Eco-Evolutionary Feedbacks and the Maintenance of Metacommunity Diversity in a Changing Environment

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Overview
2.2. Within-Patch Ecology
2.3. Evolution
2.4. Extinction
2.5. Dispersal
2.6. Environmental Dynamics
2.7. Metacommunity Initialization
2.8. Simulation Conditions
2.9. Metrics for Evolutionary and Ecological Properties
3. Results
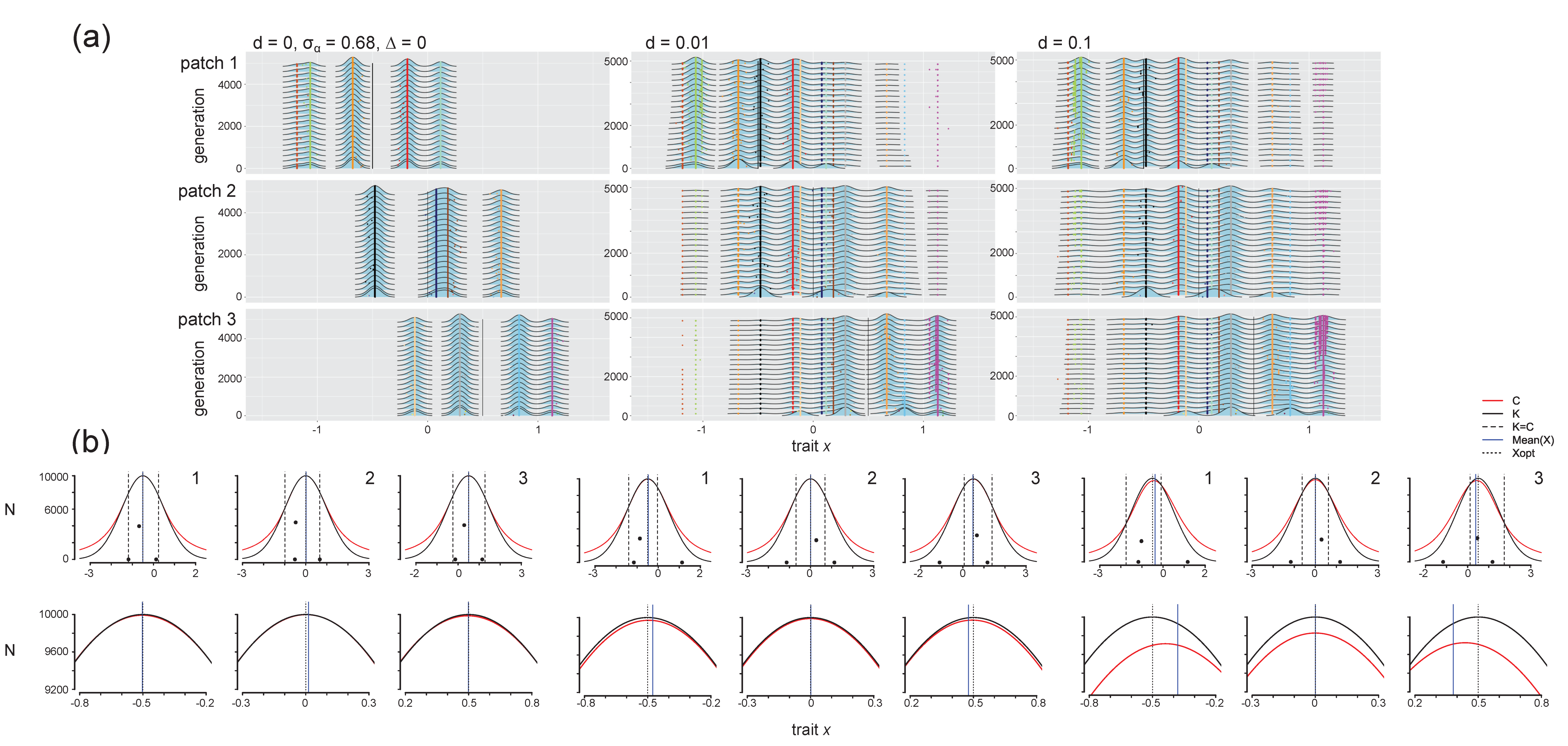
3.1. Initial Communities
3.2. Effect of Dispersal in the Absence of Environmental Change
3.2.1. Selection Pressures from Carrying Capacity and Competition
3.2.2. Phenotypic Trajectories and Metacommunity Dynamics
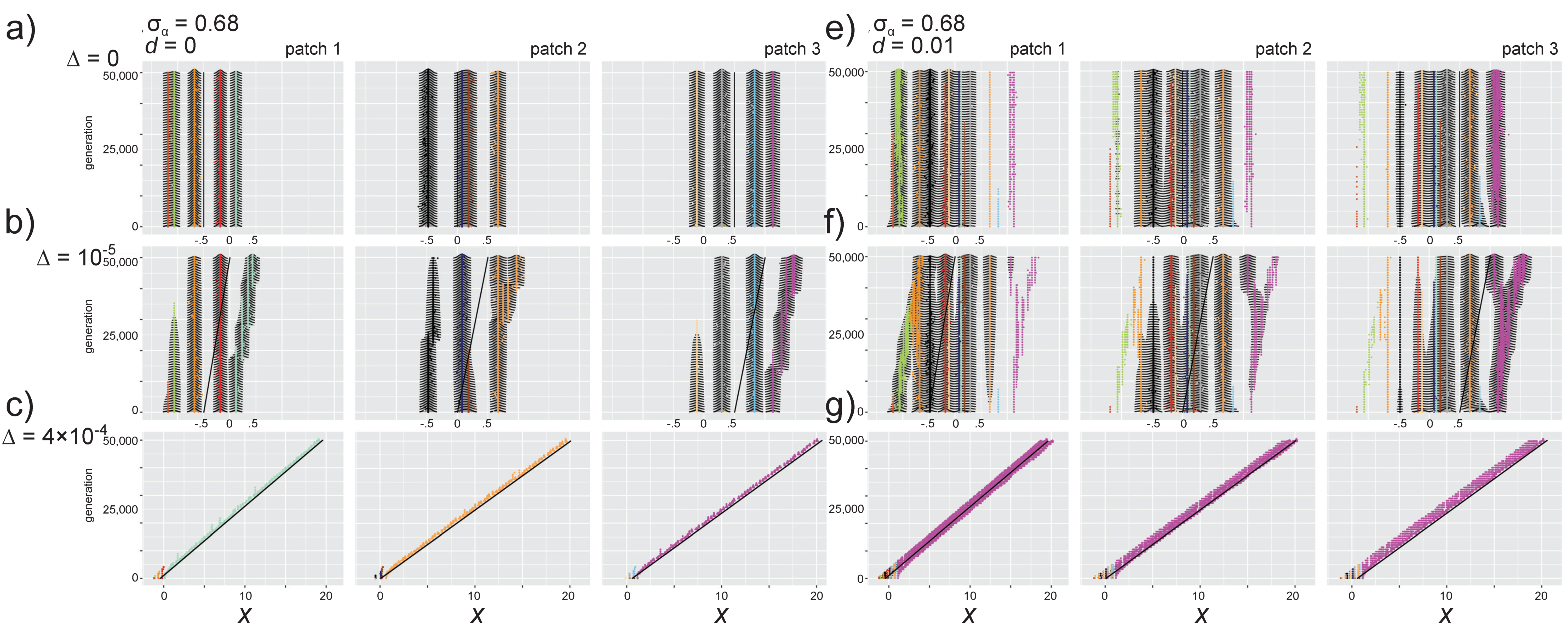
3.3. Effects of Directional Environmental Change in the Absence of Dispersal
3.3.1. Selection Pressures from Carrying Capacity and Competition
3.3.2. Phenotypic Trajectories and Metacommunity Dynamics
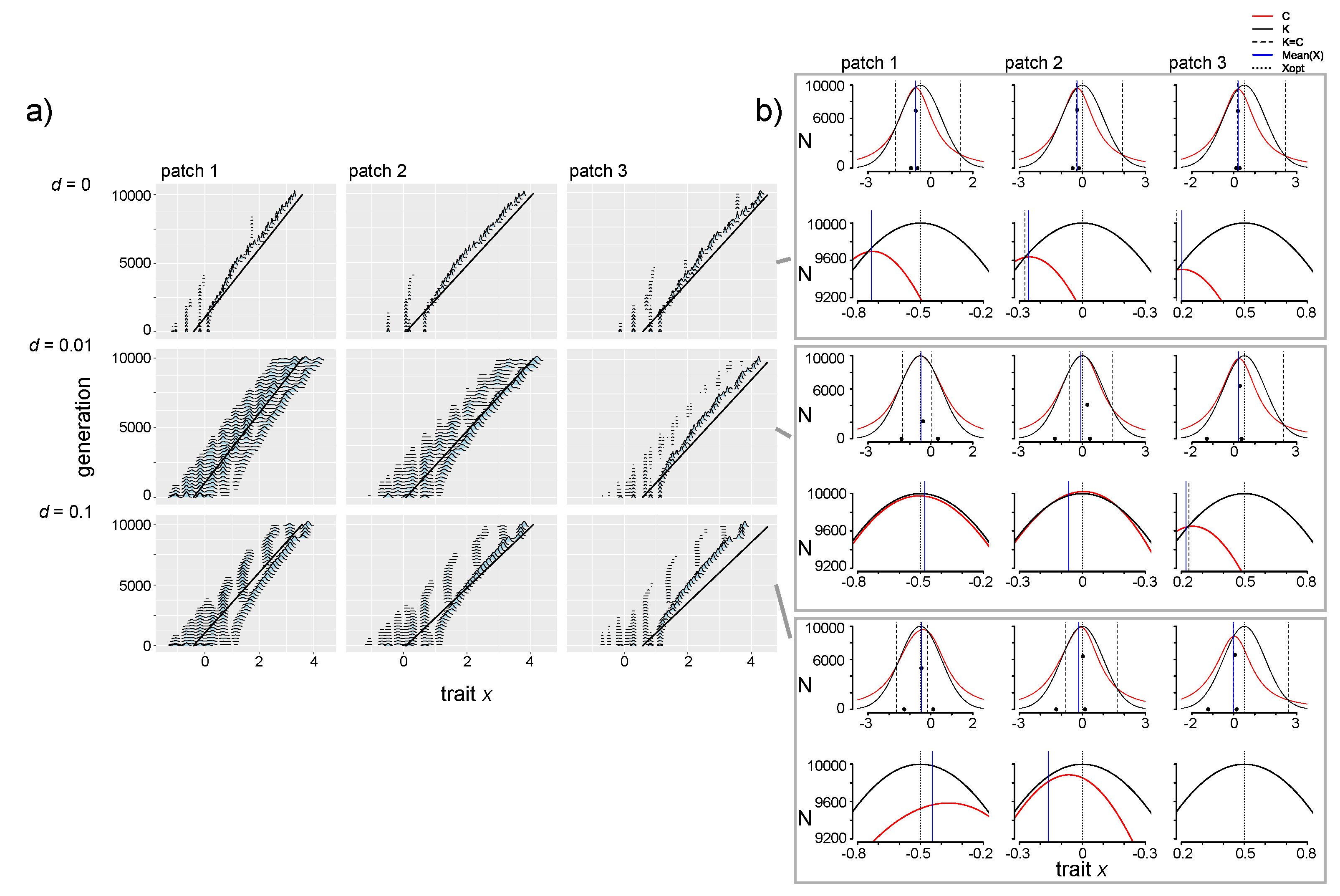
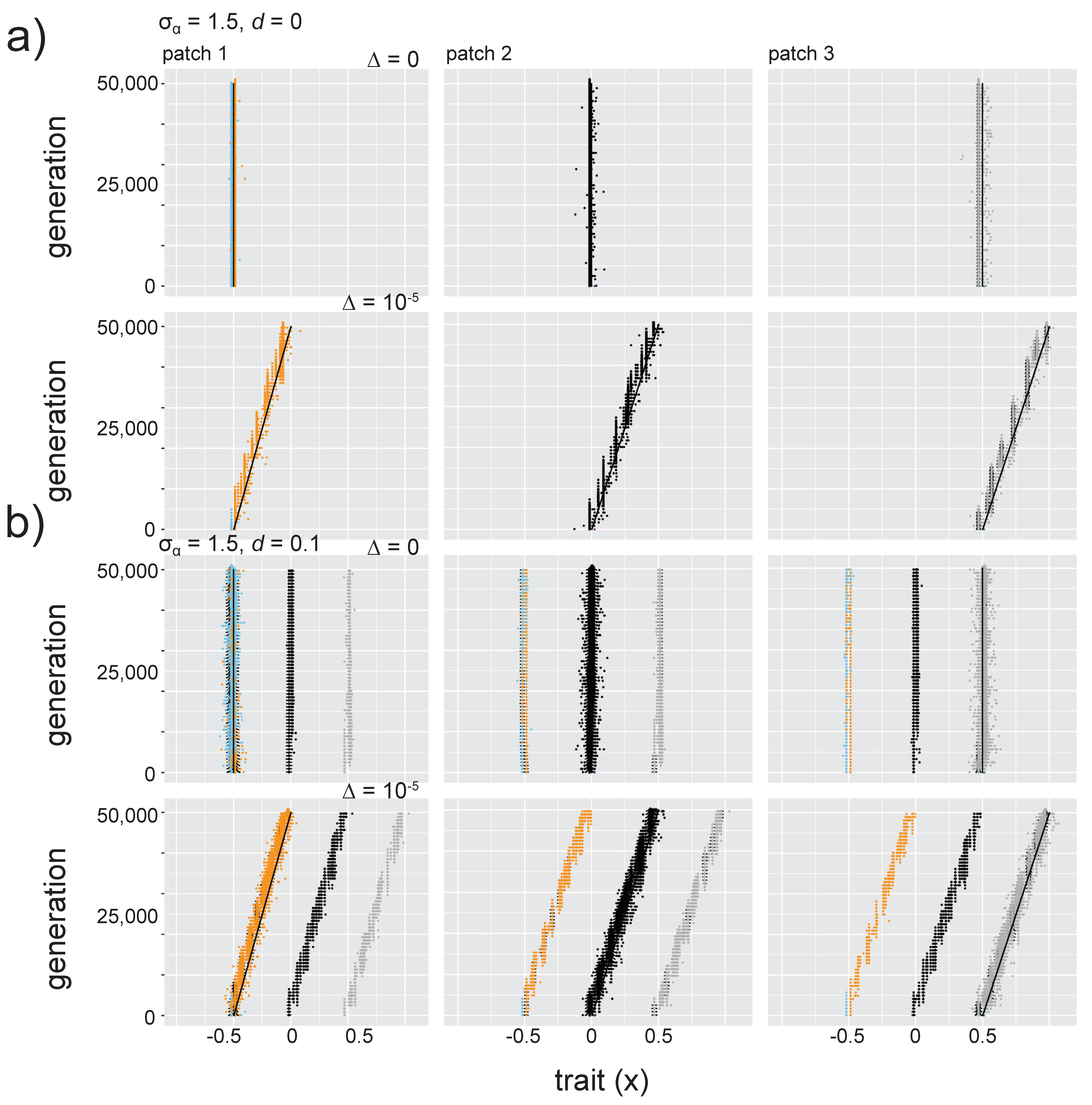
3.4. Effects of Dispersal and Directional Environmental Change
3.4.1. Selection Pressures from Carrying Capacity and Competition
3.4.2. Phenotypic Trajectories and Metacommunity Dynamics
3.4.3. Increasing Niche Width
4. Discussion
4.1. Overview
4.2. Dynamic Variation in Selection Pressures over Time and Space
4.3. Metacommunity Patterns for Trait and Species Diversity
4.4. Eco-Evolutionary Feedback Loops and Adaptive Dynamics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tilman, D. Competition and biodiversity in spatially structured habitats. Ecology 1994, 75, 2–16. [Google Scholar] [CrossRef]
