Implications of Gene Inheritance Patterns on the Heterosis of Abdominal Fat Deposition in Chickens

 ,
, 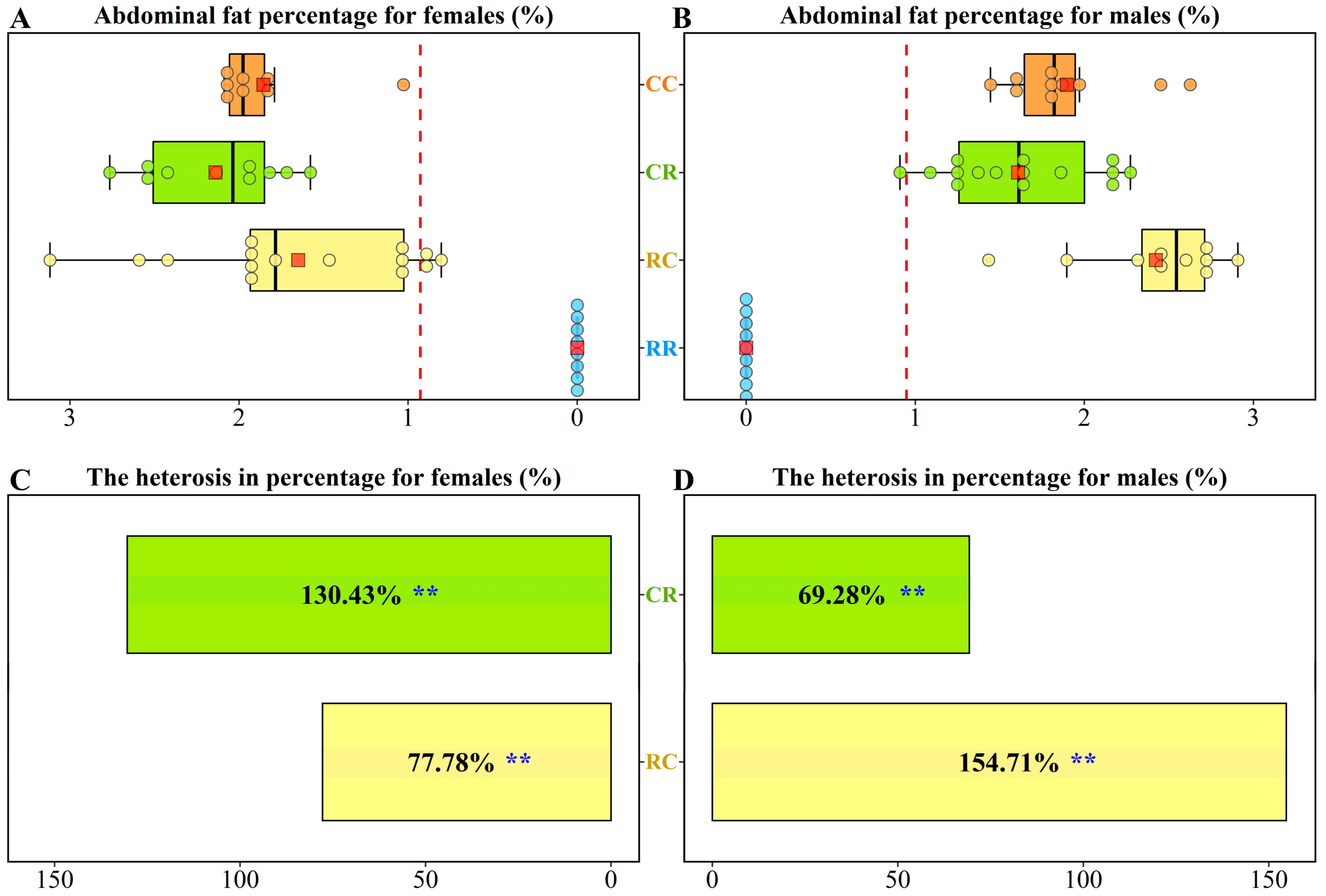{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Populations and Phenotypic Measurement
2.2. Sample Collection, RNA Extraction, and RNA Sequencing
2.3. Differentially Expressed Gene Identification
2.4. Inheritance Mode Classification
2.5. KEGG Pathway Analyses
2.6. Real-Time PCR Analysis
3. Results
3.1. Heterosis of Abdominal Fat Percentage in Chickens
3.2. Divergence of Gene Expression between Reciprocal Crosses and Parental Lines
3.3. Functional Analysis of Non-Additive Genes
3.4. Quantitative RT-PCR Validation for RNA Sequencing Data
3.5. Gene Expression Levels of Over-Dominant Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Birchler, J.A.; Yao, H.; Chudalayandi, S.; Vaiman, D.; Veitia, R. Heterosis. Plant Cell 2010, 22, 2105–2112. [Google Scholar] [CrossRef] [Green Version]
- Denning, G.; Kabambe, P.; Sanchez, P.; Malik, A.; Flor, R.; Harawa, R.; Nkhoma, P.; Zamba, C.; Banda, C.; Magombo, C.; et al. Input subsidies to improve smallholder maize productivity in Malawi: Toward an African green revolution. PLoS Biol. 2009, 7, e23. [Google Scholar] [CrossRef]
- Hara, H.; Hanzawa, K.; Yoshida, Y.; Watanabe, S. Characterization of a chicken × peahen intergeneric hybrid produced under natural mating. Hayvansal Uretim 2013, 54, 1–10. [Google Scholar]
- Springer, N.; Stupar, R. Allelic variation and heterosis in maize: How do two halves make more than a whole? Genome Res. 2007, 17, 264–275. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Ni, Z.; Yao, Y.; Nie, X.; Sun, Q. Gibberellins and heterosis of plant height in wheat (Triticum aestivum L.). BMC Genet. 2007, 8, 40. [Google Scholar] [CrossRef]
- Rahmathulla, V. Management of climatic factors for successful silkworm (Bombyx mori L.) crop and higher silk production: A review. Psyche J. Entomol. 2012, 2012, 1–12. [Google Scholar]
- Eeva, T.; Lehikoinen, E. Egg shell quality, cluth size and hatching success of the great tit (Parus major) and the pied flycatcher (Ficedula hypoleuca) in an air pollution gradient. Oecologia 1995, 102, 312–323. [Google Scholar] [CrossRef]
- Havenstein, G.; Ferket, P.; Qureshi, M. Growth, livability, and feed conversion of 1957 versus 2001 broilers when fed representative 1957 and 2001 broiler diets. Poult. Sci. 2003, 82, 1500–1508. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Wang, D.; Zhang, Y.; Yu, Y.; Xu, G.; Li, J. Differential gene expression in liver of inbred chickens and their hybrid offspring. Anim. Genet. 2005, 36, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Abasht, B.; Lamont, S. Genome-wide association analysis reveals cryptic alleles as an important factor in heterosis for fatness in chicken F2 population. Anim. Genet. 2007, 38, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, D.; Honaker, C.; Dorshorst, B.; Andersson, L.; Siegel, P. Asymmetries, heterosis, and phenotypic profiles of red junglefowl, White Plymouth Rocks, and F1 and F2 reciprocal crosses. J. Appl. Genet. 2018, 59, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Yan, W.; Sun, C.; Ji, C.; Zhou, Q.; Zhang, D.; Zheng, J.; Ning, Y. The gut microbiota is largely independent of host genetics in regulating fat deposition in chickens. ISME J. 2019, 13, 1422–1436. [Google Scholar] [CrossRef] [PubMed]
