
Genome Survey Sequencing of an Iconic ‘Trophy’ Sportfish, the Roosterfish Nematistius pectoralis: Genome Size, Repetitive Elements, Nuclear RNA Gene Operon, and Microsatellite Discovery


Abstract
:1. Introduction
2. Material and Methods
2.1. Sampling and DNA Extraction
2.2. Library Preparation and Sequencing
2.3. Genome Size Estimation in Nematistius pectoralis
2.4. Repetitive Elements in the Nuclear Genome of Nematistius pectoralis
2.5. Nuclear Ribosomal Operon in Nematistius pectoralis
2.6. Microsatellite Discovery in Nematistius pectoralis
3. Results and Discussion
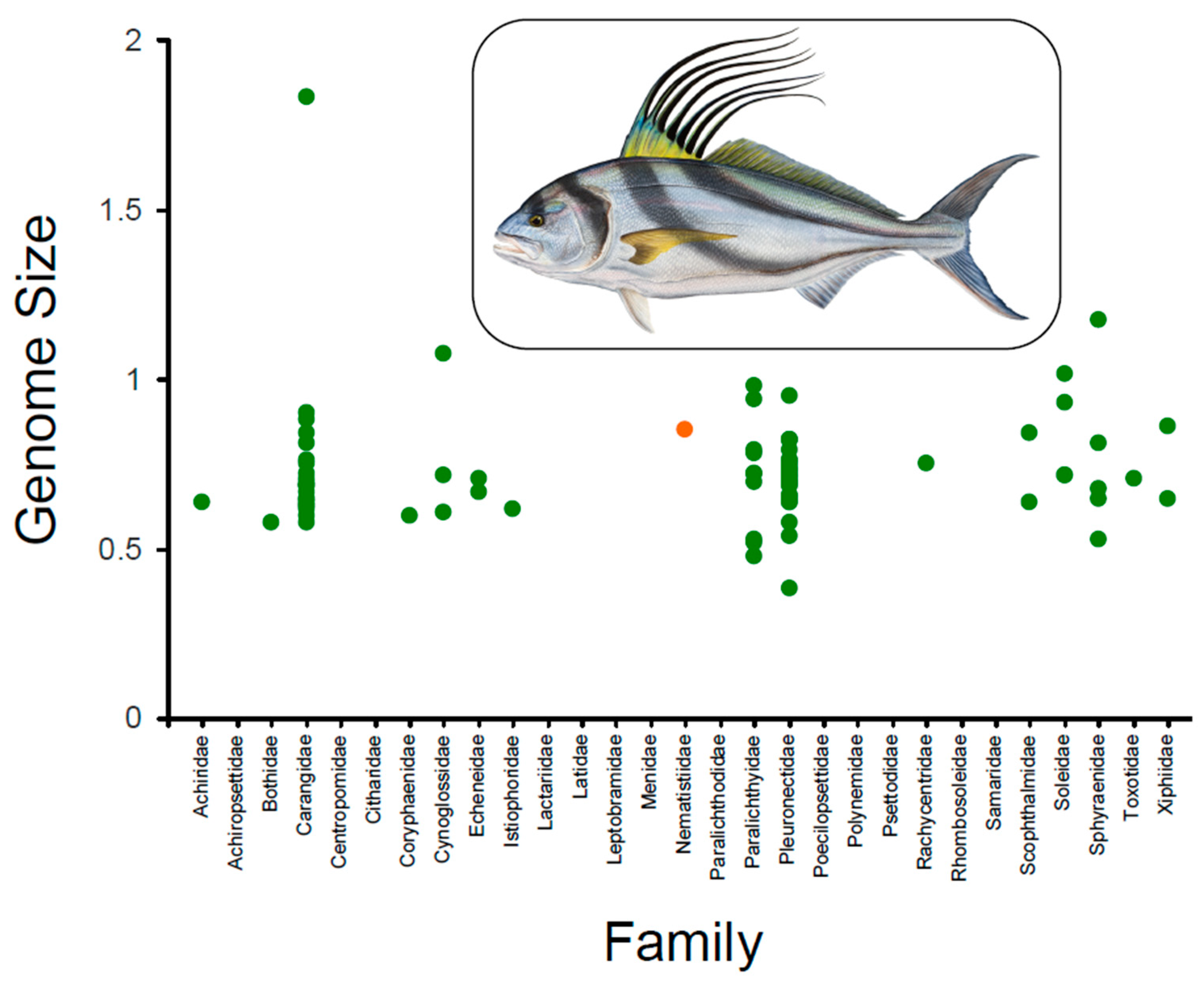
3.1. Genome Size Estimation in Nematistius pectoralis
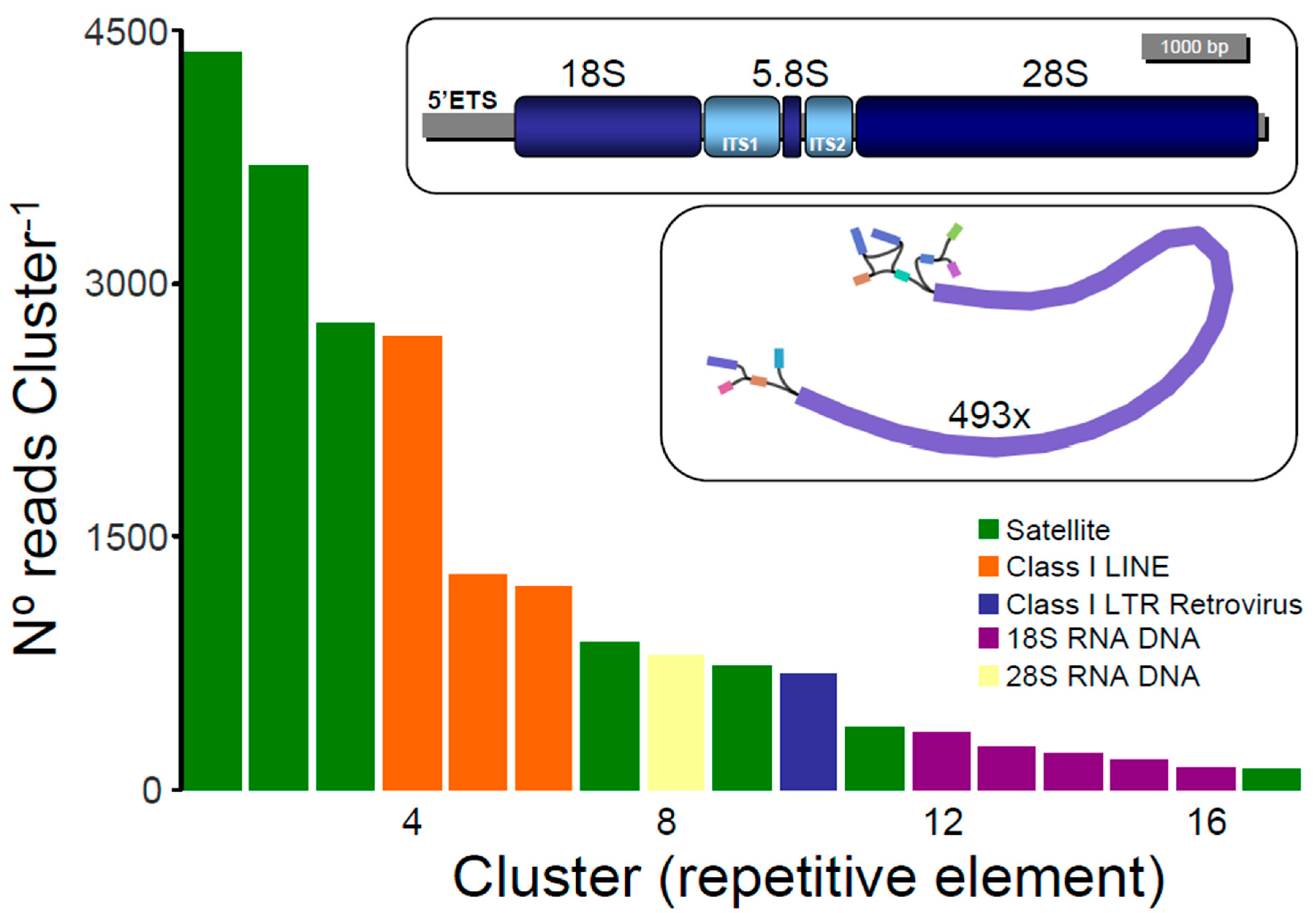
3.2. Repetitive Elements in the Nuclear Genome of Nematistius pectoralis
3.3. Nuclear Ribosomal Operon in Nematistius pectoralis
3.4. Microsatellite Discovery in Nematistius pectoralis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Linzey, D.W. Vertebrate Biology: Systematics, Taxonomy, Natural History, and Conservation; JHU Press: Baltimore, MD, USA, 2020. [Google Scholar]
- Bone, Q.; Moore, R. Biology of Fishes; Taylor & Francis: Oxfordshire, UK, 2008. [Google Scholar]
- Rodríguez-Romero, J.; Moreno-Sánchez, X.G.; Abitia-Cárdenas, L.A.; Palacios-Salgado, D.S. Trophic spectrum of the juvenile roosterfish, Nematistius pectoralis Gill, 1862 (Perciformes: Nematistiidae), in Almejas Bay, Baja California Sur, Mexico. Rev. Biol. Mar. Oceanog. 2009, 44, 803–807. [Google Scholar] [CrossRef]
- Bestley, S.; Patterson, T.A.; Hindell, M.A.; Gunn, J.S. Predicting feeding success in a migratory predator: Integrating telemetry, environment, and modeling techniques. Ecology 2010, 91, 2373–2384. [Google Scholar] [CrossRef] [PubMed]
- Sepulveda, C.A.; Aalbers, S.A.; Bernal, D. Post-release survival and movements patterns of roosterfish (Nematistius pectoralis) off the Central American coastline. Latin Am. J. Aquat. Res. 2015, 43, 162–175. [Google Scholar] [CrossRef]
- Robertson, D.R.; Allen, G.R. Peces Costeros del Pacífico Oriental Tropical: Sistema de Información en Línea; Versión 2.0; Instituto Smithsonian de Investigaciones Tropicales: Balboa, Panama, 2015. [Google Scholar]
