Chicken Embryonic-Stem Cells Are Permissive to Poxvirus Recombinant Vaccine Vectors


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
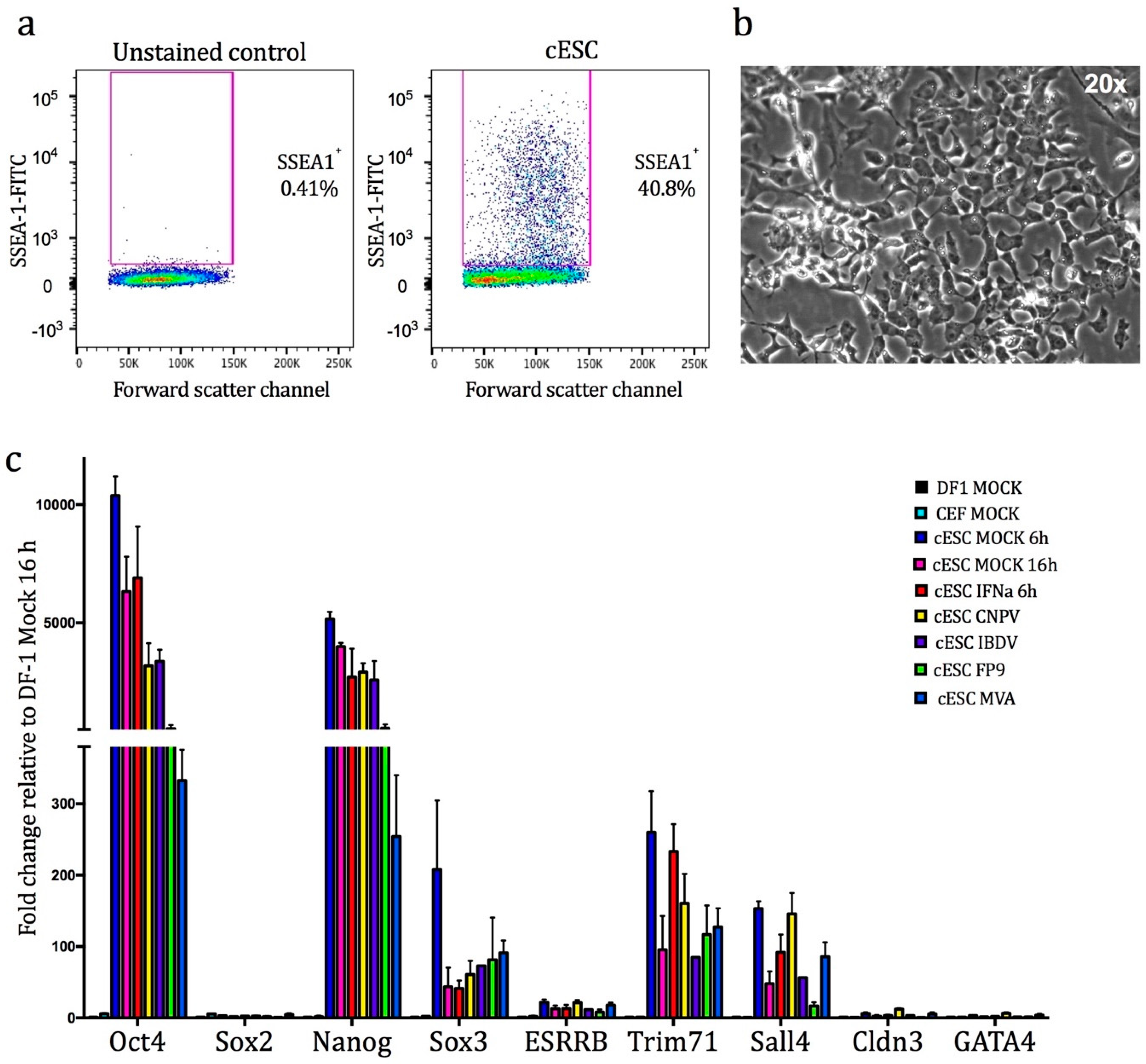
2.2. Confirmation of Pluripotency
2.3. Virus Propagation, Titration, and Purification
2.4. Viral Infections and chIFN-α Stimulation
2.5. RNA Extraction and Processing of Samples for RNA-Sequencing
2.6. IIIumina cDNA Library Preparation and Sequencing
2.7. Bioinformatic Analyses
2.8. Quantitative Real-Time RT PCR
2.9. Statistical Analyses
3. Results and Discussion
3.1. Confirmation of the Pluripotent Status of cESC
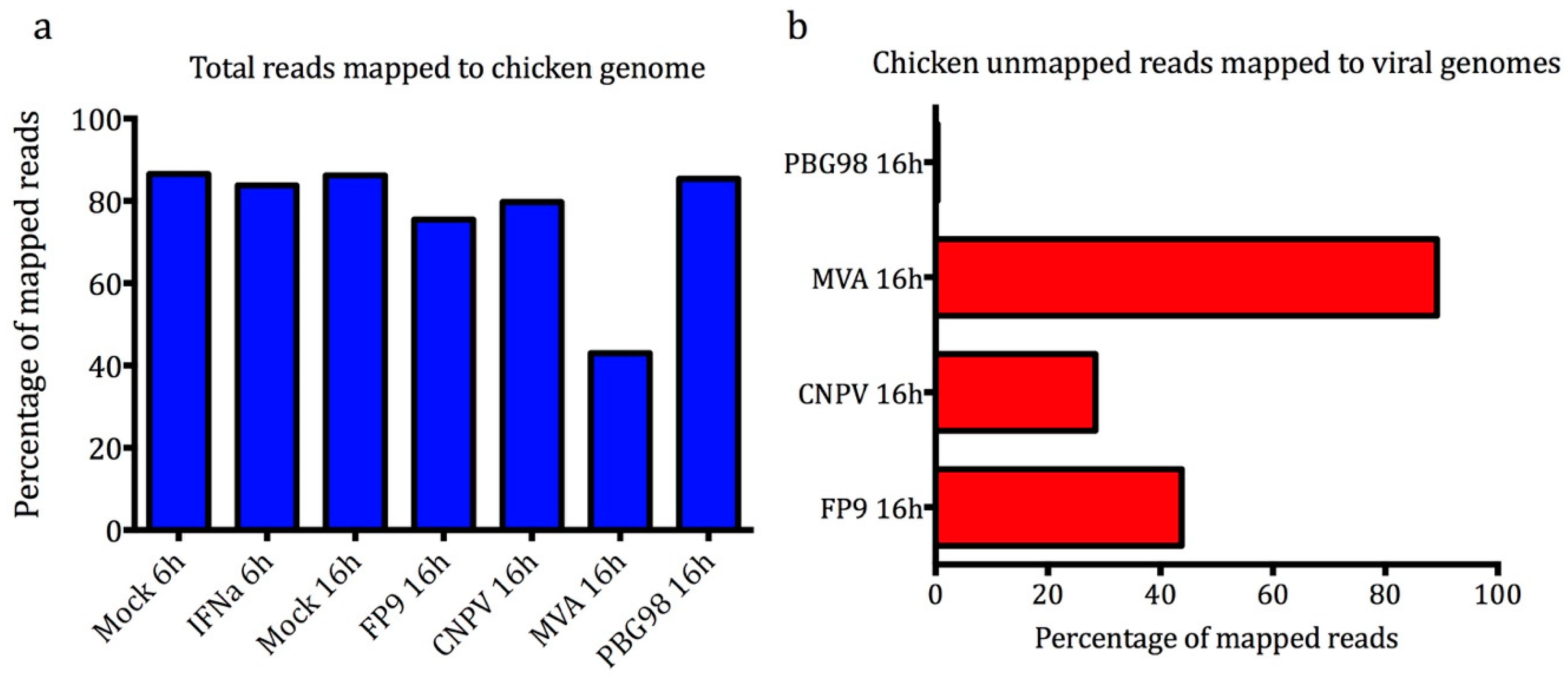
3.2. Illumina 100b Paired-End Sequencing and Sequence Quality Control
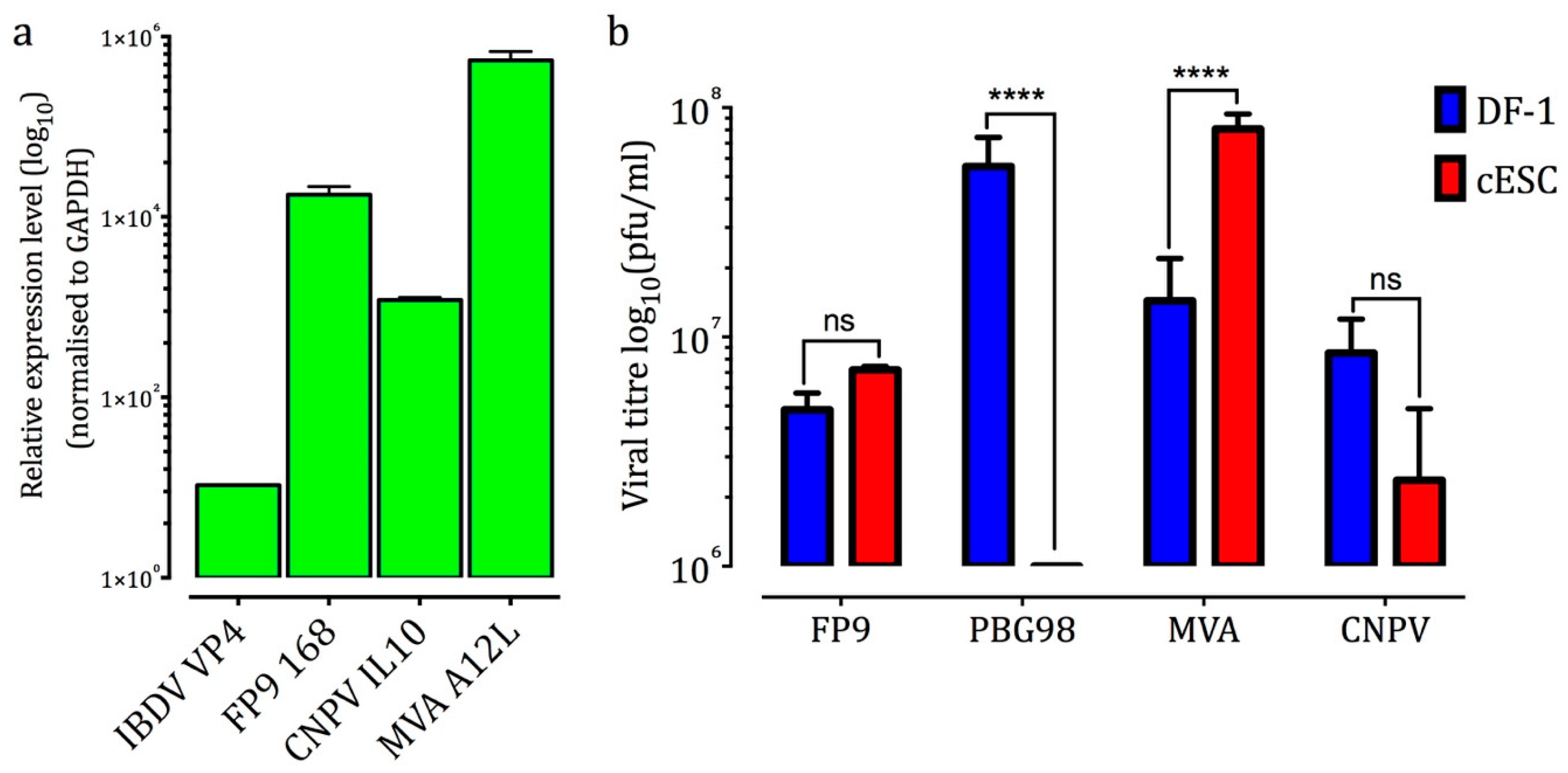
3.3. cESC Are Permissive to MVA, FP9, and CNPV But Not IBDV PBG98
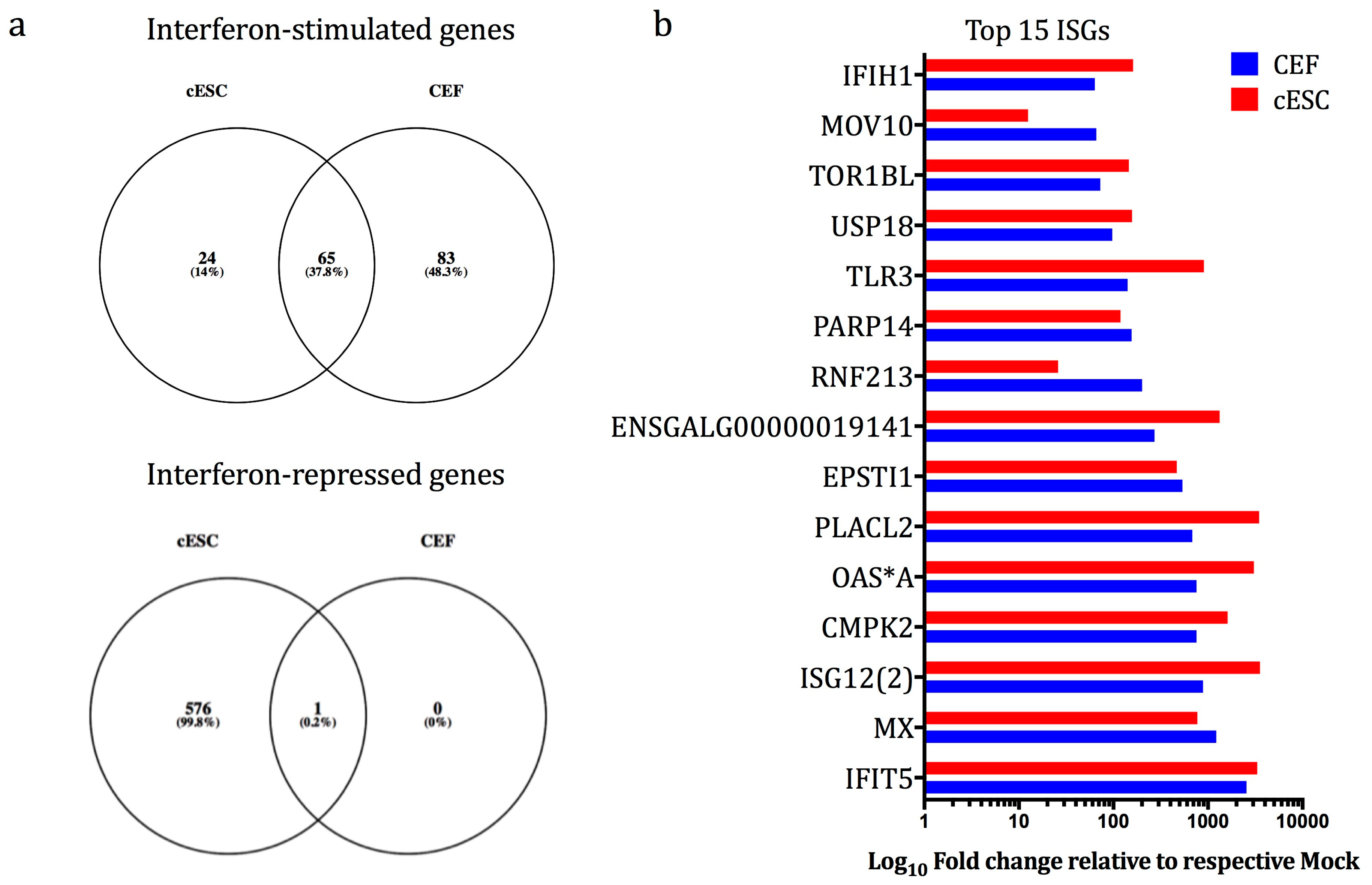
3.4. cESC are Competent for High-Level IFN-Mediated Induction of ISGs
3.5. The Vaccine Vector Viruses Induce Distinct Gene Expression Profiles in cESC
3.6. Correlation Analysis between RNA-seq and qRT-PCR Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Ethical Statement
References
- Genzel, Y. Designing cell lines for viral vaccine production: Where do we stand? Biotechnol. J. 2015, 10, 728–740. [Google Scholar] [CrossRef]
- Brown, S.W.; Mehtali, M. The avian EB66® cell line, application to vaccines, and therapeutic protein production. PDA J. Pharm. Sci. Technol. 2010, 64, 419–425. [Google Scholar]
- Naruse, T.; Fukuda, T.; Tanabe, T.; Ichikawa, M.; Oda, Y.; Tochihara, S.; Kimachi, K.; Kino, Y.; Ueda, K. A clinical phase I study of an EB66 cell-derived H5N1 pandemic vaccine adjuvanted with AS03. Vaccine 2015, 33, 6078–6084. [Google Scholar] [CrossRef] [PubMed]