- Hautier, Y.; Niklaus, P.A.; Hector, A. Competition for light causes plant biodiversity loss after eutrophication. Science 2009, 324, 636–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chesson, P. Mechanisms of maintenance of species diversity. Annu. Rev. Ecol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef] [Green Version]
- Hart, S.P.; Turcotte, M.M.; Levine, J.M. Effects of rapid evolution on species coexistence. Proc. Natl. Acad. Sci. USA 2019, 116, 2112–2117. [Google Scholar] [CrossRef] [Green Version]
- Bernhardt, J.R.; Kratina, P.; Pereira, A.L.; Tamminen, M.; Thomas, M.K.; Narwani, A. The evolution of competitive ability for essential resources. Philos. Trans. R. Soc. B 2020, 375, 20190247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillespie, R. Community assembly through adaptive radiation in Hawaiian spiders. Science 2004, 303, 356–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Losos, J.B.; Ricklefs, R.E. Adaptation and diversification on islands. Nature 2009, 457, 830–836. [Google Scholar] [CrossRef]
- Drury, J.; Clavel, J.; Manceau, M.; Morlon, H. Estimating the effect of competition on trait evolution using maximum likelihood inference. Syst. Biol. 2016, 65, 700–710. [Google Scholar] [CrossRef] [Green Version]
- Roughgarden, J. Evolution of niche width. Am. Nat. 1972, 106, 683–718. [Google Scholar] [CrossRef]
- Lankau, R.A. Rapid evolutionary change and the coexistence of species. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 335–354. [Google Scholar] [CrossRef]