- Nayeri, S.; Stothard, P. Tissues, metabolic pathways and genes of key importance in lactating dairy cattle. Springer Sci. Rev. 2016, 2, 49–77. [Google Scholar] [CrossRef]
- Fouad, A.; El-Senousey, H. Nutritional factors affecting abdominal fat deposition in poultry: A Review. Asian Australas. J. Anim. Sci. 2014, 27, 1057–1068. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Lu, K.; Chen, Z.; Mu, T.; Hu, Z.; Li, X. Dominance, overdominance and epistasis condition the heterosis in two heterotic rice hybrids. Genetics 2008, 180, 1725–1742. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Guo, Z.; Liu, Z.; Cheng, Q.; Qu, X.; Chen, R.; Jiang, D.; Liu, C.; Wang, W.; Sun, Y.; et al. Global RNA sequencing reveals that genotype-dependent allele-specific expression contributes to differential expression in rice F1 hybrids. BMC Plant Biol. 2013, 13, 221. [Google Scholar] [CrossRef]
- Seymour, D.; Chae, E.; Grimm, D.; Martín Pizarro, C.; Habring-Müller, A.; Vasseur, F.; Rakitsch, B.; Borgwardt, K.; Koenig, D.; Weigel, D. Genetic architecture of nonadditive inheritance in Arabidopsis thaliana hybrids. Proc. Natl. Acad. Sci. USA 2016, 113, E7317–E7326. [Google Scholar] [CrossRef]
- Birchler, J.; Yao, H.; Chudalayandi, S. Unraveling the genetic basis of hybrid vigor. Proc. Natl. Acad. Sci. USA 2006, 103, 12957–12958. [Google Scholar] [CrossRef] [Green Version]
- Swanson-Wagner, R.; Jia, Y.; DeCook, R.; Borsuk, L.; Nettleton, D.; Schnable, P. All possible modes of gene action are observed in a global comparison of gene expression in a maize F1 hybrid and its inbred parents. Proc. Natl. Acad. Sci. USA 2006, 103, 6805–6810. [Google Scholar] [CrossRef]
- Gu, H.; Qi, X.; Jia, Y.; Zhang, Z.; Nie, C.; Li, X.; Li, J.; Jiang, Z.; Wang, Q.; Qu, L. Inheritance patterns of the transcriptome in hybrid chickens and their parents revealed by expression analysis. Sci. Rep. 2019, 9, 5750. [Google Scholar] [CrossRef]
- Zhuo, Z.; Lamont, S.; Abasht, B. RNA-Seq analyses identify additivity as the predominant gene expression pattern in F1 chicken embryonic brain and liver. Genes 2019, 10, E27. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhang, C. Heterosis and statistical tests. Hereditas 1983, 5, 24–26. [Google Scholar]
- Kim, D.; Langmead, B.; Salzberg, S. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.; Antonescu, C.M.; Chang, T.; Mendell, J.; Salzberg, S. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Love, M.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Rapp, R.; Udall, J.; Wendel, J. Genomic expression dominance in allopolyploids. BMC Biol. 2009, 7, 18. [Google Scholar] [CrossRef]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.; Luciani, A.; Potter, S.; Qureshi, M.; Richardson, L.; Salazar, G.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.; Han, Y.; He, Q. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Schoenmakers, S.; Wassenaar, E.; Hoogerbrugge, J.; Laven, J.; Grootegoed, J.; Baarends, W. Female meiotic sex chromosome inactivation in chicken. PLoS Genet. 2009, 5, e1000466. [Google Scholar] [CrossRef]
