1. Introduction
Latent autoimmune diabetes in adults (LADA) is an autoimmune disease characterized by onset in adults and the presence of diabetes-associated autoantibodies but no requirement for insulin treatment for a period (usually six months) after diagnosis [
1]. In 1999, the World Health Organization (WHO) attributed LADA to type 1 diabetes mellitus (T1DM) [
2] because both of them present impairment of pancreatic islet β-cells and autoantibodies (usually positive glutamic decarboxylase-65 antibody (GADA positive)) [
3]. However, LADA shares similar clinical characteristics with type 2 diabetes mellitus (T2DM), such as onset in adulthood, increased likelihood in overweight patients, and association with rare ketosis as well as an increased risk of metabolic syndrome, such that it is often misdiagnosed as T2DM [
4]. The morbidity of LADA among patients with newly diagnosed type 2 diabetes ranges from 5.9% to 9.2% in China [
5] and 4 to 14% in Northern Europe [
1], which might prevent the best opportunity for correct treatment and intervention. Improving knowledge of the mechanisms underlying LADA will have applications for diagnosis and therapy. Although the exact etiology of LADA is still not completely understood, there is growing evidence that the interaction between environmental factors and genetic factors contributes to the risk of LADA. For example, LADA susceptibility is associated with variants in genes including PTPN22 [
6], CTLA4 [
7], SH2B3 [
8], and HLA [
9].
The major histocompatibility complex (MHC) is a group of genes located on the sixth chromosome that encode the human leukocyte antigen (HLA) system (IMGT/HLA Database:
http://www.ebi.ac.uk/imgt/hla). The
HLA system is divided into the class I, II, and III regions. The
HLA class II region consists of nearly 30 genes, including
HLA-DQB1,
HLA-DQB2,
HLA-DRA,
HLA-DRB1,
HLA-DRB2,
HLA-DRB3,
HLA-DRB4,
HLA-DRB5,
HLA-DQA1,
HLA-DQA2,
HLA-DPA1, and
HLA-DPB1 as well as less variable genes involved in antigen processing and presentation [
10]. The HLA class II region is critical in mediating the humoral immune response [
11]. Among the genes mentioned above,
HLA-DQB1 and
HLA-DRB1 are associated with many immunological diseases, such as pemphigus vulgaris [
12], narcolepsy [
13], Alzheimer’s disease [
14], and dermatomyositis [
15].
Over the past few decades, many studies have reported that the
HLA-DQB1 and
HLA-DRB1 genes are related to LADA. Pan X. et al. first proposed that the nonaspartic acid homozygote gene at position 57 of
HLA-DQB1 is associated with LADA susceptibility in Chinese populations [
16]. In previous studies, the allele groups of this gene, including
HLA-DQB1*02 [
17,
18,
19,
20],
HLA-DQB1*05 [
19],
HLA-DQB1*06 [
19,
20,
21],
HLA-DRB1*03 [
19,
20,
21,
22,
23],
HLA-DRB1*04 [
19,
20,
21,
22,
24],
HLA-DRB1*07 [
21],
HLA-DRB1*08 [
20,
21],
HLA-DRB1*09 [
20],
HLA-DRB1*12 [
19,
20,
21], and
HLA-DRB1*13 [
19], were reported to be associated with LADA. However, contradictory conclusions were drawn regarding these HLA allele groups. For example,
DQB1*02 was not associated with LADA risk in four previous studies of European populations [
21,
22,
23,
24], but the data from four other studies, including one European study and three Asian studies, indicated that
DQB1*02 increases susceptibility to LADA [
17,
18,
19,
20].
DQB1*06 was shown to have a protective effect against LADA in three studies, including one of Asians [
20] and two of Europeans [
19,
21], yet another four studies, including two European studies [
23,
24] and two Asian studies [
17,
18], did not find any association.
DRB1*03 was found to increase susceptibility to LADA in four European studies [
19,
21,
22,
23] and one Asian study [
20], but its effect was not significant in the European population study by Vatay A. et al.
DRB1*04 was reported in five studies, including four among Europeans and one among Asians, to play a role in the risk of developing LADA [
19,
20,
21,
22,
24], but Cejkova P. et al. [
23] reported no significant correlation between
DRB1*04 and LADA among Europeans.