- Moran, E.V.; Alexander, J.M. Evolutionary responses to global change: Lessons from invasive species. Ecol. Lett. 2014, 17, 637–649. [Google Scholar] [CrossRef] [PubMed]
- De Mazancourt, C.; Johnson, E.; Barraclough, T.G. Biodiversity inhibits species’ evolutionary responses to changing environments. Ecol. Lett. 2008, 11, 380–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chase, J.M.; Leibold, M.A. Ecological Niches: Linking Classical and Contemporary Approaches; University of Chicago Press: Chicago, IL, USA, 2003. [Google Scholar]
- Leibold, M.A.; Holyoak, M.; Mouquet, N.; Amarasekare, P.; Chase, J.M.; Hoopes, M.F.; Holt, R.D.; Shurin, J.B.; Law, R.; Tilman, D.; et al. The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 2004, 7, 601–613. [Google Scholar] [CrossRef]
- Urban, M.C.; Leibold, M.A.; Amarasekare, P.; De Meester, L.; Gomulkiewicz, R.; Hochberg, M.E.; Klausmeier, C.A.; Loeuille, N.; De Mazancourt, C.; Norberg, J.; et al. The evolutionary ecology of metacommunities. Trends Ecol. Evol. 2008, 23, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Loeuille, N.; Leibold, M.A. Evolution in metacommunities: On the relative importance of species sorting and monopolization in structuring communities. Am. Nat. 2008, 171, 788–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urban, M.C.; De Meester, L. Community monopolization: Local adaptation enhances priority effects in an evolving metacommunity. Proc. R. Soc. B Biol. Sci. 2009, 276, 4129–4138. [Google Scholar] [CrossRef]
