1. Introduction
The world population is currently 7.8 billion and expected to increase by 2 billion by 2050. Between 1961 and 2016, the population growth (3.2%) outpaced the food production (1.6%) and also exceeded the total meat production (2.8%) [
1]. However, the fish consumption is increasing constantly at an average rate of 1.5% per year, and the UN estimate that one in five persons depends on fish as the primary source of protein. This tremendous growth depends directly on the availability of feed resources. Atlantic salmon (
Salmo salar) is a commercially important species [
1], with the major share produced in Norway, even increasing in export volume by 4% from 2019 to 2020 [
2]. Fish consumption is expected to increase in the coming years to meet the demands of the growing population. Fish is an important part of the human diet, contributing with essential amino acids, omega-3 long chain polyunsaturated fatty acids such as eicosapentaenoic acid and docosahexaenoic acid, essential minerals (Ca, P, Zn, Fe, Se, I), and vitamins (A, B, D) [
3]. Growth in aquaculture is necessarily accompanied by an increase in feed production, and consequently by a need for alternative feed ingredients. The inclusion rates of fishmeal (FM) and fish oil (FO) in the diets of Atlantic salmon have decreased from 90% in 1990 to 14.5% in 2016 [
4,
5]. The shift to plant-based feed ingredients is a direct consequence of the reduced global availability of FM and FO. Moreover, a rising market pressure to improve the sustainability of fish farming has encouraged this development [
6] and this caused the marine protein dependency ratio to decrease from 3.8 kg (1990) for 1 kg of salmon to 0.7 (2013), along with a parallel decrease in the use of marine oils [
4].
Novel protein sources, e.g., from insects, are commercially available, but plant-based materials continue to be the prevalent replacement for marine ingredients. Mostly used are soybean protein concentrate (SPC), wheat and wheat gluten (WG), along with corn, faba beans, sunflower meal, pea protein concentrate, and other vegetable proteins [
5]. The optimal growth of Atlantic salmon depends not only on the fish genetic profiles, rearing conditions, and diet composition but also on the feed formulation and processing [
7]. Feed processing has a significant impact on the nutritional quality and digestible energy of the complete diet [
8]. It has been shown that soybean meal can cause enteritis in salmon, whereas alcohol-extracted SPC appears not to affect the growth and intestinal integrity, and even enhances weight gain better than FM [
9]. However, some studies have indicated that also protein concentrates from soybean and pea can trigger moderate changes in the intestine of salmon, while comparable effects were not reported for WG [
10]. WG is a good pellet binder in extruded diets. It is highly digestible [
11] and can replace up to 35% of FM in salmonid diets without significant negative effects [
12], so that it has become widely accepted as ingredient in fish diets. However, higher levels can only be incorporated after adequate supplementation with limiting amino acids, especially lysine.
Inclusion of plant ingredients pose a potential threat to fish due to the presence of various undesirable substances including anti-nutritional factors (ANFs) such as phytoestrogens and mycotoxins as well as chemical contaminants, which can interfere with nutrient digestibility, absorption and utilisation, and negatively affect growth and health [
13]. Undesirable substances may be present in feed at low levels and thus remain unnoticed due to lack of analysis, methods with insufficient detection limits, or the non-availability of reference standards, yet their presence alone or in combination may have negative implications. Dietary exposure to harmful contaminants can be detected by transcriptomic analysis showing affected pathways. WG contains comparably less ANF than protein concentrates from legumes [
11]. However, major protein fractions in WG, i.e., the gliadins and glutenins [
14], are associated to a range of intestinal health effects and disorders in humans [
15]. So far, sensitivity to wheat gluten has not been reported in fish.
Many proteins of different origins are assessed as novel ingredients in fish feed as replacement for FM, making thorough evaluation using multidisciplinary approaches for studying effects of on fish metabolism, health, and growth performance necessary. The present study was designed to identify the impact of WG on growth, feed efficiency, metabolism, and intestinal health in Atlantic salmon. WG was administered at two inclusion levels (15% or 30%) in the diet and compared to a diet containing only FM as the protein source. Blood parameters and liver fat were analysed to identify potential health effects. Gene expression profiling by microarray analysis was performed to study the response in two metabolic important tissues, intestine and liver.