The previous conclusions drawn regarding the correlations between these genetic polymorphisms and LADA are inconsistent, possibly due to the small sample sizes used in the individual studies. In addition, when strong associations are found between certain alleles and LADA, it may be difficult to determine whether an allele is truly protective or if the observed effect is actually the result of another allele that has a stronger impact. Less effective alleles can also be masked by stronger alleles in a similar way. These effects have probably led to the contradictory conclusions in previous studies on
DQB1 and
DRB1. The relative predispositional effect (RPE) method, which can identify the sequential association between alleles and LADA, may clarify the actual associations between alleles and LADA, regardless of the strength of the associations [
25].
No meta-analysis or systematic review has provided a precise assessment of the relationships between HLA-DQB1 and HLA-DRB1 polymorphisms and LADA. We performed this study to determine these relationships by combining meta-analysis and RPE methods. Our aim was to provide a better understanding of the etiology and pathogenesis of LADA as well as the assessment and diagnosis of LADA in order to predict high-risk LADA patients with polymorphisms of these two HLA genes.
4. Discussion
LADA is an autoimmune disease, the pathogenesis of which is currently unclear but is likely the result of environmental factors interacting with genetic susceptibility. Studies have been conducted on the association between
HLA-DQB1 or
HLA-DRB1 polymorphisms and the risk of LADA, but their outcomes were conflicting. On the one hand, although LADA is the most frequent form of adult-onset autoimmune diabetes [
1], the diagnosis of LADA is still a challenge in clinics [
4], leading to difficulty in the collection of data regarding related genes. The inconsistent results from previous studies seem to be mainly due to the small sample sizes used in the studies leading to low statistical power. On the other hand,
HLA is a highly heterogeneous gene, with 1142 and 2103 alleles in the
DQB1 and
DRB1 regions, respectively [
33], and easily misinterpreted variation in the frequency of some alleles because of the presence of a strong disease-associated allele or allele frequencies that are too low to calculate [
34]. Thus, we used meta-analysis to integrate the results of independent studies and employed the RPE method to sequentially detect associated allele groups in order to minimize deviation. Briefly, the combination of meta-analysis and RPE methods can determine the relations between LADA and
HLA-DQB1 or
HLA-DRB1 polymorphisms more accurately and more consistently.