- Vanoverbeke, J.; Urban, M.C.; De Meester, L. Community assembly is a race between immigration and adaptation: Eco-evolutionary interactions across spatial scales. Ecography 2016, 39, 858–870. [Google Scholar] [CrossRef]
- Norberg, J.; Urban, M.C.; Vellend, M.; Klausmeier, C.A.; Loeuille, N. Eco-evolutionary responses of biodiversity to climate change. Nat. Clim. Chang. 2012, 2, 747–751. [Google Scholar] [CrossRef]
- McKee, D.; Ebert, D. The effect of temperature on maturation threshold body-length in Daphnia magna. Oecologia 1996, 108, 627–630. [Google Scholar] [CrossRef]
- Burns, C.W. Relation between filtering rate, temperature, and body size in four species of Daphnia. Limnol. Oceanogr. 1969, 14, 693–700. [Google Scholar] [CrossRef]
- Dodson, S.I. Zooplankton competition and predation: An experimental test of the size-efficiency hypothesis. Ecology 1974, 55, 605–613. [Google Scholar] [CrossRef]
- Hall, D.J.; Threlkeld, S.T.; Burns, C.W.; Crowley, P.H. The size-efficiency hypothesis and the size structure of zooplankton communities. Annu. Rev. Ecol. Syst. 1976, 7, 177–208. [Google Scholar] [CrossRef]
- Terhorst, C.P.; Miller, T.E.; Levitan, D.R. Evolution of prey in ecological time reduces the effect size of predators in experimental microcosms. Ecology 2010, 91, 629–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- TerHorst, C.P. Experimental evolution of protozoan traits in response to interspecific competition. J. Evol. Biol. 2011, 24, 36–46. [Google Scholar] [CrossRef]
- Dieckmann, U.; Doebeli, M. On the origin of species by sympatric speciation. Nature 1999, 400, 354–357. [Google Scholar] [CrossRef] [Green Version]
