; Li, X.; Yang, K.; Wei, W.; Zhang, L.; Chen, J. SESN3 Inhibited SMAD3 to Relieve Its Suppression for MiR-124, Thus Regulating Pre-Adipocyte Adipogenesis. Genes 2021, 12, 1852. https://doi.org/10.3390/genes12121852
Lin W, Zhao J, Yan M, Li X, Yang K, Wei W, Zhang L, Chen J. SESN3 Inhibited SMAD3 to Relieve Its Suppression for MiR-124, Thus Regulating Pre-Adipocyte Adipogenesis. Genes. 2021; 12(12):1852. https://doi.org/10.3390/genes12121852
Chicago/Turabian StyleLin, Weimin, Jindi Zhao, Mengting Yan, Xuexin Li, Kai Yang, Wei Wei, Lifan Zhang, and Jie Chen. 2021. "SESN3 Inhibited SMAD3 to Relieve Its Suppression for MiR-124, Thus Regulating Pre-Adipocyte Adipogenesis" Genes 12, no. 12: 1852. https://doi.org/10.3390/genes12121852