
Association of PIP4K2A Polymorphisms with Alcohol Use Disorder

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Genetic Analysis
2.3. Statistical Analysis
3. Results
3.1. PIP4K2A rs746203
3.2. PIP4K2A rs2230469 (rs10828317)
4. Discussion
Limitations and Strengths of Our Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Status Report on Alcohol and Health 2018; WHO: Geneva, Switzerland, 2018; 472p, Available online: https://apps.who.int/iris/bitstream/handle/10665/274603/9789241565639-eng.pdf (accessed on 20 July 2020).
- Verhulst, B.; Neale, M.C.; Kendler, K.S. The heritability of alcohol use disorders: A meta-analysis of twin and adoption studies. Psychol. Med. 2015, 45, 1061–1072. [Google Scholar] [CrossRef] [Green Version]
- Reilly, M.T.; Noronha, A.; Goldman, D.; Koob, G.F. Genetic studies of alcohol dependence in the context of the addiction cycle. Neuropharmacology 2017, 122, 3–21. [Google Scholar] [CrossRef]
- Hart, A.B.; Kranzler, H.R. Alcohol Dependence Genetics: Lessons Learned From Genome-Wide Association Studies (GWAS) and Post-GWAS Analyses. Alcohol. Clin. Exp. Res. 2015, 39, 1312–1327. [Google Scholar] [CrossRef] [Green Version]
- Bhaskar, L.V.K.S.; Kumar, S.A. Polymorphisms in genes encoding dopamine signalling pathway and risk of alcohol dependence: A systematic review. Acta Neuropsychiatr. 2013, 26, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Forero, D.A.; López-León, S.; Shin, H.D.; Park, B.L.; Kim, D.-J. Meta-analysis of six genes (BDNF, DRD1, DRD3, DRD4, GRIN2B and MAOA) involved in neuroplasticity and the risk for alcohol dependence. Drug Alcohol Depend. 2015, 149, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Kranzler, H.R.; Zhou, H.; Kember, R.L.; Smith, R.V.; Justice, A.C.; Damrauer, S.; Tsao, P.S.; Klarin, D.; Baras, A.; Reid, J.; et al. Genome-wide association study of alcohol consumption and use disorder in 274,424 individuals from multiple populations. Nat. Commun. 2019, 10, 1499. [Google Scholar] [CrossRef] [Green Version]
- Thompson, A.; Cook, J.; Choquet, H.; Jorgenson, E.; Yin, J.; Kinnunen, T.; Barclay, J.; Morris, A.P.; Pirmohamed, M. Functional validity, role, and implications of heavy alcohol consumption genetic loci. Sci. Adv. 2020, 6, eaay5034. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Fan, R.; Li, M.D. Meta-Analysis Reveals Significant Association of the 3′-UTR VNTR in SLC6A3 with Alcohol Dependence. Alcohol. Clin. Exp. Res. 2016, 40, 1443–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koob, G.F.; Le Moal, M. Drug Abuse: Hedonic Homeostatic Dysregulation. Science 1997, 278, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Batalla, A.; Homberg, J.R.; Lipina, T.V.; Sescousse, G.; Luijten, M.; Ivanova, S.A.; Schellekens, A.F.; Loonen, A.J. The role of the habenula in the transition from reward to misery in substance use and mood disorders. Neurosci. Biobehav. Rev. 2017, 80, 276–285. [Google Scholar] [CrossRef] [Green Version]