- Olivier, S.; Jacoby, M.; Brillon, C.; Bouletreau, S.; Mollet, T.; Nerriere, O.; Angel, A.; Danet, S.; Souttou, B.; Guehenneux, F.; et al. EB66 cell line, a duck embryonic stem cell-derived substrate for the industrial production of therapeutic monoclonal antibodies with enhanced ADCC activity. MAbs 2010, 2, 405–415. [Google Scholar] [CrossRef]
- Jordan, I.; Vos, A.; Beilfuss, S.; Neubert, A.; Breul, S.; Sandig, V. An avian cell line designed for production of highly attenuated viruses. Vaccine 2009, 27, 748–756. [Google Scholar] [CrossRef]
- Pain, B.; Clark, M.E.; Shen, M.; Nakazawa, H.; Sakurai, M.; Samarut, J.; Etches, R.J. Long-term in vitro culture and characterisation of avian embryonic stem cells with multiple morphogenetic potentialities. Development 1996, 122, 2339–2348. [Google Scholar]
- Lavial, F.; Pain, B. Chicken embryonic stem cells as a non-mammalian embryonic stem cell model. Dev. Growth Differ. 2010, 52, 101–114. [Google Scholar] [CrossRef]
- Aubel, P.; Pain, B. Chicken embryonic stem cells: Establishment and characterization. Methods Mol. Biol. 2013, 1074, 137–150. [Google Scholar]
- Pain, B.; Kress, C.; Rival-Gervier, S. Pluripotency in avian species. Int. J. Dev. Biol. 2018, 62, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Schuind, A.; Segall, N.; Drame, M.; Innis, B.L. Immunogenicity and safety of an EB66 cell-culture-derived Influenza A/Indonesia/5/2005(H5N1) AS03-adjuvanted vaccine: A phase 1 randomized trial. J. Infect. Dis. 2015, 212, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Mehtali, M.; Champion-Arnaud, P.; Arnaud, L. Production of viral vaccines in suspension on avian embryonic derived stem cell lines. U.S. Patent No. 9040296, 26 May 2015. [Google Scholar]