2. Materials and Methods
2.1. Preparation of Customised Salmon Diets
The fish diets were prepared at Nofima Feed Technology Centre, Fyllingsdalen, Norway by incorporating WG as the plant protein source, replacing 15% or 30% of the fishmeal (FM). The feed materials were purchased from the commercial companies Norsildmel AS (FM) (Bergen, Norway) and Tereos Syral (WG) (Marckolsheim, France). The plant ingredients used were of good quality and contained only low levels of undesirable substances such as mycotoxins, often below the limit of detection [
16]. Therefore, we assumed that any change in the gene expression in different salmon tissues was not caused by these contaminants, but was a direct consequence of the presence of WG in the diet. The manufacturing process of the experimental diets for salmon has been reported previously in detail [
16]. Diet formulations and compositions are given in
Table 1. The fish diets were formulated in such a way that contents of total proteins, dry matter, lipids, and energy were approximately the same for all diet groups. The pellet size was adjusted to the size of the on-growing salmon. All non-oil ingredients were mixed, extruded, dried, and coated with oil. Yttrium oxide was included at 0.01% as an indigestible inert marker to quantify apparent nutrient digestibility. The diets were not balanced for amino acids and the other nutrients.
2.2. Fish and Feeding Trial
The experimental set-up, feeding trial, and sampling have been previously explained in detail [
16]. Briefly, one-year-old post-smolt Atlantic salmon with a mean weight of 223 g were used in a nine-week feeding trial at the Nofima’s Research Station, Sunndalsøra, Norway. The experimental groups consisted of a control group receiving diet with FM as the only protein source, and two groups fed with WG-containing diets with, respectively, 15% or 30% replacement of FM. Fish were randomly distributed to the tanks (1 m
3;
n = 30 fish per tank, except one tank of the FM group, which had 29 fish) supplied with seawater, using three replicate tanks for each of the diet groups. Bulk weight of the fish per tank was recorded at the start of the experiment. Excess feed was collected daily from the tanks for the calculation of feed intake [
17]. Fish health, feed intake, and overall welfare were regularly monitored. The water temperature was maintained at an average of 10.6 (±0.6) °C. The oxygen level at the tank outlets was higher than 90% at study start and about 80% at study end. The water flow in each tank was set to 20 L/min.
At the end of the trial, the fish were anaesthetised with a standard dose of tricaine methanesulfonate MS222 (Sigma-Aldrich, St. Louis, MO, USA), transferred to a smaller tank and euthanised with a lethal dose of 200 mg/L of the same chemical. Individual weights, liver weights, and lengths were recorded for the sampled fish (n = 5 fish per tank, i.e., in total 15 for each of the diets) and the rest weighed in bulk. Tissues and blood samples were taken. The sampled fish were opened and the mid-intestine was dissected out and digesta removed. The intestines were rinsed clean with phosphate-buffered saline (PBS) pH 7.4 and snap-frozen in liquid nitrogen. Livers were removed, weighed, and cut into small pieces of 1 cm before snap-freezing in liquid nitrogen. The liver and intestine samples were stored at −80 °C until analysis.
Faecal samples were collected, pooled per tank, and stored at −20 °C prior to analysis for calculating apparent digestibility coefficients (ADC). The ADC of lipid, nitrogen, and energy in the experimental diets were determined by using the equation , where d is for diet, f for faeces, Y for yttrium content, and N for nutrient content. The growth rates were calculated according to the equation for the specific growth rate , where W1 is the final weight, W0 the initial weight, and d are the days in experiment, and for the thermal growth coefficient , where ddg represents degree-days, i.e., the product of the water temperature (°C) and the number of days at this temperature in the experiment. Furthermore, the feed conversion ratio (FCR = feed consumed/biomass increase), condition factor (CF = (fish weight (g)/fish fork length (cm) × 100), and hepatosomatic index (HSI = 100 × (liver weight/total fish weight) were calculated.
2.3. Measurements of Blood Parameters
Blood samples were centrifuged for serum preparations, and free fatty acids (FFA), total protein (Tprot), triglycerides (TG), alanine aminotransferase (ALT), and aspartate aminotransferase (AST) were analysed at the Central Laboratory of the Norwegian University of Life Science (NMBU), Oslo, Norway.