- Loonen, A.J.; Ivanova, S.A. Circuits regulating pleasure and happiness: Evolution and role in mental disorders. Acta Neuropsychiatr. 2018, 30, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Loonen, A.J.M.; Ivanova, S.A. Evolution of circuits regulating pleasure and happiness with the habenula in control. CNS Spectrums 2019, 24, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Clarke, J.H.; Irvine, R.F. Enzyme activity of the PIP4K2A gene product polymorphism that is implicated in schizophrenia. Psychopharmacology 2013, 230, 329–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiselton, D.L.; Maher, B.S.; Webb, B.T.; Bigdeli, T.B.; O’Neill, F.A.; Walsh, D.; Kendler, K.S.; Riley, B.P. Association analysis of the PIP4K2A gene on chromosome 10p12 and schizophrenia in the Irish study of high density schizophrenia families (ISHDSF) and the Irish case-control study of schizophrenia (ICCSS). Am. J. Med. Genet. B Neuropsychiatr. Genet. 2010, 153, 323–331. [Google Scholar] [CrossRef] [Green Version]
- Fedorenko, O.I.; Rudikov, E.V.; Gavrilova, V.A.; Boiarko, E.G.; Semke, A.V.; Ivanova, S. Association of (N251S)-PIP5K2A with schizophrenic disorders: A study of the Russian population of Siberia. Zhurnal Nevrol. Psikhiatrii Im. SS Korsakova 2013, 113, 58–61. [Google Scholar]
- Fedorenko, O.Y.; Loonen, A.J.M.; Lang, F.; Toshchakova, V.A.; Boyarko, E.G.; Semke, A.V.; Bokhan, N.; Govorin, N.V.; Aftanas, L.I.; Ivanova, S. Association Study Indicates a Protective Role of Phosphatidylinositol-4-Phosphate-5-Kinase against Tardive Dyskinesia. Int. J. Neuropsychopharmacol. 2015, 18, pyu098. [Google Scholar] [CrossRef] [Green Version]
- Fedorenko, O.; Strutz-Seebohm, N.; Henrion, U.; Ureche, O.N.; Lang, F.; Seebohm, G.; Lang, U.E. A schizophrenia-linked mutation in PIP5K2A fails to activate neuronal M channels. Psychopharmacology 2008, 199, 47–54. [Google Scholar] [CrossRef]
- Fedorenko, O.; Tang, C.; Sopjani, M.; Foeller, M.; Gehring, E.-M.; Strutz-Seebohm, N.; Ureche, O.N.; Ivanova, S.; Semke, A.V.; Lang, F.; et al. PIP5K2A-dependent regulation of excitatory amino acid transporter EAAT3. Psychopharmacology 2009, 206, 429–435. [Google Scholar] [CrossRef]