- Jean, C.; Oliveira, N.M.; Intarapat, S.; Fuet, A.; Mazoyer, C.; De Almeida, I.; Trevers, K.; Boast, S.; Aubel, P.; Bertocchini, F.; et al. Transcriptome analysis of chicken ES, blastodermal and germ cells reveals that chick ES cells are equivalent to mouse ES cells rather than EpiSC. Stem Cell Res. 2015, 14, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Fuet, A.; Montillet, G.; Jean, C.; Aubel, P.; Kress, C.; Rival-Gervier, S.; Pain, B. NANOG is required for the long-term establishment of avian somatic reprogrammed cells. Stem Cell Rep. 2018, 11, 1272–1286. [Google Scholar] [CrossRef] [PubMed]
- Pain, B.; Chenevier, P.; Samarut, J. Chicken embryonic stem cells and transgenic strategies. Cells Tissues Org. 1999, 165, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Giotis, E.S.; Skinner, M.A. Spotlight on avian pathology: Fowlpox virus. Avian Pathol. 2018, 87–90. [Google Scholar] [CrossRef]
- Laidlaw, S.M.; Skinner, M.A. Comparison of the genome sequence of FP9, an attenuated, tissue culture-adapted European strain of Fowlpox virus, with those of virulent American and European viruses. J. Gen. Virol. 2004, 85, 305–322. [Google Scholar] [CrossRef]
- Jarmin, S.; Manvell, R.; Gough, R.E.; Laidlaw, S.M.; Skinner, M.A. Avipoxvirus phylogenetics: Identification of a PCR length polymorphism that discriminates between the two major clades. J. Gen. Virol. 2006, 87, 2191–2201. [Google Scholar] [CrossRef]