2.4. Chemical Analysis of Liver Fat
Total lipids were extracted as previously described [
18]. Briefly, 0.4 g liver samples of five fish in a tank were pooled, homogenised in chloroform/methanol (2:1,
v/
v), filtered, washed in isotonic saline, and quantified gravimetrically.
2.5. RNA Extraction
Total RNA was extracted from the mid-intestine and liver samples (≈10 mg) using a Biomek 4000 Automated Workstation (Beckman Coulter, Indianapolis, IN, USA), applying the Agencourt® RNAdvance tissue kit (Agencourt Bioscience Corporation, Beverly, MA, USA) according to the manufacturer’s instructions: the tissues (10 mg) were transferred into 1.2 mL collection microtubes (QIAGEN, Venio, The Netherlands) with 3 mm magnetic beads, 400 µL lysis buffer, and 1 mg proteinase K. The samples were homogenised in a tissue lyser (180 s 1800 rpm; FastPrep-96, Beckman Coulter), centrifuged (1 min, 1600 rpm, Avanti™ J-301, Beckman Coulter), and placed in a heated cabinet (Termaks, Bergen, Norway) for 25 min or more until the tissue samples were completely lysed. Subsequently, the samples were processed in the Biomek 4000 workstation. RNA concentration and quality were determined using a NanoDrop 8000 Spectrophotometer (Thermo Scientific, Bremen, Germany) and an Agilent 2100 Bioanalyzer instrument with Agilent RNA 6000 Nano Kit (Agilent Technologies, Santa Clara, CA, USA) in accordance to the respective instrument protocols. An RNA integrity number (RIN) value above 7.5 was considered as satisfactory. The RNA samples were stored at −80 °C until use.
2.6. Microarray
Nofima’s genome-wide Atlantic salmon oligonucleotide microarray with 4 × 44k 60-mer probes, manufactured by Agilent Technologies (Santa Clara, CA, USA), was used [
19]. All reagents and equipment were purchased from the same source. Two fish from each replicate tank, in total six fish per dietary group for both intestine and liver, were selected for the microarray analysis. Amplification and labelling of the total RNA (200 ng/reaction) with cyanine 3-cytidine triphosphate (CTP) was performed with the Low-Input Quick-Amp Labeling Kit (Santa Clara, CA, USA). cRNA was quantified using the NanoDrop 8000 Spectrophotometer. The Gene Expression Hybridization Kit was used for fragmentation of the labelled RNA. The samples were immediately loaded onto the microarray for hybridisation for 17 h in a hybridisation oven at 65 °C with a rotation speed of 10 rpm. The arrays were then washed for 1 min at room temperature with Gene Expression Wash Buffer I and for 1 min with Gene Expression Wash Buffer II, the latter was prewarmed and kept at 37 °C until use. The microarrays were scanned with a SureScan Microarray Scanner from the same manufacturer.
Gene expression data were processed and analysed with Nofima’s bioinformatics database STARS (Salmon and Trout Annotated Reference Sequences) [
19]. Global normalisation was performed by equalising the mean intensities of all microarrays. The normalised values for the individual features were then divided by the mean value of all samples, thus determining the specific expression ratios (ER). Since the expression changes in the WG-containing diets were correlated, just the scale being greater for the WG30 diet, the results were combined for further analysis. Finally, log
2-ER were calculated and normalised by locally weighted nonlinear regression (Lowess).