- Seebohm, G.; Wrobel, E.; Pusch, M.; Dicks, M.; Terhag, J.; Matschke, V.; Rothenberg, I.; Ursu, O.N.; Hertel, F.; Pott, L.; et al. Structural basis of PI(4,5)P2-dependent regulation of GluA1 by phosphatidylinositol-5-phosphate 4-kinase, type II, alpha (PIP5K2A). Pflügers Archiv-Eur. J. Physiol. 2014, 466, 1885–1897. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. International Statistical Classification of Diseases and Health Related Problems ICD-10; World Health Organization: Geneva, Switzerland, 2010; 201p, Available online: https://www.who.int/classifications/icd/ICD10Volume2_en_2010.pdf (accessed on 9 September 2021).
- Kegeles, L.S.; Horga, G.; Ghazzaoui, R.; Rosengard, R.; Ojeil, N.; Xu, X.; Slifstein, M.; Petrakis, I.; O’Malley, S.S.; Krystal, J.H.; et al. Enhanced Striatal Dopamine Release to Expectation of Alcohol: A Potential Risk Factor for Alcohol Use Disorder. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2018, 3, 591–598. [Google Scholar] [CrossRef]
- Stephenson-Jones, M.; Kardamakis, A.; Robertson, B.; Grillner, S. Independent circuits in the basal ganglia for the evaluation and selection of actions. Proc. Natl. Acad. Sci. USA 2013, 110, E3670–E3679. [Google Scholar] [CrossRef] [Green Version]
- Stephenson-Jones, M.; Yu, K.; Ahrens, S.; Tucciarone, J.M.; Van Huijstee, A.N.; Mejia, L.A.; Penzo, M.A.; Tai, L.-H.; Wilbrecht, L.-H.T.L.; Li, B. A basal ganglia circuit for evaluating action outcomes. Nat. Cell Biol. 2016, 539, 289–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephenson-Jones, M.; Bravo-Rivera, C.; Ahrens, S.; Furlan, A.; Xiao, X.; Fernandes-Henriques, C.; Li, B. Opposing Contributions of GABAergic and Glutamatergic Ventral Pallidal Neurons to Motivational Behaviors. Neuron 2020, 105, 921–933.e5. [Google Scholar] [CrossRef]
- Shah, A.; Zuo, W.; Kang, S.; Li, J.; Fu, R.; Zhang, H.; Bekker, A.; Ye, J.-H. The lateral habenula and alcohol: Role of glutamate and M-type potassium channels. Pharmacol. Biochem. Behav. 2017, 162, 94–102. [Google Scholar] [CrossRef]
- Nieuwenhuys, R. The Chemoarchitecture of the Brain; Springer: Berlin, Germany, 1985. [Google Scholar]
- Loonen, A.J.; Wilffert, B.; Ivanova, S. Putative role of pharmacogenetics to elucidate the mechanism of tardive dyskinesia in schizophrenia. Pharmacogenomics 2019, 20, 1199–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, A.; Homma, D.; Gibb, L.G.; Amemori, K.-I.; Rubin, S.J.; Hood, A.S.; Riad, M.H.; Graybiel, A.M. A Corticostriatal Path Targeting Striosomes Controls Decision-Making under Conflict. Cell 2015, 161, 1320–1333. [Google Scholar] [CrossRef] [Green Version]