- Stickl, H.; Hochstein-Mintzel, V.; Mayr, A.; Huber, H.C.; Schafer, H.; Holzner, A. MVA vaccination against smallpox: Clinical tests with an attenuated live vaccinia virus strain (MVA) (author’s transl). Dtsch. Med. Wochenschr. 1974, 99, 2386–2392. [Google Scholar] [CrossRef]
- Brown, M.D.; Skinner, M.A. Coding sequences of both genome segments of a European ‘very virulent’ infectious bursal disease virus. Virus Res. 1996, 40, 1–15. [Google Scholar] [CrossRef]
- Fernandez-Arias, A.; Risco, C.; Martinez, S.; Albar, J.P.; Rodriguez, J.F. Expression of ORF A1 of infectious bursal disease virus results in the formation of virus-like particles. J. Gen. Virol. 1998, 79, 1047–1054. [Google Scholar] [CrossRef]
- Buttigieg, K.; Laidlaw, S.M.; Ross, C.; Davies, M.; Goodbourn, S.; Skinner, M.A. Genetic screen of a library of chimeric poxviruses identifies an ankyrin repeat protein involved in resistance to the avian type I interferon response. J. Virol. 2013, 87, 5028–5040. [Google Scholar] [CrossRef]
- Laidlaw, S.M.; Robey, R.; Davies, M.; Giotis, E.S.; Ross, C.; Buttigieg, K.; Goodbourn, S.; Skinner, M.A. Genetic screen of a mutant poxvirus library identifies an ankyrin repeat protein involved in blocking induction of avian type I interferon. J. Virol. 2013, 87, 5041–5052. [Google Scholar] [CrossRef]
- Giotis, E.S.; Scott, A.; Rothwell, L.; Hu, T.; Talbot, R.; Todd, D.; Burt, D.W.; Glass, E.J.; Kaiser, P. Chicken anaemia virus evades host immune responses in transformed lymphocytes. J. Gen. Virol. 2018. [Google Scholar] [CrossRef]