2.7. Data Analysis
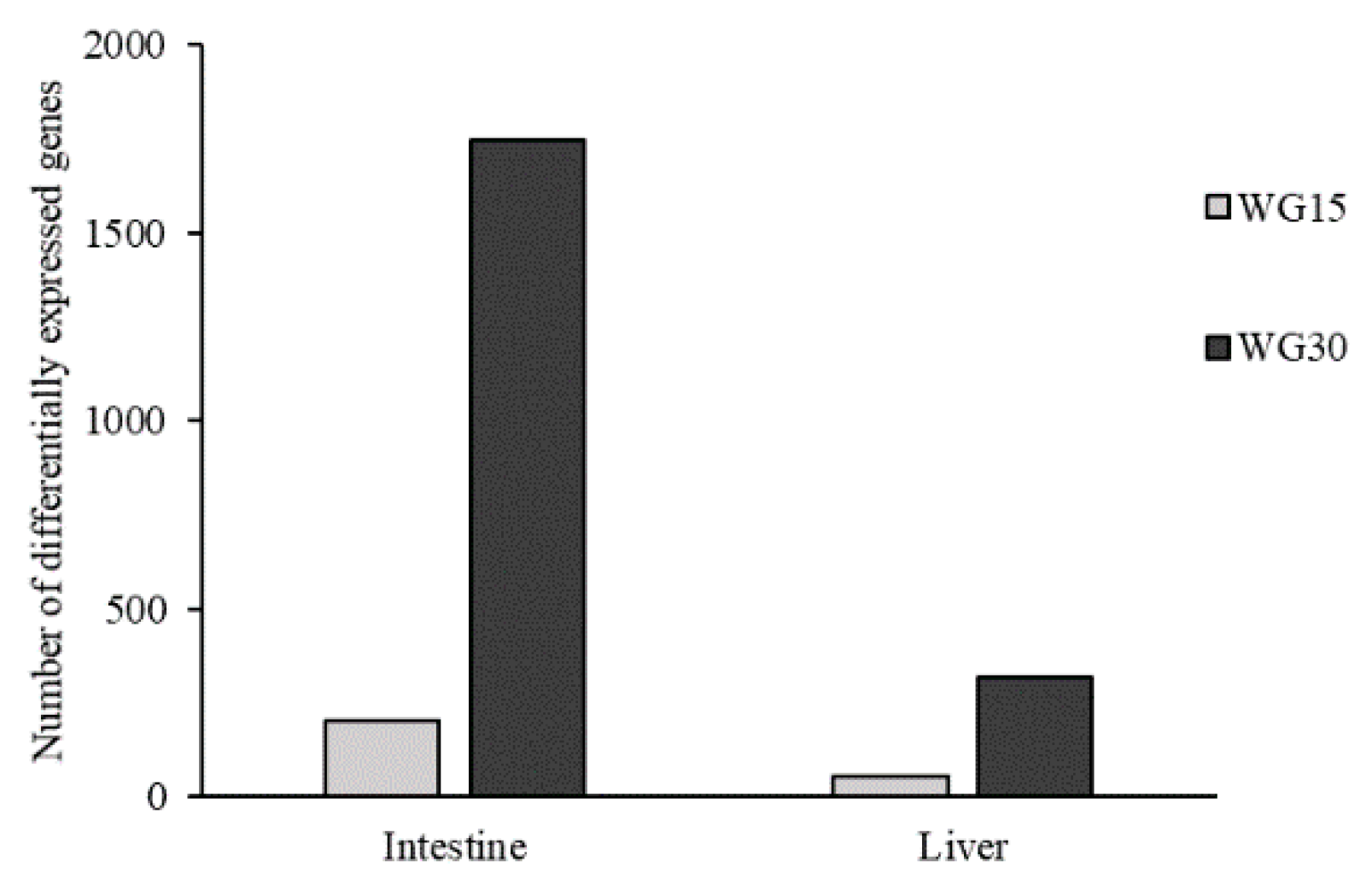
The growth rates and other parameters (body weights, feed intake, FCR, and ADC) of salmon fed with different diets were recorded for each dietary group (considering the means of three replicate tanks, n = 3) and statistically tested by one-way ANOVA to assess the effects of the diets. For comparison of the individual measured parameters from the final sampling (HSI and CF), a nested mixed model was used with diet as a fixed variable and tank as a random variable, and by using tank within diet as error term for testing. Statistical differences between the dietary groups with respect to the blood parameters and liver fat contents were evaluated by one-way ANOVA followed by Duncan’s multiple range test. In microarray analysis, differently expressed genes (DEGs) were selected by difference from control (>1.75-fold and p < 0.05). Enrichment of GO terms (Gene Ontology) and KEGG pathways (Kyoto Encyclopedia of Genes and Genomes) was assessed with Yates’ corrected chi-square test.