- Wu, X.; Li, R.; Li, Q.; Bao, H.; Wu, C. Comparative transcriptome analysis among parental inbred and crosses reveals the role of dominance gene expression in heterosis in Drosophila melanogaster. Sci. Rep. 2016, 6, 21124. [Google Scholar] [CrossRef]
- Wang, H.; Fang, Y.; Wang, L.; Zhu, W.; Ji, H.; Wang, H.; Xu, S.; Sima, Y. Heterosis and differential gene expression in hybrids and parents in Bombyx mori by digital gene expression profiling. Sci. Rep. 2016, 6, 21124. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, J.; He, T.; Becker, S.; Zhang, G.; Li, D.; Ma, X. Butyrate: A double-edged sword for health? Adv. Nutr. 2018, 9, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Ajuwon, K. Mechanism of butyrate stimulation of triglyceride storage and adipokine expression during adipogenic differentiation of porcine stromovascular cells. PLoS ONE 2015, 10, e145940. [Google Scholar] [CrossRef] [PubMed]
- Thomas, L.; Ittmann, M.; Cooper, C. The role of leucine in ketogenesis in starved rats. Biochem. J. 1982, 204, 399–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puchalska, P.; Crawford, P. Multi-dimensional roles of ketone bodies in fuel metabolism, signaling, and therapeutics. Cell Metab. 2017, 25, 262–284. [Google Scholar] [CrossRef]
- Geelen, M.; Lopes-Cardozo, M.; Edmond, J. Acetoacetate: A major substrate for the synthesis of cholesterol and fatty acids by isolated rat hepatocytes. FEBS Lett. 1983, 163, 269–273. [Google Scholar] [CrossRef]
- Freed, L.; Endemann, G.; Tomera, J.; Gavino, V.; Brunengraber, H. Lipogenesis from ketone bodies in perfused livers from streptozocin-induced diabetic rats. Diabetes 1988, 37, 50–55. [Google Scholar] [CrossRef]
- Du, Y.; Meng, Q.; Zhang, Q.; Guo, F. Isoleucine or valine deprivation stimulates fat loss via increasing energy expenditure and regulating lipid metabolism in WAT. Amino Acids 2012, 43, 725–734. [Google Scholar] [CrossRef]
- Halama, A.; Horsch, M.; Kastenmüller, G.; Möller, G.; Kumar, P.; Prehn, C.; Laumen, H.; Hauner, H.; Hrabĕde Angelis, M.; Beckers, J.; et al. Metabolic switch during adipogenesis: From branched chain amino acid catabolism to lipid synthesis. Arch. Biochem. Biophys. 2016, 589, 93–107. [Google Scholar] [CrossRef] [Green Version]
- Maltest, W.; Reitz, B.; Volpe, J. Effects of isoleucine deprivation on synthesis of sterols and fatty acids in LM-cells. J. Biol. Chem. 1981, 256, 2185–2193. [Google Scholar]
- Alam, M.; Rahman, M. Mitochondrial dysfunction in obesity: Potential benefit and mechanism of Co-enzyme Q10 supplementation in metabolic syndrome. J. Diabetes Metab. Disord. 2014, 13, 60. [Google Scholar] [CrossRef] [PubMed]
- Ruby, M.; Massart, J.; Hunerdosse, D.; Schönke, M.; Correia, J.; Louie, S.; Ruas, J.; Näslund, E.; Nomura, D.; Zierath, J. Human carboxylesterase 2 reverses obesity-induced diacylglycerol accumulation and glucose intolerance. Cell Rep. 2017, 18, 636–646. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.; Taylor, A.; Tong, E.; Repa, J. Carboxylesterases are uniquely expressed among tissues and regulated by nuclear hormone receptors in the mouse. Drug Metab. Dispos. 