- Kisdi, É. Evolutionary branching under asymmetric competition. J. Theor. Biol. 1999, 197, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Doebeli, M. Adaptive Diversification. Monographs in Population Biology; Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Aguilée, R.; Claessen, D.; Lambert, A. Adaptive radiation driven by the interplay of eco-evolutionary and landscape dynamics. Evolution 2013, 67, 1291–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rettelbach, A.; Kopp, M.; Dieckmann, U.; Hermisson, J. Three modes of adaptive speciation in spatially structured populations. Am. Nat. 2013, 182, E215–E234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, J. Evolutionary responses to environmental changes: How does competition affect adaptation? Evolution 2008, 62, 421–435. [Google Scholar] [CrossRef] [PubMed]
- Osmond, M.M.; de Mazancourt, C. How competition affects evolutionary rescue. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120085. [Google Scholar] [CrossRef] [Green Version]
- Gomulkiewicz, R.; Holt, R.D. When does evolution by natural selection prevent extinction? Evolution 1995, 49, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Bell, G. Evolutionary rescue. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 605–627. [Google Scholar] [CrossRef]
- Metz JA, J.; Nisbet, R.M.; Geritz SA, H. How should we define ‘fitness’ for general ecological scenarios? Trends Ecol. Evol. 1992, 7, 198–202. [Google Scholar] [CrossRef]