2.8. Ethical Statement
The study was performed in compliance with the laws regulating experimentation with live animals in Norway and the experimental protocol was approved by the Norwegian Animal Research Authority (Forsøksdyrutvalget). The methods were carried out in accordance with the relevant guidelines and regulations by EU (2010/63/EU) [
20]. The experiment was considered as not requiring a specific license from Norwegian Food Safety Authority as the fish received uncontaminated feed, were not subjected to experimental treatments, had not been exposed to any pain or distress, and were euthanised by an ethically approved method.
4. Discussion
The present study reports the effects of the dietary inclusion of the plant ingredient WG on the growth, metabolism, and overall health of Atlantic salmon. In agreement with previous studies, there were no major effects on the digestibility and growth rate of fish fed with diet at moderate inclusion (15%) of WG [
12,
21,
22]. There was, however, a significant reduction of the feed intake in the WG30 group, resulting in significantly lower growth rate and a tendency towards a decreasing final weight with the increased inclusion of WG. The lower feed intake is in agreement with the observed upregulation of cholecystokinin genes
cck in both WG groups, most pronounced in the WG30 group. Cholecystokinin is a hormone involved in the regulation of food intake and satiation and regulates the digestion of fat and protein [
23]. The differential expression of these genes as well as the growth hormone secretagogue receptors (
ghsr-a:
motilin receptor and
ghsr-1: ghrelin) indicate an effect of gluten in the diets on appetite regulation by signalling satiation that will reduce feed intake in the salmon. The increased expression of
cck might be caused by a gluten-induced metabolic disorder in the intestine. The lower feed intake in the WG groups might thus be explained by an imbalance in the intestine caused by low tolerance for gluten in salmon, which has not been reported previously. This is in contradiction to previous studies showing no signs of intestinal pathology in salmon fed with up to 35% crude WG protein [
12] or with wheat meal [
24]. A comprehensive transcriptomic analysis of effects in salmon liver and intestine connected to the dietary inclusion of WG has not been performed so far. However, changes in the expression of genes associated to muscle growth, function, metabolism, and homeostasis from exposure to WG30 have been recently determined in the fast muscle of salmon and zebrafish in a different activity of the present project [
25].
Our findings with regard to a supposed gluten sensitivity in salmon were further supported by the increased expression of genes involved in lipid metabolism and transport in fish receiving the WG-containing diets, indicating a compensatory response in the intestine and problems with the transport of lipids from the intestine to the blood circulation. Similar responses were observed in the liver, however to a lower extent, comparable to observations in previous studies [
26,
27]. Changes in lipid metabolism and transport, steroid biosynthesis, and protein synthesis by dietary inclusion of plant proteins in fish have been reported in a number of studies [
27,
28,
29].
In several vertebrate species, reduced tolerance to gluten can cause intestinal inflammation and malabsorption syndromes [
30]. In our study, high dietary levels of WG resulted in similar symptoms of intestinal lipid malabsorption as previously described for choline deficiency in Atlantic salmon [
27]. Fishmeal is the main source of choline in salmon diets. Since the customised diet in the present study had a relatively high level of FM varying from 33.4% to 63.4%, it was expected to cover the requirement for phospholipids and choline. We therefore think that the observed upregulation of several genes involved in the choline pathway (
chka,
chkb,
pcyt1b,
pmt2,
mfsd2a,
pgs1) is not a result of choline deficiency but probably a response to an imbalance in the synthesis of lipoproteins that are involved in intestinal lipid transport. Similar changes were found in the liver transcriptome of WG30-fed fish, showing altered expression of genes for phosphatidyl choline biosynthesis (
mfsd2a and
ptdss1). Thus, it can be assumed that the intestinal imbalance and metabolic disorder caused by the WG-containing diet lead also to changes in the liver metabolism.