- Villalobos-Rojas, F.; Herrera-Correal, J.; Garita-Alvarado, C.A.; Clarke, T.; Beita-Jiménez, A. Actividades pesqueras dependientes de la ictiofauna en el Pacífico Norte de Costa Rica. Rev. Biol. Trop. 2014, 62, 119–137. [Google Scholar] [CrossRef]
- Girard, M.G.; Davis, M.P.; Smith, W.L. The phylogeny of Carangiform fishes: Morphological and genomic investigations of a new fish clade. Copeia 2020, 108, 265–298. [Google Scholar] [CrossRef]
- Miller, D.J.; Lea, R. Guide to the Coastal Marine Fishes of California; Department of Fish and Game, State of California, The Resources Agency: California, CA, USA, 1972; pp. 1–235. [Google Scholar]
- Ortega-Garcia, S.; Sepulveda, C.; Aalbers, S.; Jakes-Cota, U.; Rodriguez-Sanchez, R. Age, growth, and length-weight relationship of roosterfish (Nematistius pectoralis) in the eastern Pacific Ocean. Fish. Bull. 2017, 115, 117–124. [Google Scholar] [CrossRef]
- Molina-Quirós, J.; Hernández-Muñoz, S. Isolation and characterization of 16 novel microsatellite loci in the roosterfish Nematistius pectoralis Gill, 1862 by Illumina sequencing. J. Appl. Ichthyol. 2020, 36, 737–739. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Marçais, G.; Kingsford, C. A fast, lock-free approach for efficient parallel counting of occurrences of k-mers. Bioinformatics 2011, 27, 764–770. [Google Scholar] [CrossRef] [Green Version]
- Sarmashghi, S.; Balaban, M.; Rachtman, E.; Touri, B.; Mirarab, S.; Bafna, V. Estimating repeat spectra and genome length from low-coverage genome skims with RESPECT. bioRxiv 2021. [Google Scholar] [CrossRef]
- Baeza, J.A. A first genomic portrait of the Florida stone crab Menippe mercenaria: Genome size, mitochondrial chromosome, and repetitive elements. Mar. Genom. 2021, 57, 100821. [Google Scholar] [CrossRef] [PubMed]
- Novak, P.; Neumann, P.; Pech, J.J.; Steinhais, L.; Macas, J. RepeatExplorer: A galaxy-based web server for genome-wide characterization of eukaryotic repetitive elements from next generation sequence reads. Bioinformatics 2013, 29, 792–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novak, P.; Neumann, P.; Macas, J. Global analysis of repetitive DNA from unassembled sequence reads using RepeatExplorer2. Nat. Protoc. 2020, 15, 3745–3776. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Madan, A. CAP3: A DNA sequence assembly program. Genome Res. 1999, 9, 868–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Pevzner, P.A. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wick, R.R.; Schultz, M.B.; Zobel, J.; Holt, K.E. Bandage: Interactive visualization of de novo genome assemblies. Bioinformatics 2015, 31, 3350–3352. [Google Scholar] [CrossRef] [Green Version]
- Richard, G.F.; Kerrest, A.; Dujon, B. Comparative genomics and molecular dynamics of DNA repeats in eukaryotes. Microbiol. Mol. Biol. Rev. 2008, 72, 686–727. [Google Scholar] [CrossRef] [Green Version]
- Hubley, R.; Finn, R.D.; Clements, J.; Eddy, S.R.; Jones, T.A.; Bao, W.; Smit, A.F.; Wheeler, T.J. The Dfam database of repetitive DNA families. Nucleic Acids Res. 2016, 44, D81–D89. [Google Scholar] [CrossRef] [Green Version]
- Kalvari, I.; Argasinska, J.; Quinones-Olvera, N.; Nawrocki, E.P.; Rivas, E.; Eddy, S.R.; Bateman, A.; Finn, R.D.; Petrov, A.I. Rfam 13. 0: Shifting to a genome-centric resource for non-coding RNA families. Nucleic Acids Res. 2018, 46, D335–D342. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Lagesen, K.; Hallin, P.F.; Rødland, E.; Stærfeldt, H.H.; Rognes, T.; Ussery, D.W. RNammer: Consistent annotation of rRNA genes in genomic sequences. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson-Palme, J.; Ryberg, M.; Hartmann, M.; Branco, S.; Wang, Z.; Godhe, A.; De Wit, P.; Sanchez-Garcia, M.; Ebersberger, I.; De Sousa, F.; et al. Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data. Methods Ecol. Evol. 2013, 4, 914–919. [Google Scholar] [CrossRef]
- Griffiths, S.M.; Fox, G.; Briggs, P.J.; Donaldson, I.J.; Hood, S.; Richardson, P.; Leaver, G.W.; Truelove, N.K.; Preziosi, R.F. A galaxy-based bioinformatics pipeline for optimized, streamlined microsatellite development from Illumina next-generation sequencing data. Conserv. Genet. Res. 2016, 8, 481–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.; Liu, S.; Zhou, T.; Tian, C.; Bao, L.; Dunham, R.; Liu, Z. Comparative genome analysis of 52 fish species suggests differential associations of repetitive elements with their living aquatic environments. BMC Genom. 2018, 19, 1–10. [Google Scholar] [CrossRef]