Consistent results from both the meta-analysis and RPE methods indicate that some allele groups of the
HLA-DQB1 gene confer strong susceptibility to LADA. In our study,
HLA-DQB1*02 was demonstrated to be the main allele group in the
DQB1 region predisposing patients to LADA, although moderate heterogeneity was found among the included studies (
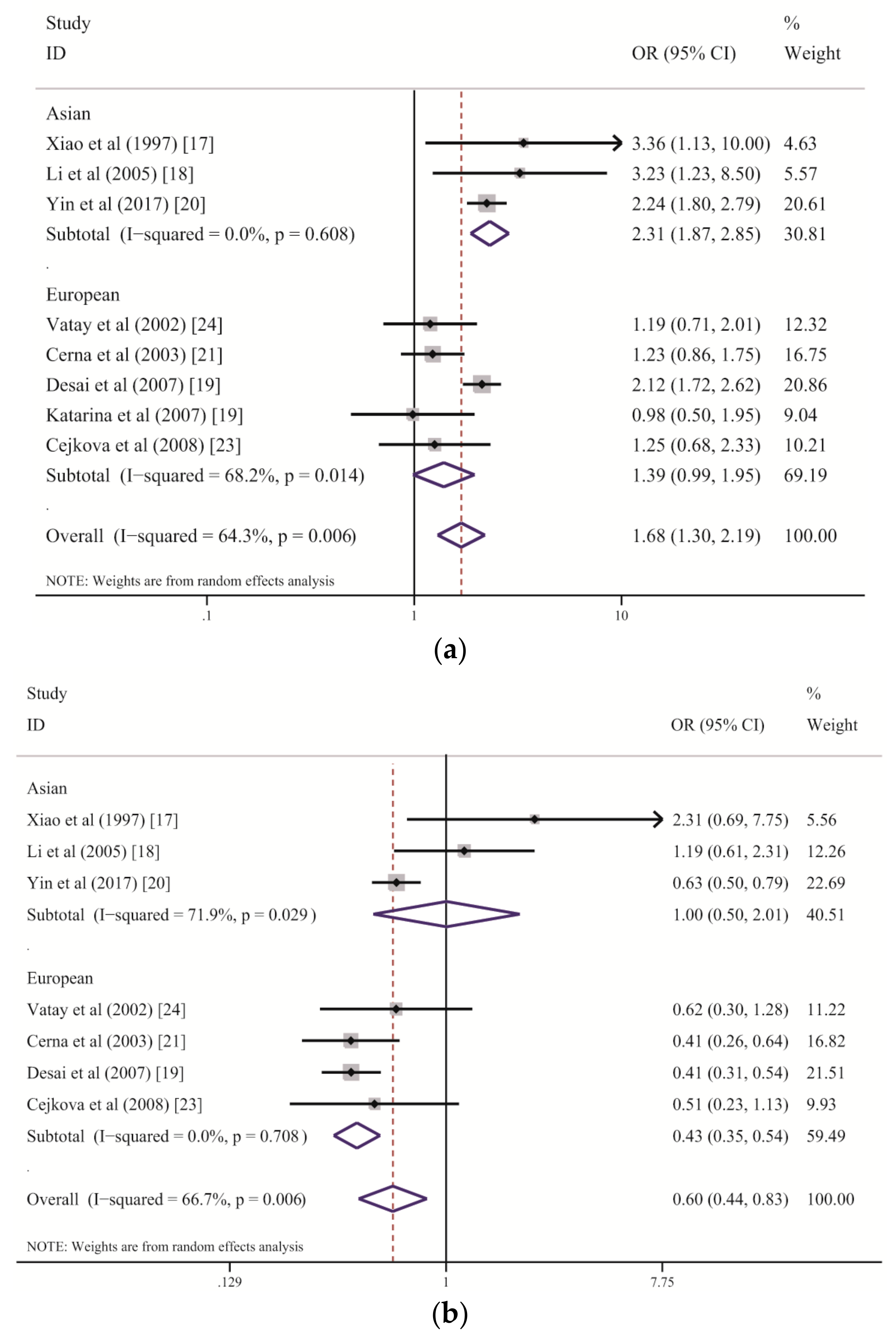
Table 2). The subgroup analysis stratified by ethnicity suggested that
DQB1*02 increases the risk of LADA among the Chinese Han population but has only a marginal association with LADA among people of European descent (
Figure 3a and
Table 3). Genetic and humoral heterogeneity between LADA and HLA genes were also found in previous studies. In Chinese studies [
17,
18,
20], all correlations between
DQB1*02 and the risk of LADA were consistent (
Figure 3a). In contrast, three of five eligible studies of European populations demonstrated slightly higher frequencies of
DQB1*02 among patients with LADA than among the controls but with wide CIs that marginally crossed one [
21,
23,
24]; the study by Desai M. et al. was weighted the heaviest and displayed a significant association [
19]. These results suggest that larger sample sizes are required to detect the role of
DQB1*02 among Europeans because its effect strength in European populations is weaker than that in Chinese populations. In fact, the combined ORs suggest that the
DQB1*02 allele group confers a two-fold higher risk of LADA in the Chinese populations than in the European populations (Chinese: OR = 2.31; European: OR = 1.39). We further analyzed
DQB1*02:01 separately (7 of 8 studies) and found a significant association of this allele with the risk of LADA (OR = 2.015, 95% CI: 1.459–2.784,
p < 0.001, not shown in the tables). The above results suggest that
HLA-DQB1*02 is a stable allele group predisposing patients to LADA, although its effect strength varies across ethnicities.
In contrast to the predisposing effect of
DQB1*02 with regard to LADA,
DQB1*06 within the
HLA-DQB1 gene plays a significantly protective role, as revealed by both the meta-analysis and RPE methods, although a high level of heterogeneity was found among studies (
Table 2 and
Figure 3b). The subgroup analysis stratified by ethnicity demonstrated an association between decreased susceptibility to LADA and
DQB1*06 in the European populations, with no heterogeneity, but there was no effect of this allele group on the disease in the Chinese populations, with a high degree of between-study heterogeneity (
Table 3 and
Figure 3b). We suspect that the variance among populations is primarily due to environmental differences and the effect strength of
DQB1*02.