The imbalance in the intestine induced by the WG diets is further evidenced by an upregulation in the bile acid and fatty acid metabolism, especially in the WG30 dietary group. The cholesterol biosynthetic pathway was affected in both liver and intestine of WG30-fed fish, indicating hypocholesterolaemia and an upregulated capacity for cholesterol synthesis. The observed effects are similar to those reported in previous studies on the use of plant proteins in salmon diets, where decreased lipid digestibility, reduced bile salt levels and hypocholesterolaemia were observed [
31]. The detected upregulation of the squalene and lanosterol biosynthetic pathway, producing precursors of cholesterol biosynthesis, in the WG30 group could be a compensatory mechanism for low cholesterol body pools or an imbalance in the metabolism. The upregulation of markers for cholesterol biosynthesis (
dhcr7,
srepb1,
pparγ,
hmgcs) in the WG30 group indicated increased production of cholesterol. This might be a consequence of a reduced dietary level of cholesterol due to the exchange of marine proteins by plant proteins in the diet. Alternatively, impaired cholesterol and bile acid reabsorption may result from intestinal inflammation as observed in a study on soybean meal in salmon diets [
28]. An imbalance in the cholesterol and bile acid absorption might also be the cause for the detected strong downregulation of intestinal ABC transporters (
abcb11) in the WG30 group.
Abcb11 mediates the efflux of cholesterol and bile acids from liver into bile [
28,
32], and its mRNA expression is positively regulated by bile acid receptors (
fxr) [
33]. Although the bile acid receptor level was unaffected in the liver of the WG-fed salmon, it was upregulated in the intestine along with bile acid cotransporters.
Dietary effects on intestine and liver are connected, which we could confirm with our findings regarding WG-containing salmon. Apart from transcriptome changes, we also detected that typical markers for liver damage were elevated in the WG30 dietary group. The increase was significant for ALT and close to significant for AST. The liver fat levels were within the normal range, but were notably increased by WG dietary inclusion. The HSI of fish in the WG30 group was higher than that of the other groups. This may be caused by increased fat retention or liver fatty acid synthesis, which was observed in salmon fed with mainly plant protein- and plant oil-containing diet [
34]. The assumed connection between dietary WG and an increased fatty acid synthesis in our study is supported by the detected upregulation of the transcription factor
srebp1 and the lipid metabolism genes
fadsd5,
fadsd6,
fasn,
elovl6,
fabp,
sc5d,
b3gnt5,
ddhd2, and
aacs. Acetoacetyl-coA synthetase (
aacs) catalyses the first reaction in fatty acid metabolism and plays a major role in the lipid synthesis of triacylglycerols, phospholipids, and cholesterol esters [
35]. These responses to the WG diets, in combination with the downregulation of lipid-hydrolysing lipases (
lmf2,
nape-pld,
mgl) and
cpt1, which is involved in the β-oxidation of fatty acids, might be a compensatory response to the malabsorption of lipids in the intestine [
36].
The present study also showed a high upregulation of lipid transport proteins, apolipoproteins, and perilipins, supporting an imbalance in the intestine caused by gluten sensitivity-like reactions. Deficiency in the bile transport, as discussed previously, can cause lipid droplet accumulation and triggers the increase of transport-related genes [
28]. Apolipoproteins are proteins, which bind to lipids and form lipoproteins and thereby act as transport vehicles [
37]. These proteins play an important role in the transport of cholesterol, triglycerides, phospholipids, and fat-soluble vitamins between the intestine, liver, and peripheral tissues. We observed the upregulation of several apolipoproteins in the intestine of both WG-fed groups as compared to the FM group, indicating an accumulation of lipids in the intestine and reduced transport of lipids to the circulation. A similar effect has been reported in a previous study on salmon fed with diets containing considerable levels of plant protein and oils [
38]. The increased expression of
plin2, involved in the coating of lipid droplets, in the intestine of both WG dietary groups further supports this possible link. Our findings are, however, in contrast with a salmon study on choline deficiency, where reduced intracellular lipid levels were reflected by the suppression of
plin2 expression [
27]. However, WG-containing diets seem to primarily affect the absorption and transport of lipids in the intestine.