- Dieckmann, U.; Law, R. The dynamical theory of coevolution: A derivation from stochastic ecological processes. J. Math. Biol. 1996, 34, 579–612. [Google Scholar] [CrossRef]
- Barton, N.H.; Polechová, J. The limitations of adaptive dynamics as a model of evolution. J. Evol. Biol. 2005, 18, 1186–1190. [Google Scholar] [CrossRef]
- Goulden, C.E.; Henry, L.L.; Tessier, A.J. Body size, energy reserves, and competitive ability in three species of Cladocera. Ecology 1982, 63, 1780–1789. [Google Scholar] [CrossRef]
- Lamichhaney, S.; Han, F.; Berglund, J.; Wang, C.; Almén, M.S.; Webster, M.T.; Grant, B.R.; Grant, P.R.; Andersson, L. A beak size locus in Darwin’s finches facilitated character displacement during a drought. Science 2016, 352, 470–474. [Google Scholar] [CrossRef]
- Huston, M.A.; DeAngelis, D.L. Competition and coexistence: The effects of resource transport and supply rates. Am. Nat. 1994, 144, 954–977. [Google Scholar] [CrossRef]
- Brännström, Å.; Johansson, J.; von Festenberg, N. The Hitchhiker’s Guide to Adaptive Dynamics. Games 2013, 4, 304–328. [Google Scholar] [CrossRef] [Green Version]
- Doebeli, M.; Dieckmann, U. Evolutionary branching and sympatric speciation caused by different types of ecological interactions. Am. Nat. 2000, 156, S77–S101. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Wilke, C.O. ggridges: Ridgeline Plots in ‘ggplot2’. R Package Version 0.5.1. 2018. Available online: https://CRAN.R-project.org/package=ggridges (accessed on 28 November 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 28 November 2020).