- Gregory, T.R. Animal Genome Size Database 2021. Available online: http://www.genomesize.com (accessed on 5 April 2021).
- Zhang, D.C.; Guo, L.; Guo, H.Y.; Zhu, K.C.; Li, S.Q.; Zhang, Y.; Zhang, N.; Liu, B.S.; Jiang, S.G.; Li, J.T. Chromosome-level genome assembly of golden pompano (Trachinotus ovatus) in the family Carangidae. Sci. Data 2019, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Long, E.O.; Dawid, I.B. Repeated genes in eukaryotes. Annu. Rev. Biochem. 1980, 49, 727–764. [Google Scholar] [CrossRef]
- Syaifudin, M.; Bekaert, M.; Taggart, J.B.; Bartie, K.L.; Wehner, S.; Palaiokostas, C.; Khan, M.G.; Selly, S.L.; Hulata, G.; D’Cotta, H.; et al. Species-specific marker discovery in Tilapia. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Faircloth, B.C.; Branstetter, M.G.; White, N.D.; Brady, S.G. Target enrichment of ultraconserved elements from arthropods provides a genomic perspective on relationships among Hymenoptera. Mol. Ecol. Res. 2015, 15, 489–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemmon, A.R.; Emme, S.A.; Lemmon, E.M. Anchored hybrid enrichment for massively high-throughput phylogenomics. Syst. Biol. 2012, 61, 727–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Item | Description |
|---|---|
| Submitted_to_insdc | Yes (SRA) |
| Investigation_type | Eukaryote |
| Project_name | Nematistius pectoralis WGS |
| Geo_loc_name | Paquera, Costa Rica |
| Lat_lon | 9.490091° N, 84.51915° W |
| Depth | 10 m |
| Alt_elev | 0 m |
| Collection_date | 2016-10-06 |
| Collected_by | José Luis Molina-Quirós |
| Env_biome | Seawater (ENVO:00002149) |
| Env_feature | Bay (ENVO:00000032) |
| Env_material | Seawater (ENVO:00002149) |
| Env_package | Water |
| Temp | NA |
| Salinity | NA |
| Sequencing method | Illumina HiSeq2000 |
| Assembly method | Shovill v.1.0.0 |
| Motifs(Bases) | Forward Primer Name | Forward Primer Sequence | Reverse Primer Name | Reverse Primer Sequence |
|---|---|---|---|---|
| AAGAG(50) | rooster_F1 | TGACCAATCGTCTCGTCTCG | rooster_R1 | ACTGCTGGGCAGCTTTTAGC |
| AAGAG(40) | rooster_F2 | TGTGTATTTTATTTTCCAATACATGTAGGC | rooster_R2 | CTGTGTGTGTCCCTTGCTCG |
| AAAAG(50) | rooster_F3 | CAGACAGTGTTGGGTACACCG | rooster_R3 | CCTGTGCTGTTTCTTGCTGC |
| TGC(18) | rooster_F4 | AGGTGAGAGTCGCTGCTGG | rooster_R4 | TCAAACCTCCTCAGCATCCC |