The detected changes in several genes connected to pathways important for maintaining the intestinal balance that were induced by the WG-containing diets, especially at the high inclusion level, gave additional evidence for the proposed connection. The regulation of the genes such as
pept1,
occludin,
Rho GTPase activating proteins,
myosin,
cck,
ghsr,
cdkn1b,
fibronectin,
collagen,
mucin,
hes, and
calpain indicated a direct effect of WG on the salmon intestine. Upregulation of
pept1, involved in peptide transport across the enterocyte membrane [
39], might be a compensatory response to alterations in the intestinal membrane and increased peptide absorption. Expression of
pept1 has been shown to be affected by dietary plant protein sources in sea bream [
40].
Occludin expression was upregulated in the WG30 group and may indicate WG-induced reorganisation or strengthening of cell junctions. A similar response was observed in a study on salmon fed with pea proteins combined with soybean saponins [
41]. The
cdkn inhibitors, linked to cell cycle progression, were induced by inclusion of WG in the diet in our study, and might be a compensatory mechanism to reduce the rapid proliferation of cells in the intestine, whereas the downregulation of
fibronectin,
collagen, and
mucin genes in WG30-fed salmon might further indicate an altered intestinal integrity. The downregulation of genes belonging to extracellular matrix (
mucins and
collagen) and proteases (
hes and
calpain) involved in intestinal development and homeostasis in the WG30 group point also at a negative effect on intestinal integrity, as observed for soybean enteritis in salmon [
42].
We were also able to show that WG altered the expression of many immune genes, which has been reported in studies on chronically inflamed intestine in salmon fed with plant diets [
26,
41,
43]. We observed downregulation of the
CD28 antigen,
ctla4,
IL-17,
IL-22, T
-cell receptor-signalling pathway and
TNF-like domains,
NF-kB and
macrophage stimulating receptor genes, along with an increase in anti-inflammatory markers in the WG30 group. These findings support the assumption that salmon have a low tolerance to wheat gluten and that exposure can lead to an imbalance in the intestine with increasing inclusion of WG in the diet. Interestingly, genes related to cell apoptosis (cell death activator genes) were strongly upregulated in both WG dietary groups, and the most in the WG30 group. The upregulated
annexin A1 and annexin A2 (anxa2,
anxb11),
cannabinoid 2 receptors and
α-1-macroglobulins (
a1m) are inflammation inhibitors [
20,
36], and previous studies on humans with gluten sensitivity also showed increased plasma concentrations of these biomarkers during increased gluten intake [
44]. Moreover, the upregulation of
serum amyloid A protein (SAA5),
phospholipase A2 inhibitor-like genes (differentially-regulated trout protein 1),
arginase 1,
angionenin 1 precursor,
pstbps, and
a2m supported the initiation of a gluten-caused inflammatory effect in the salmon. DEGs that are considered as proinflammatory and anti-inflammatory markers in humans with gluten sensitivity were also upregulated in the WG30-exposed salmon. Together with the observed changes in the expression of α-macroglobulins and transglutaminase 2 (TG2), this indicated the onset of intolerance towards gluten. These genes are associated with gluten sensitivity, as they play a role in host defence mechanisms, inflammation, and protease inhibition. TG2 can catalyse the deamidation of gliadin peptides and thereby creates epitopes that are recognised by gliadin-specific T-cells in the gut [
45]. Their activation triggers autoimmune enteropathological responses leading to intestinal lesions and celiac disease [
46]. The detected downregulation of TG2 and upregulation of α-macroglobulins in the intestine of WG30-fed salmon could indicate a physiological effort to reduce the gluten-induced stress. The potential of the WG-based diet to trigger sensitivity responses in salmon is also supported by a previous experiment conducted in zebrafish using the same feed ingredients as in this study. Increased infiltration of eosinophilic granulocytes to the intestinal lumen was revealed by histology of the mid-intestine of gluten-fed zebrafish [
47]. In humans, gluten sensitivity or celiac disease has been shown to be connected to neurological dysfunctions in addition to intestinal and extra-intestinal symptoms [
48]. Similarly, the WG30-fed salmon showed a differential expression of several neural genes expressed in the mid-intestine, which can affect enteric nervous system-controlled gut functions like satiety or hunger [
47,
49]. Further studies are needed to understand the consequences of WG-based fish diets on neural functions and gut-to-brain signal transmission in fish and the impact on fish welfare.


 and
and 



{kind=link}
{kind=link}
{kind=link}