2012, 41, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Howie, D.; Cobbold, S.; Adams, E.; Ten, B.; Necula, A.; Zhang, W.; Huang, H.; Roberts, D.; Thomas, B.; Hester, S.; et al. Foxp3 drives oxidative phosphorylation and protection from lipotoxicity. JCI Insight 2017, 2, e89160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golson, M.; Kaestner, K. Fox transcription factors: From development to disease. Development 2016, 143, 4558–4570. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.; Kang, N.; Koh, H.; Shin, S.; Lee, B.; Jeong, B.; Chang, Y. Enhancement of biomass and lipid productivity by overexpression of a bZIP transcription factor in Nannochloropsis salina. Biotechnol. Bioeng. 2018, 115, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, Y.; Shin, S.; Choi, B.; Kim, H.; Jang, S.; Kajikawa, M.; Yamano, T.; Kong, F.; Légeret, B.; Fukuzawa, H.; et al. The bZIP1 transcription factor regulates lipid remodeling and contributes to ER stress management in Chlamydomonas reinhardtii. Plant Cell. 2019, 31, 1127–1140. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Balamurugan, S.; Yang, Y.; Zheng, J.; Huang, D.; Zou, L.; Yang, W.; Liu, J.; Guan, Y.; Li, H. Transcriptional regulation of microalgae for concurrent lipid overproduction and secretion. Sci. Adv. 2019, 5. [Google Scholar] [CrossRef]
- Sension, M.; Deckx, H. Lipid metabolism and lipodystrophy in HIV-1-infected patients: The role played by nonnucleoside reverse transcriptase inhibitors. AIDS Rev. 2015, 17, 21–36. [Google Scholar]
- Luo, W.; Qin, L.; Li, B.; Liao, Z.; Liang, J.; Xiao, X.; Xiao, X.; Mo, Y.; Huang, G.; Zhang, Z.; et al. Inactivation of HMGCL promotes proliferation and metastasis of nasopharyngeal carcinoma by suppressing oxidative stress. Sci. Rep. 2017, 7, 11954. [Google Scholar] [CrossRef]
- Lv, Z.; Fan, H.; Song, B.; Li, G.; Liu, D.; Guo, Y. Supplementing genistein for breeder hens alters the fatty acid metabolism and growth performance of offsprings by epigenetic modification. Oxid. Med. Cell. Longev. 2019, 2019, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Cui, X.; Liu, R.; Li, Q.; Zheng, M.; Zhao, G.; Ge, C.; Wen, J.; Hu, Y.; Cui, H. Identification of differentially expressed genes and pathways for abdominal fat deposition in ovariectomized and sham-operated chickens. Genes 2019, 10, E155. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Yan, W.; Zheng, J.; Ji, C.; Zhang, D.; Sun, C.; Yang, N. Feed efficiency measures and their relationships with production and meat quality traits in slower growing broilers. Poult. Sci. 2018, 97, 2356–2364. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mai, C.; Wen, C.; Sun, C.; Xu, Z.; Chen, S.; Yang, N. Implications of Gene Inheritance Patterns on the Heterosis of Abdominal Fat Deposition in Chickens. Genes 2019, 10, 824. https://doi.org/10.3390/genes10100824
Mai C, Wen C, Sun C, Xu Z, Chen S, Yang N. Implications of Gene Inheritance Patterns on the Heterosis of Abdominal Fat Deposition in Chickens. Genes. 2019; 10(10):824. https://doi.org/10.3390/genes10100824
Chicago/Turabian StyleMai, Chunning, Chaoliang Wen, Congjiao Sun, Zhiyuan Xu, Sirui Chen, and Ning Yang. 2019. "Implications of Gene Inheritance Patterns on the Heterosis of Abdominal Fat Deposition in Chickens" Genes 10, no. 10: 824. https://doi.org/10.3390/genes10100824