DQB1*06 showed a strong protective role against LADA among the European populations because of the relatively weak role played by
DQB1*02; however, its protective effect was substantially reduced by the stronger effect of
DQB1*02 among the Chinese populations. This finding was also confirmed by the RPE analysis. Consequently,
DQB1*06 is an allele group that confers resistance to LADA but that might be influenced by
DQB1*02.
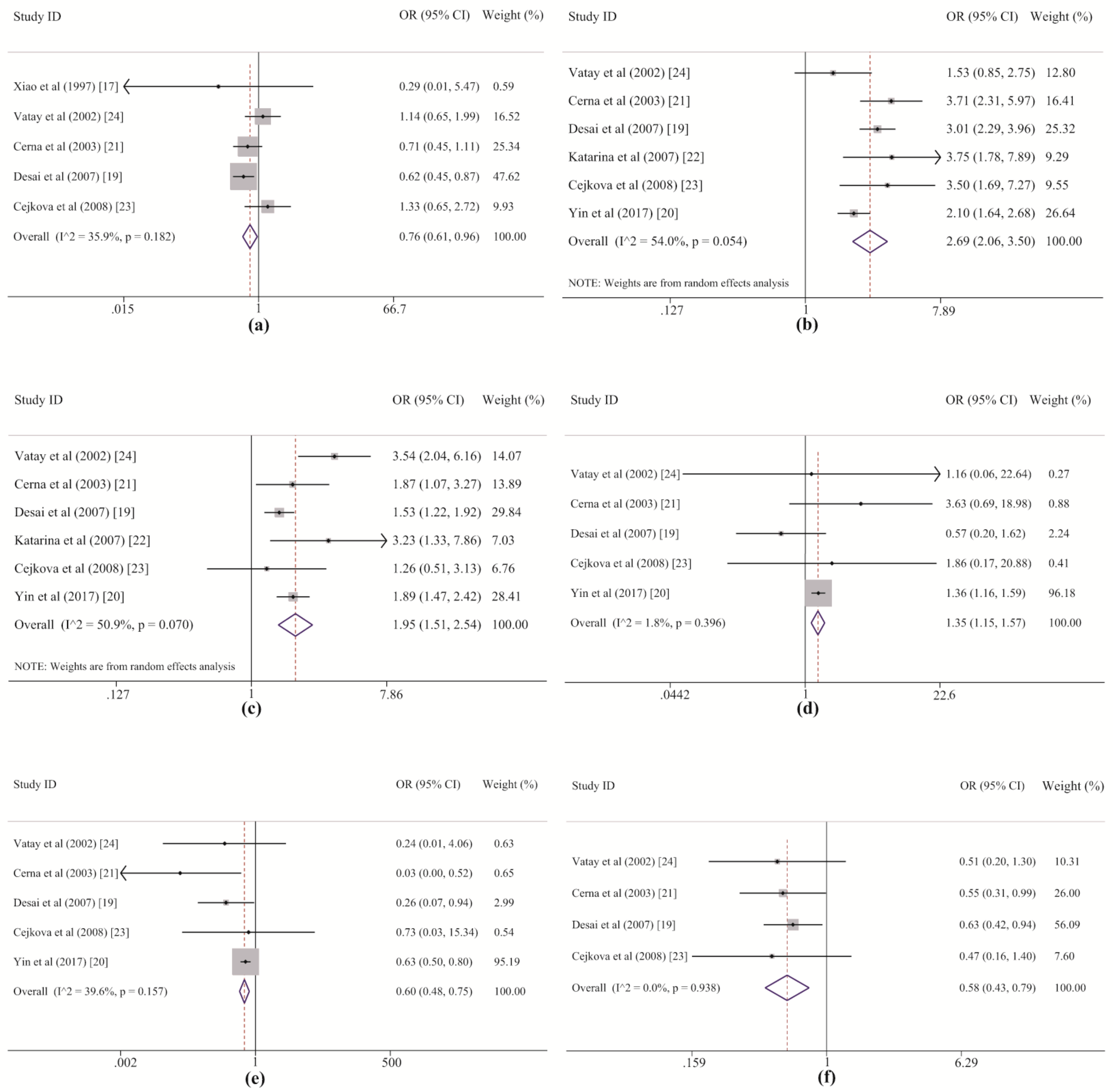
Our meta-analysis initially suggested a protective effect of
DQB1*05 against LADA, but this effect disappeared after Bonferroni correction. RPE analysis further showed the protective effect of this allele group after removing
DQB1*02 and
DQB1*06. Thus,
DQB1*05 might play a protective role through an alternate pathway that is independent of
DQB1*02 and
DQB1*06. With regard to the
DQB1*05 allele group, the heterogeneity between studies was at an acceptable level (
Table 2 and
Figure 4a). The dataset from the study by Desai M. et al. (2007) [
19] revealed a correlation between LADA and
DQB1*05 among Europeans, but no statistically significant associations were found in the other studies, including three in Europeans [
21,
23,
24] and one in Asians [
17]. Additionally, no effects of
DQB1*05 alleles on the risk of LADA were detected in previous studies [
17,
19,
21]. These prior studies may have failed to demonstrate a correlation between LADA and
DQB1*05 because the weak effect of the allele group led to low statistical power for the test and was easily masked by the strong effect of
DQB1*02. This point is also supported by the results of the sensitivity analysis, which suggested that this association was dependent on the two most heavily weighted studies (
Table S4 and
Figure 4a). Therefore, we conclude that
DQB1*05 likely plays a protective role against the development of LADA, but its effect is weak and easily masked by the stronger effects of
DQB1*02 and
DQB1*06.
In this systematic study,
DRB1*03,
DRB1*04, and
DRB1*09 within the
HLA-DRB1 gene were also demonstrated to strongly confer susceptibility to LADA in both the meta-analysis and the RPE analysis, and
DRB1*03 was revealed as the allele group in the
DRB1 region with the strongest effect on LADA. For
DRB1*03 and
DRB1*04, the heterogeneity among the included studies was acceptable. Five of six of the original datasets showed good agreement with our results, and only the studies of European populations by Vatay A. et al. (2002) [