| TTG(15) | rooster_F5 | CATGAAGCTGGTTAACTGTGCC | rooster_R5 | ACAGACAACGGCAACTGTGG |
| TGG(21) | rooster_F6 | ACACAGGGCTCTGACAAGGG | rooster_R6 | CCATACAAGCGATGTCTGGC |
| TGC(15) | rooster_F7 | ACAACCTTTCCCCTCAGTGC | rooster_R7 | GAGCTGGCAGGATCTGTGG |
| AGG(18) | rooster_F8 | ATAGTTGCCCGCCAAACG | rooster_R8 | CGGCTTCAGCTTCCTACTCC |
| ATT(21) | rooster_F9 | TACATGGGACACATCACCCG | rooster_R9 | TTGTGTTGGGATTCAGTGCC |
| TGG(15) | rooster_F10 | TTAAAGCAACGCTGCTGACG | rooster_R10 | ACCGAATAGGTTGTTGTTGGG |
| AAG(15) | rooster_F11 | TCGGTGCTGGTTACCATTAGG | rooster_R11 | CAAACGTTCCACCCAGAAGG |
| AAC(18) | rooster_F12 | AGGATGGGGATTCCTTCACC | rooster_R12 | GGGCAATCTCTTAAGCTGCC |
| AC(18) | rooster_F13 | TTCTGGAGTTTACTGGGGTTCC | rooster_R13 | AGGTGACCTGGAAGCAAAGG |
| AG(16) | rooster_F14 | AGACCAGGCTGTCTCTCTCTCC | rooster_R14 | GCTAATTGAAATGCCGCTGG |
| AG(12) | rooster_F15 | AGTGAGTTTGCGTGATTGGG | rooster_R15 | CATGGAAACCTTGCCAGAGC |
| TG(12) | rooster_F16 | CCCTCCAGGGAATTTGTACG | rooster_R16 | CTGACAGCTAGCCCAGGTCC |
| TG(40) | rooster_F17 | CATGTATATGCCATTTTATGTCTGTCC | rooster_R17 | TCGGTGGTTGTTGTCTTTTCC |
| TG(22) | rooster_F18 | CAGTCTAGCACCATTCTGGGG | rooster_R18 | TTCTGCTACTTGCTGAGCCG |
| AC(12) | rooster_F19 | GGCAGCTGGAGTGAAAGTAAGG | rooster_R19 | TGCAGAAAGTAGTGTGGACTTGG |
| TG(14) | rooster_F20 | CTTCGAGGAGGCCTGTTACC | rooster_R20 | TGGCCTAAATACAGGCTTGG |
| AT(14) | rooster_F21 | GTGCTGGTTTAAAGGCAACG | rooster_R21 | GCAGCTCATCGAAAGAATGC |
| AC(20) | rooster_F22 | TAGCGATGGCACTTTCATGG | rooster_R22 | GGCAGAGATCATAATTGCTGTGG |
| AC(20) | rooster_F23 | TGTGCGTCTCTTGTGTCTGG | rooster_R23 | AGATTAAGAGAGCGTGTGAGCG |
| AG(12) | rooster_F24 | CCATCTCTCGCCAATTCTCC | rooster_R24 | TGTTGCAATTTGATAGTCTGGC |
| AG(18) | rooster_F25 | AAGATTCACTTTGCTTCAAGGC | rooster_R25 | TAACGAGTATCCAGAGCGGG |
| AG(12) | rooster_F26 | CAGCAGGGTCTGAAGCAAGC | rooster_R26 | CTGCCCTTCCTGCTGTTACC |
| TG(24) | rooster_F27 | CTCATGGGAAGAGACAAGTAGTCC | rooster_R27 | GCCTCCTGTTGTAAGCCTGC |
| AC(12) | rooster_F28 | TTAAACCATCCTTGAGTGTGTGG | rooster_R28 | TCCCAAAGCAGATACCCACC |
| TG(24) | rooster_F29 | TGGGCATATTTTGGTTAACGG | rooster_R29 | AGTGGTTGTCCTCATCACCG |
| AC(18) | rooster_F30 | CGAAAAGGTCCTTGACGAGC | rooster_R30 | ACATGTCGCAAAGGAGAGGG |