- Kal, A.J.; van Zonneveld, A.J.; Benes, V.; van den Berg, M.; Koerkamp, M.G.; Albermann, K.; Strack, N.; Ruijter, J.M.; Richter, A.; Dujon, B.; et al. Dynamics of gene expression revealed by comparison of serial analysis of gene expression transcript profiles from yeast grown on two different carbon sources. Mol. Biol. Cell 1999, 10, 1859–1872. [Google Scholar] [CrossRef]
- Giotis, E.S.; Robey, R.C.; Skinner, N.G.; Tomlinson, C.D.; Goodbourn, S.; Skinner, M.A. Chicken interferome: Avian interferon-stimulated genes identified by microarray and RNA-seq of primary chick embryo fibroblasts treated with a chicken type I interferon (IFN-α). Vet. Res. 2016, 47, 75. [Google Scholar] [CrossRef]
- Vandepoele, K.; Saeys, Y.; Simillion, C.; Raes, J.; Van De Peer, Y. The automatic detection of homologous regions (ADHoRe) and its application to microcolinearity between Arabidopsis and rice. Genome Res. 2002, 12, 1792–1801. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Fuet, A.; Pain, B. Chicken induced pluripotent stem cells: Establishment and characterization. Methods Mol. Biol. 2017, 1650, 211–228. [Google Scholar] [PubMed]
- Allegrucci, C.; Young, L.E. Differences between human embryonic stem cell lines. Hum. Reprod. Update 2007, 13, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Giotis, E.S.; Ross, C.S.; Robey, R.C.; Nohturfft, A.; Goodbourn, S.; Skinner, M.A. Constitutively elevated levels of SOCS1 suppress innate responses in DF-1 immortalised chicken fibroblast cells. Sci. Rep. 2017, 7, 17485. [Google Scholar] [CrossRef] [PubMed]
- Ivashkiv, L.B.; Donlin, L.T. Regulation of type i interferon responses. Nat. Rev. Immunol. 2014, 14, 36–49. [Google Scholar] [CrossRef]