24] and Cejkova P. et al. [
23] respectively displayed marginal associations since their individual ORs were larger than one but their individual CIs crossed one slightly (
Table 5 and
Figure 4b,c). In the subgroup analysis by ethnicity, both
DRB1*03 and
DRB1*04 increased susceptibility to LADA in Asian and European populations, but high heterogeneity was observed for
DRB1*04 among five studies in Europeans (
Table 6). The highest statistical power (value of one) was detected in the associations between the risk of LADA and the
DRB1*03 and
DRB1*04 frequencies, indicating that these results are robust. For
DRB1*09, only minor heterogeneity was found among the included studies (
Table 5).
DRB1*09 showed a significant association with an increased risk of LADA in the study by Yin N.N. et al. (2017) [
20] among Asian populations (
p < 0.001,
pc < 0.013) but not among individuals of European origin (
p < 0.904), for which none of four studies revealed a statistically significant association with the disease [
19,
21,
23,
24] (
Table 6 and
Figure 4d). The sensitivity analysis also revealed that the significance of the association changed after removing Yin et al.’s study. In addition, it is worth noting that the frequency of
DRB1*09 was much lower in the European populations than in the Asian populations (European: 0.010; Asian: 0.226, shown in
Table S6) and that the total numbers of cases and controls from the Asian study were even larger than those from four European studies (LADA: Asian: 652; European: 518; controls: Asian: 1171; European: 869, which can be derived from
Table 1) in this meta-analysis. Therefore, based on the above results, three conclusions can be drawn. First,
DRB1*09 is an allele group that predisposes Asians, especially Chinese populations, to LADA, even though only one study was performed to investigate their association. Second,
DRB1*09 may not play a role in the disease for Europeans. Third, we prefer to consider the positive association rather than the negative association between this allele group and the risk of LADA in the overall population because the former association had a statistical power of 0.723 that was further confirmed by RPE analysis. However, these conclusions should be treated with caution, and further studies with larger sample sizes should be conducted to verify them in the future.