| TG(50) | rooster_F31 | GTTGCATGGCAGCTCTATCG | rooster_R31 | AACCCACCCCAGCAAAGC |
| TG(16) | rooster_F32 | GAAAACACGAGGGCAGTACG | rooster_R32 | CCACAGCAGAAACACAATGG |
| AT(12) | rooster_F33 | TTTGATACAGGATTTAGGTGCCC | rooster_R33 | GGAGAGGAGCGTAGGAATGG |
| AC(54) | rooster_F34 | TCGAAATAAGGGAGAGAGCAGC | rooster_R34 | GGAACAGCTTTGGAGGATGG |
| AC(20) | rooster_F35 | CACAATGCATTAGGACCTCCG | rooster_R35 | AGAAGGAGAGATAGCCCCGC |
| TG(18) | rooster_F36 | AGACACCAGCACACACGTCC | rooster_R36 | GTTGTCCAAACACCAGCAGC |
| TG(12) | rooster_F37 | GAAATCAATAGCGGATTCGACC | rooster_R37 | ACCATGATATTTCTGCCGGG |
| TG(12) | rooster_F38 | GATAAATGCGCCACACTTGG | rooster_R38 | GCATGTAAGAGCAGGGTTGC |
| AC(36) | rooster_F39 | CTTTGAGTCTTACTTTTATAATGTGCTCC | rooster_R39 | CTTTGGAAAAGGGACAACCG |
| TG(12) | rooster_F40 | CGGCAGAAATGTGTTTGAGC | rooster_R40 | CCACATAGCCTTCATTTCACTCC |
| TG(14) | rooster_F41 | ACACCAACCCACCCACTAGC | rooster_R41 | TGAATGCGTGGATGGTATCG |
| AG(12) | rooster_F42 | ACAGAGCAGCCTGTATGGGG | rooster_R42 | CTGAGCCAGAGAAAGGAGGG |
| AG(12) | rooster_F43 | CCCAGATCCTTTCATCCAGC | rooster_R43 | AATCTCACCGATGCGTTTCC |
| TG(32) | rooster_F44 | ATGATGATGAACGCAGAGGG | rooster_R44 | GAGCCACTAGCCAGTCCTGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baeza, J.A.; Molina-Quirós, J.L.; Hernández-Muñoz, S. Genome Survey Sequencing of an Iconic ‘Trophy’ Sportfish, the Roosterfish Nematistius pectoralis: Genome Size, Repetitive Elements, Nuclear RNA Gene Operon, and Microsatellite Discovery. Genes 2021, 12, 1710. https://doi.org/10.3390/genes12111710
Baeza JA, Molina-Quirós JL, Hernández-Muñoz S. Genome Survey Sequencing of an Iconic ‘Trophy’ Sportfish, the Roosterfish Nematistius pectoralis: Genome Size, Repetitive Elements, Nuclear RNA Gene Operon, and Microsatellite Discovery. Genes. 2021; 12(11):1710. https://doi.org/10.3390/genes12111710
Chicago/Turabian StyleBaeza, J. Antonio, José Luis Molina-Quirós, and Sebastián Hernández-Muñoz. 2021. "Genome Survey Sequencing of an Iconic ‘Trophy’ Sportfish, the Roosterfish Nematistius pectoralis: Genome Size, Repetitive Elements, Nuclear RNA Gene Operon, and Microsatellite Discovery" Genes 12, no. 11: 1710. https://doi.org/10.3390/genes12111710