- Chambers, I.; Silva, J.; Colby, D.; Nichols, J.; Nijmeijer, B.; Robertson, M.; Vrana, J.; Jones, K.; Grotewold, L.; Smith, A. Nanog safeguards pluripotency and mediates germline development. Nature 2007, 450, 1230–1234. [Google Scholar] [CrossRef]
- Masuda, Y.; Matsuda, A.; Usui, T.; Sugai, T.; Asano, A.; Yamano, Y. Biological effects of chicken type III interferon on expression of interferon-stimulated genes in chickens: Comparison with type I and type II interferons. J. Vet. Med. Sci. 2012, 74, 1381–1386. [Google Scholar] [CrossRef]
- Qu, H.; Yang, L.; Meng, S.; Xu, L.; Bi, Y.; Jia, X.; Li, J.; Sun, L.; Liu, W. The differential antiviral activities of chicken interferon α (ChIFN-α) and ChIFN-β are related to distinct interferon-stimulated gene expression. PLoS ONE 2013, 8, e59307. [Google Scholar] [CrossRef]
- Karpala, A.J.; Lowenthal, J.W.; Bean, A.G. Identifying innate immune pathways of the chicken may lead to new antiviral therapies. Vet. Immunol. Immunopathol. 2012, 148, 100–109. [Google Scholar] [CrossRef]
- Liem, J.; Liu, J. Stress beyond translation: Poxviruses and more. Viruses 2016, 8, 169. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.J.; Iyer, S.; Sataur, A.; Covello, K.L.; Chodosh, L.A.; Simon, M.C. Differential regulation of the transcriptional activities of hypoxia-inducible factor 1 α (HIF-1α) and HIF-2α in stem cells. Mol. Cell. Biol. 2006, 26, 3514–3526. [Google Scholar] [CrossRef] [PubMed]
- Covello, K.L.; Kehler, J.; Yu, H.; Gordan, J.D.; Arsham, A.M.; Hu, C.J.; Labosky, P.A.; Simon, M.C.; Keith, B. Hif-2α regulates Oct-4: Effects of hypoxia on stem cell function, embryonic development, and tumor growth. Genes Dev. 2006, 20, 557–570. [Google Scholar] [CrossRef]