The meta-analysis and RPE analysis also led to contrasting conclusions. In the meta-analysis, both
DRB1*12 and
DRB1*13 conferred protection against LADA. For
DRB1*12, three studies (one by Yin N.N. et al. of an Asian population and one each by Cerna M. et al. and Desai M. et al. of European populations) reported a protective role for
DRB1*12 [
19,
21], but the other two European datasets from Cejkova P et al. and Vatay A et al. showed negative results (
Figure 4e). However, although the associations could not be demonstrated in these two studies, because the frequencies were <5% in both the LADA and control groups [
23,
24], the OR for
DRB1*12 was still lower than one, indicating a possible protective role (
Figure 4e). For
DRB1*13, allelic data from the studies of European populations by Cerna M. et al. and Desai M. et al. also showed significantly lower susceptibility to LADA among
DRB1*13 carriers than among noncarriers; in contrast, although the other two European studies by Cejkova P et al. and Vatay A et al. also showed a strong protective role of
DRB1*13, with very small ORs (0.47 and 0.51), the effects were not statistically significant (
Figure 4f). The results of the meta-analysis for these two allele groups reinforced their protective role in LADA, indicating the advantage of the method, by which it is possible to properly evaluate real genetic effects on disease development with greater statistical power than that afforded by individual analyses pooling all samples or synthesizing the overall data available in previous studies. However, the results of the RPE analysis suggested that the protective effects of
DRB1*12 and
DRB1*13 were somewhat dependent on the susceptibility conferred by
DRB1*03 and
DRB1*04. Regardless of whether this dependence exists, the understanding of the associations between
DRB1*12 and
DRB1*13 and the risk of LADA in addition to the interactions between the different alleles will benefit from further studies with larger sample sizes and more haplotype data.
There were also contradictory results from the meta-analysis and the RPE analysis with regard to the associations of LADA with
DRB1*01,
DRB1*07, and
DRB1*08. The meta-analysis suggested
DRB1*01 and
DRB1*07 have protective effects against LADA, with marginal or no statistical significance (
Table 5); however, the RPE analysis indicated roles played by
DRB1*01 and
DRB1*07 in susceptibility to LADA in the fourth and sixth rounds, respectively. These relationships may be explained by the relatively weak effect of
DRB1*01 and
DRB1*07 on the risk of LADA compared with that of
DRB1*03,
DRB1*04, and
DRB1*09. Based on the above results,
DRB1*01 and
DRB1*07 might promote the development of LADA. Thus, these substantially different results from the RPE method compared to those from meta-analysis or the individual studies might reflect the advantages of RPE analysis, which allows us to sequentially detect the associated alleles or allele group and minimize the variation caused by some specific loci or locus groups. Although the results from RPE analysis suggested that
DRB1*08 increased disease risk, the pooled results of the meta-analysis showed no association with LADA, with a high degree of heterogeneity between studies. In contrast, in the sensitivity analysis, the results showed a protective role of
DRB1*08 against the development of LADA after the European study by Cerna M. et al. [
21] was removed, which also significantly reduced the heterogeneity (
I2 = 37.0%, shown in
Table S4). In addition, the distinctive characteristics of cases and controls in individual studies, such as differences in the age of onset, varying degrees of disease development among patients, and dissimilarities in allele group frequency distributions among regions, might explain this high heterogeneity. However, the currently available evidence is insufficient to indicate whether LADA is related to the frequency of
DRB1*08. Consequently, the roles played by
DRB1*01,
DRB1*07, and
DRB1*08 in the development of LADA still require more data for verification.
The above allele groups, including
DQB1*02,
DQB1*05,
DQB1*06,
DRB1*01,
DRB1*03,
DRB1*04,
DRB1*07,
DRB1*08,
DRB1*09,
DRB1*12, and
DRB1*13, were found to be associated with LADA risk to some extent by the meta-analysis and/or RPE method in our study. The groups were also previously correlated with other types of autoimmune diseases. For example,
DQB1*02 and
DQB1*05 are significantly associated with a decreased risk and an increased risk of pemphigus vulgaris (PV), respectively [
35].
DQB1*06 and
DRB1*13 have a protective effect against rheumatoid arthritis (RA), while
DRB1*01 and
DRB1*04 have a detrimental effect [
36,
37,
38].
DRB1*03 and
DRB1*12 are respectively protective against or predisposing to autoimmune thyroid diseases (AITDs) [
39].