- Geritz SA, H.; Kisdi, E.; Mesze, G.; Metz JA, J. Evolutionarily singular strategies and the adaptive growth and branching of the evolutionary tree. Evol. Ecol. 1998, 12, 35–57. [Google Scholar] [CrossRef]
- Christiansen, F.B.; Loeschcke, V. Evolution and intraspecific exploitative competition I. One-locus theory for small additive gene effects. Theor. Popul. Biol. 1980, 18, 297–313. [Google Scholar] [CrossRef]
- Bürger, R.; Lynch, M. Evolution and extinction in a changing environment: A quantitative-genetic analysis. Evolution 1995, 49, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Kopp, M.; Matuszewski, S. Rapid evolution of quantitative traits: Theoretical perspectives. Evol. Appl. 2014, 7, 169–191. [Google Scholar] [CrossRef] [PubMed]
- Barraclough, T.G. How do species interactions affect evolutionary dynamics across whole communities? Annu. Rev. Ecol. Evol. Syst. 2015, 46, 25–48. [Google Scholar] [CrossRef]
- Voje, K.L.; Holen, Ø.H.; Liow, L.H.; Stenseth, N.C. The role of biotic forces in driving macroevolution: Beyond the Red Queen. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150186. [Google Scholar] [CrossRef] [PubMed]
- Urban, M.C.; Bocedi, G.; Hendry, A.P.; Mihoub, J.B.; Pe’er, G.; Singer, A.; Bridle, J.R.; Crozier, L.G.; De Meester, L.; Godsoe, W.; et al. Improving the forecast for biodiversity under climate change. Science 2016, 353, aad8466. [Google Scholar] [CrossRef] [Green Version]
- Vasseur, D.A.; Amarasekare, P.; Rudolf, V.H.; Levine, J.M. Eco-evolutionary dynamics enable coexistence via neighbor-dependent selection. Am. Nat. 2011, 178, E96–E109. [Google Scholar] [CrossRef] [Green Version]
- Kremer, C.T.; Klausmeier, C.A. Coexistence in a variable environment: Eco-evolutionary perspectives. J. Theor. Biol. 2013, 339, 14–25. [Google Scholar] [CrossRef]
- Cottenie, K. Integrating environmental and spatial processes in ecological community dynamics. Ecol. Lett. 2005, 8, 1175–1182. [Google Scholar] [CrossRef]
- Soininen, J. A quantitative analysis of species sorting across organisms and ecosystems. Ecology 2014, 95, 3284–3292. [Google Scholar] [CrossRef]
- Leibold, M.A. The metacommunity concept and its theoretical underpinnings. In The Theory of Ecology; Scheiner, S.M., Willig, M.R., Eds.; University of Chicago Press: Chicago, IL, USA, 2011; pp. 163–184. [Google Scholar]
- Heino, J.; Melo, A.S.; Siqueira, T.; Soininen, J.; Valanko, S.; Bini, L.M. Metacommunity organisation, spatial extent and dispersal in aquatic systems: Patterns, processes and prospects. Freshw. Biol. 2015, 60, 845–869. [Google Scholar] [CrossRef]
- Jones, A.G.; Bürger, R.; Arnold, S.J.; Hohenlohe, P.A.; Uyeda, J.C. The effects of stochastic and episodic movement of the optimum on the evolution of the G-matrix and the response of the trait mean to selection. J. Evol. Biol. 2012, 25, 2210–2231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, K.F.; Kremer, C.T.; Miller, E.T.; Osmond, M.M.; Litchman, E.; Klausmeier, C.A. Evolutionarily stable communities: A framework for understanding the role of trait evolution in the maintenance of diversity. Ecol. Lett. 2018, 21, 1853–1868. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.C.; Loewen, C.J.; Vinebrooke, R.D.; Chimimba, C.T. Net effects of multiple stressors in freshwater ecosystems: A meta-analysis. Glob. Chang. Biol. 2016, 22, 180–189. [Google Scholar] [CrossRef]
- Post, D.M.; Palkovacs, E.P. Eco-evolutionary feedbacks in community and ecosystem ecology: Interactions between the ecological theatre and the evolutionary play. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1629–1640. [Google Scholar] [CrossRef]
- Yoshida, T.; Jones, L.E.; Ellner, S.P.; Fussmann, G.F.; Hairston, N.G. Rapid evolution drives ecological dynamics in a predator–prey system. Nature 2003, 424, 303–306. [Google Scholar] [CrossRef]
- Turcotte, M.M.; Reznick, D.N.; Daniel Hare, J. Experimental test of an eco-evolutionary dynamic feedback loop between evolution and population density in the green peach aphid. Am. Nat. 2013, 181, S46–S57. [Google Scholar] [CrossRef] [Green Version]