- Cameron, C.M.; Harding, F.; Hu, W.S.; Kaufman, D.S. Activation of hypoxic response in human embryonic stem cell-derived embryoid bodies. Exp. Biol. Med. 2008, 233, 1044–1057. [Google Scholar] [CrossRef] [PubMed]
- Xie, P. TRAF molecules in cell signaling and in human diseases. J. Mol. Signal 2013, 8, 7. [Google Scholar] [CrossRef]
- Dulwich, K.L.; Giotis, E.S.; Gray, A.; Nair, V.; Skinner, M.A.; Broadbent, A.J. Differential gene expression in chicken primary B cells infected ex vivo with attenuated and very virulent strains of infectious bursal disease virus (IBDV). J. Gen. Virol. 2017, 98, 2918–2930. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comparisons vs. Respective | # Upregulated Genes | # Downregulated Genes | ||
|---|---|---|---|---|
| Mock (FDR < 0.05) | ≥1.5 | ≥3 | ≥−1.5 | ≥−3 |
| chIFN-α 6 h | 534 | 278 | 412 | 238 |
| FP9 16 h | 98 | 32 | 116 | 39 |
| CNPV 16 h | 164 | 86 | 1403 | 693 |
| PBG98 16 h | 13 | 5 | 26 | 5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giotis, E.S.; Montillet, G.; Pain, B.; Skinner, M.A. Chicken Embryonic-Stem Cells Are Permissive to Poxvirus Recombinant Vaccine Vectors. Genes 2019, 10, 237. https://doi.org/10.3390/genes10030237
Giotis ES, Montillet G, Pain B, Skinner MA. Chicken Embryonic-Stem Cells Are Permissive to Poxvirus Recombinant Vaccine Vectors. Genes. 2019; 10(3):237. https://doi.org/10.3390/genes10030237
Chicago/Turabian StyleGiotis, Efstathios S., Guillaume Montillet, Bertrand Pain, and Michael A. Skinner. 2019. "Chicken Embryonic-Stem Cells Are Permissive to Poxvirus Recombinant Vaccine Vectors" Genes 10, no. 3: 237. https://doi.org/10.3390/genes10030237