DRB1*07 and
DRB1*08 are significantly correlated with the development of type 2 and type 1 autoimmune hepatitis (AIH), respectively [
40], and
DRB1*08 also increases susceptibility to juvenile idiopathic arthritis (JIA) [
41]. In addition,
DRB1*09 was detected to be a risk factor for systemic lupus erythematosus (SLE) [
42]. Notably, the effects of these allele groups on susceptibility to other types of autoimmune diseases were similar to those on susceptibility to LADA.
Finally, both the meta-analysis and the RPE method demonstrated a lack of correlations between the other HLA genetic allele groups and LADA. We did not find any associations of
HLA-DQB1*03,
HLA-DQB1*04,
HLA-DRB1*10,
HLA-DRB1*11,
HLA-DRB1*14,
HLA-DRB1*15, and
HLA-DRB1*16 with the risk of LADA when using either method, although some studies reported that these polymorphisms may be related to other autoimmune diseases, such as multiple sclerosis (MS) [
43], Henoch–Schönlein purpura (HSP) [
44], and T1DM [
45].
In the past, LADA was considered to be a form of adult-onset T1DM, with many clinical features similar to those of T2DM. However, some evidence indicates that there is likely a different pathological mechanism driving LADA than either T1DM or T2DM [
4,
46,
47]. Furthermore, there is genetic, phenotypic, and humoral heterogeneity in the degrees of insulin resistance and autoimmunity. Comparing our conclusion with that of a previously published meta-analysis on the association between HLA class II alleles and T1DM in Latin American patients [
45], these two diseases display some heterogeneity in their correlations with
HLA-DQB1 and
HLA-DRB1. First, although
DQB1*02,
DRB1*03, and
DRB1*04 increase an individual’s susceptibility to both LADA and T1DM, while
DQB1*05,
DQB1*06, and
DRB1*13 protect an individual from developing LADA and T1DM, these loci clearly have a stronger impact on T1DM than on LADA (e.g.,
DRB1*13: LADA: OR = 0.583; T1DM: OR = 0.300 [
45]). In addition, the allele groups conferring susceptibility to or protection against LADA and T1DM differ. For example,
DRB1*09 increases susceptibility to LADA, but this variant is not significantly associated with T1DM. In contrast,
DQB1*03 increases susceptibility to T1DM, while
DRB1*14 and
DRB1*15 have protective effects; there are no obvious correlations between these variants and LADA. Moreover, some particular allele groups have opposite effects on the risk of LADA and T1DM. Based on our conclusions,
DRB1*12 decreases the risk of developing LADA; however, it was shown to increase the risk of T1DM [
45]. The abovementioned results partly support the view that LADA might have a different etiology than T1DM. However, the variance observed in the associations of T1DM and LADA with
HLA-DQB1 and
HLA-DRB1 could also be due to differences in the inclusion criteria and study design (such as diagnostic criteria, gender, and genotyping method) as well as statistical power and ethnicity. Thus, this conclusion needs to be treated with caution, and more evidence is needed to verify our hypothesis in the future.
The limitations of this retrospective study warrant further discussion. First, in the meta-analysis, high between-study heterogeneity was detected in the association between some allele groups, such as DRB1*07 and DRB1*11, and the risk for LADA; however, we failed to find the source of this heterogeneity through meta-regression, subgroup analysis, and sensitivity analysis. Second, although we performed a thorough and meticulous search to identify all available literature, we obtained only nine studies that fit the eligibility criteria for this meta-analysis. We even contacted the authors of original studies lacking sufficient allele data, but we did not receive a reply. Third, the small sample size of several case–control studies included here is an unavoidable limitation of the meta-analysis, and more studies with larger sample sizes are needed to support future retrospective analysis. Fourth, data on the HLA-DQB1 and HLA-DRB1 alleles (with four digits and high resolution) have been provided in increasing the numbers of original studies, but the data are still too limited for further exploration of the relationships of these alleles with the risk of developing LADA. Fifth, previous studies also reported relationships between the HLA-DQB1 and HLA-DRB1 genotypes, haplotypes, and LADA, but it is difficult to systematically evaluate these relationships due to sparse data. Therefore, future research should focus on the interplay between the HLA-DQB1 and HLA-DRB1 genes as well as alleles (with four digits) and investigate the stability of their associations with LADA under different conditions of environmental factors, genetic background, and experimental design.







{kind=link}
{kind=link}
{kind=link}
{kind=link}