- Camchong, J.; Endres, M.; Fein, G. Decision making, risky behavior, and alcoholism. Frontal Lobes 2014, 125, 227–236. [Google Scholar] [CrossRef]
- Kovács, I.; Richman, M.J.; Janka, Z.; Maraz, A.; Andó, B. Decision making measured by the Iowa Gambling Task in alcohol use disorder and gambling disorder: A systematic review and meta-analysis. Drug Alcohol Depend. 2017, 181, 152–161. [Google Scholar] [CrossRef]
- Coker, A.R.; Weinstein, D.N.; Vega, T.A.; Miller, C.S.; Kayser, A.S.; Mitchell, J.M. The catechol-O-methyltransferase inhibitor tolcapone modulates alcohol consumption and impulsive choice in alcohol use disorder. Psychopharmacology 2020, 237, 3139–3148. [Google Scholar] [CrossRef] [PubMed]
- McHugh, P.C.; Buckley, D.A. The Structure and Function of the Dopamine Transporter and its Role in CNS Diseases. Vitam. Horm. 2015, 98, 339–369. [Google Scholar] [CrossRef] [PubMed]


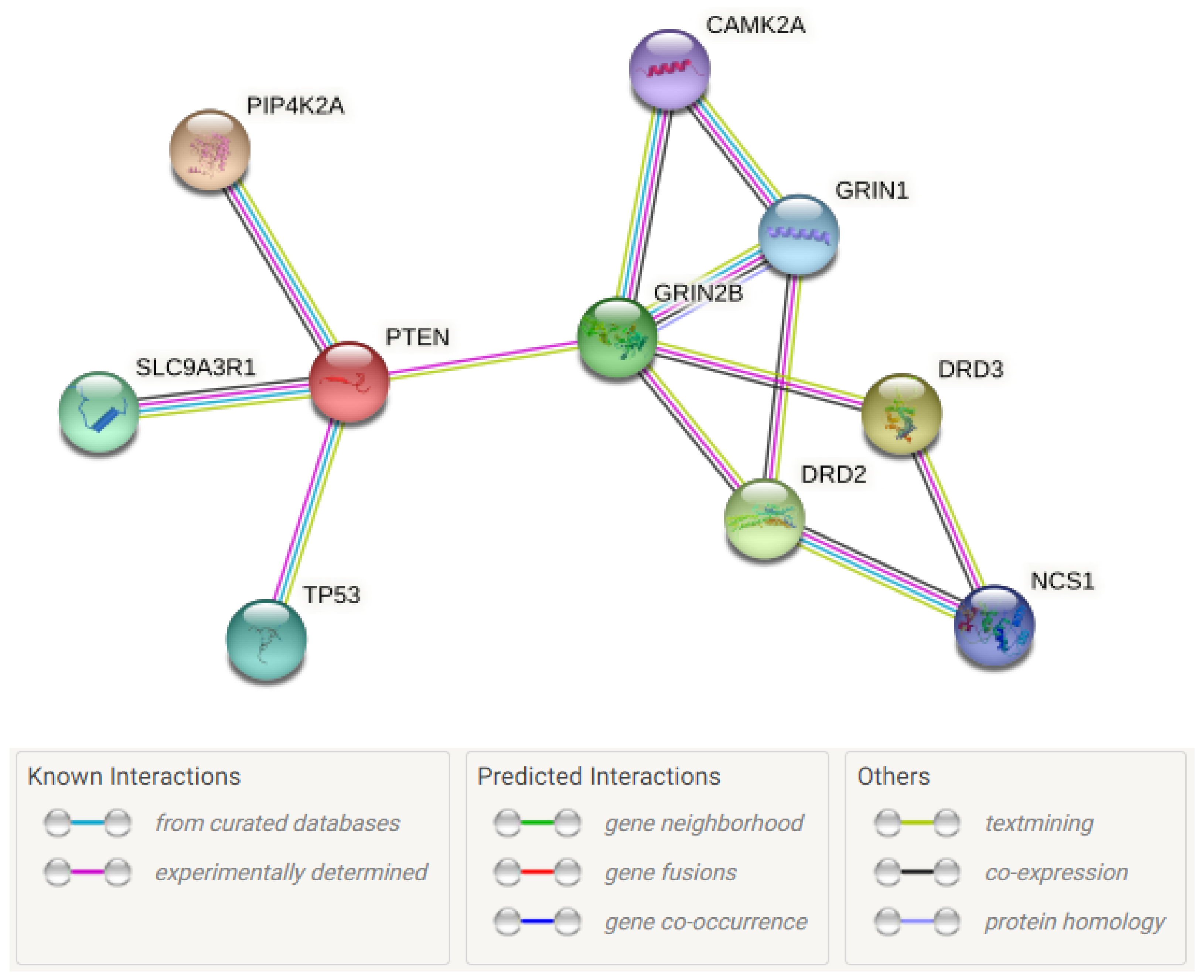{kind=link}
| Genotypes/ Alleles | Patients with AUD (n = 253) | Controls (n = 220) | x2, p | OR | 95% CI | |
|---|---|---|---|---|---|---|
| CC | 34 (13.44%) | 39 (17.72%) | 5.31 0.07 | 0.72 | 0.44 | 1.19 |
| CT | 106 (41.90%) | 105 (47.72%) | 0.79 | 0.55 | 1.14 | |
| TT | 113 (44.67%) | 76 (34.55%) | 1.53 | 1.05 | 2.22 | |
| C | 174 (34.39%) | 183 (41.59%) | 5.20 0.023 * | 0.74 | 0.57 | 0.96 |
| T | 332 (65.61%) | 257 (58.41%) | 1.36 | 1.04 | 1.77 | |
| Genotypes/ Alleles | Patients with AUD (n = 250) | Controls (n = 216) | x2, p | OR | 95% CI | |
|---|---|---|---|---|---|---|
| CC | 20 (8.0%) | 33 (15.28%) | 7.27 0.026 * | 0.48 | 0.27 | 0.87 |
| CT | 97 (38.8%) | 87 (40.28%) | 0.94 | 0.65 | 1.36 | |
| TT | 133 (53.2%) | 96 (44.44%) | 1.42 | 1.05 | 2.05 | |
| C | 137 (27.4%) | 153 (35.42%) | 6.95 0.0084 * | 0.69 | 0.52 | 0.91 |
| T | 363 (72.6%) | 279 (64.58%) | 1.45 | 1.10 | 1.92 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fedorenko, O.Y.; Mikhalitskaya, E.V.; Toshchakova, V.A.; Loonen, A.J.M.; Bokhan, N.A.; Ivanova, S.A. Association of PIP4K2A Polymorphisms with Alcohol Use Disorder. Genes 2021, 12, 1642. https://doi.org/10.3390/genes12101642
Fedorenko OY, Mikhalitskaya EV, Toshchakova VA, Loonen AJM, Bokhan NA, Ivanova SA. Association of PIP4K2A Polymorphisms with Alcohol Use Disorder. Genes. 2021; 12(10):1642. https://doi.org/10.3390/genes12101642
Chicago/Turabian StyleFedorenko, Olga Yu., Ekaterina V. Mikhalitskaya, Valentina A. Toshchakova, Anton J. M. Loonen, Nikolay A. Bokhan, and Svetlana A. Ivanova. 2021. "Association of PIP4K2A Polymorphisms with Alcohol Use Disorder" Genes 12, no. 10: 1642. https://doi.org/10.3390/genes12101642