- Matthews, B.; Aebischer, T.; Sullam, K.E.; Lundsgaard-Hansen, B.; Seehausen, O. Experimental evidence of an eco-evolutionary feedback during adaptive divergence. Curr. Biol. 2016, 26, 483–489. [Google Scholar] [CrossRef] [Green Version]
- Brunner, F.S.; Anaya-Rojas, J.M.; Matthews, B.; Eizaguirre, C. Experimental evidence that parasites drive eco-evolutionary feedbacks. Proc. Natl. Acad. Sci. USA 2017, 114, 3678–3683. [Google Scholar] [CrossRef] [Green Version]
- Fronhofer, E.A.; Altermatt, F. Eco-evolutionary feedbacks during experimental range expansions. Nat. Commun. 2015, 6, 6844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legrand, D.; Cote, J.; Fronhofer, E.A.; Holt, R.D.; Ronce, O.; Schtickzelle, N.; Travis, J.M.; Clobert, J.; Clobert, J. Eco-evolutionary dynamics in fragmented landscapes. Ecography 2017, 40, 9–25. [Google Scholar] [CrossRef] [Green Version]
- Terhorst, C.P.; Zee, P.C.; Heath, K.D.; Miller, T.E.; Pastore, A.I.; Patel, S.; Schreiber, S.J.; Wade, M.J.; Walsh, M.R. Evolution in a community context: Trait responses to multiple species interactions. Am. Nat. 2018, 191, 368–380. [Google Scholar] [CrossRef]
- Colombo, E.H.; Martínez-García, R.; López, C.; Hernández-García, E. Spatial eco-evolutionary feedbacks mediate coexistence in prey-predator systems. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ferriere, R.; Legendre, S. Eco-evolutionary feedbacks, adaptive dynamics and evolutionary rescue theory. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120081. [Google Scholar] [CrossRef] [Green Version]
- Ferriere, R. Adaptive Responses to Environmental Threats: Evolutionary Suicide, Insurance, and Rescue. In Options Spring 2000; International Institute for Applied Systems Analysis: Laxenburg, Austria, 2000; pp. 12–16. [Google Scholar]
- Gyllenberg, M.; Parvinen, K. Necessary and sufficient conditions for evolutionary suicide. Bull. Math. Biol. 2001, 63, 981–993. [Google Scholar] [CrossRef]
- Gyllenberg, M.; Parvinen, K.; Dieckmann, U. Evolutionary suicide and evolution of dispersal in structured metapopulations. J. Math. Biol. 2002, 45, 79–105. [Google Scholar] [CrossRef]
- Dercole, F.; Ferrière, R.; Rinaldi, S. Ecological bistability and evolutionary reversals under asymmetrical competition. Evolution 2002, 56, 1081–1090. [Google Scholar] [CrossRef] [Green Version]
- Bocedi, G.; Palmer, S.C.; Pe’er, G.; Heikkinen, R.K.; Matsinos, Y.G.; Watts, K.; Travis, J.M. Range Shifter: A platform for modelling spatial eco-evolutionary dynamics and species’ responses to environmental changes. Methods Ecol. Evol. 2014, 5, 388–396. [Google Scholar] [CrossRef]
- Wisz, M.S.; Pottier, J.; Kissling, W.D.; Pellissier, L.; Lenoir, J.; Damgaard, C.F.; Dormann, C.F.; Forchhammer, M.C.; Grytnes, J.; Guisan, A.; et al. The role of biotic interactions in shaping distributions and realised assemblages of species: Implications for species distribution modelling. Biol. Rev. 2013, 88, 15–30. [Google Scholar] [CrossRef] [Green Version]
- Pollock, L.J.; Tingley, R.; Morris, W.K.; Golding, N.; O’Hara, R.B.; Parris, K.M.; Vesk, P.A.; McCarthy, M.A. Understanding co-occurrence by modelling species simultaneously with a Joint Species Distribution Model (JSDM). Methods Ecol. Evol. 2014, 5, 397–406. [Google Scholar] [CrossRef]
- Benito Garzón, M.; Robson, T.M.; Hampe, A. ΔTrait SDMs: Species distribution models that account for local adaptation and phenotypic plasticity. New Phytol. 2019, 222, 1757–1765. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fielding, A.P.; Pantel, J.H. Eco-Evolutionary Feedbacks and the Maintenance of Metacommunity Diversity in a Changing Environment. Genes 2020, 11, 1433. https://doi.org/10.3390/genes11121433
Fielding AP, Pantel JH. Eco-Evolutionary Feedbacks and the Maintenance of Metacommunity Diversity in a Changing Environment. Genes. 2020; 11(12):1433. https://doi.org/10.3390/genes11121433
Chicago/Turabian StyleFielding, Aidan P., and Jelena H. Pantel. 2020. "Eco-Evolutionary Feedbacks and the Maintenance of Metacommunity Diversity in a Changing Environment" Genes 11, no. 12: 1433. https://doi.org/10.3390/genes11121433