Ivermectin as a Broad-Spectrum Host-Directed Antiviral: The Real Deal?


Abstract
:1. Introduction
2. Ivermectin as a US Food and Drug Administration-Approved Anti-Parasitic Agent
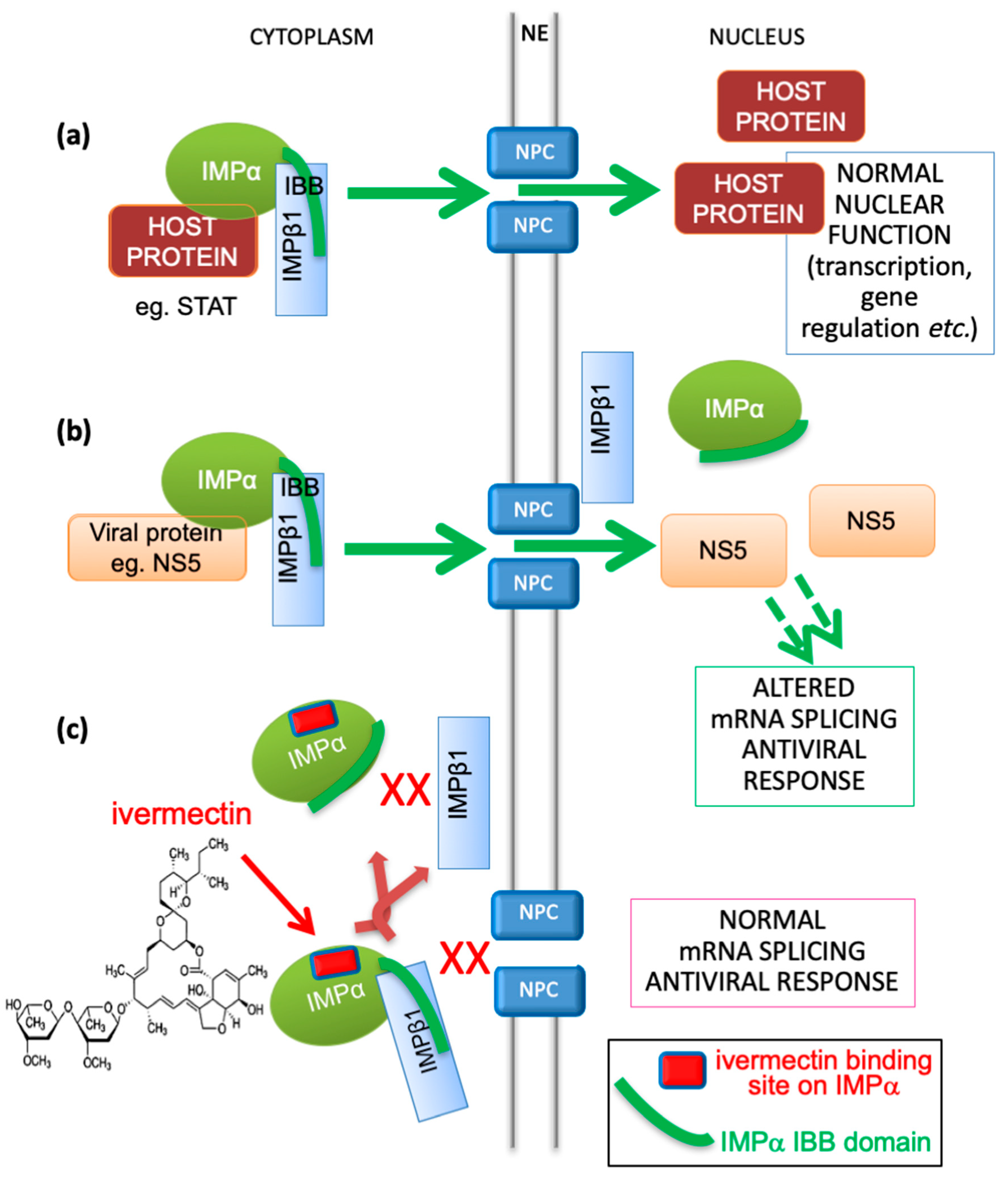
3. Ivermectin as an IMPα Targeting Agent with Antiviral Activity
4. Ivermectin as an Antiviral
5. Ivermectin as an Antiviral in the Clinic
6. A Viable Treatment for SARS-CoV-2?
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Crump, A.; Omura, S. Ivermectin, wonder drug from Japan: The human use perspective. Proc. Jpn. Acad. 2011, 87, 13–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobel Foundation. The Nobel Prize in Medicine or Physiology. 2015. Available online: https://www.nobelprize.org/prizes/medicine/2015/press-release (accessed on 11 September 2020).
- González Canga, A.; Sahagún Prieto, A.M.; Diez Liébana, M.J.; Fernández Martínez, N.; Sierra Vega, M.; García Vieitez, J.J. The pharmacokinetics and interactions of ivermectin in humans—A mini-review. AAPSJ 2008, 10, 42–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crump, A.; Omura, S. Ivermectin: Panacea for resource-poor communities? Trends Parasitol. 2014, 30, 445–455. [Google Scholar]
- World Health Organization. World Health Organization’s List of Essential Medicines 21st List 2019. Available online: https://apps.who.int/iris/bitstream/handle/10665/325771/WHO-MVP-EMP-IAU-2019.06-eng.pdf (accessed on 11 September 2020).
- Kumar, B.S.; Jeyaraman, M.; Jain, R.; Anudeep, T.C. A Wonder Drug in the Arsenal against COVID—19: Medication Evidence from Ivermectin. J. Adv. Med. Med. Res. 2020, 32, 30–37. [Google Scholar] [CrossRef]
- Wagstaff, K.M.; Sivakumaran, H.; Heaton, S.; Harrich, D.; Jans, D.A. Ivermectin is a specific inhibitor of importin α/β-mediated nuclear import able to inhibit replication of HIV-1 and dengue virus. Biochem. J. 2012, 443, 851–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tay, M.Y.; Fraser, J.E.; Chan, W.K.K.; Moreland, N.J.; Rathore, A.P.; Wang, C.; Vasudevan, S.G.; Jans, D.A. Nuclear localization of dengue virus (DENV) 1-4 non-structural protein 5; protection against all 4 DENV serotypes by the inhibitor Ivermectin. Antivir. Res. 2013, 99, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, L.; Pinkhan, C.; Baer, A.; Amaya, M.; Narayan, A.; Wagstaff, K.M.; Jans, D.A.; Kehn-Hall, K. Nuclear import and export inhibitors alter capsid protein distribution in mammalian cells and reduce Venezuelan Equine Encephalitis Virus replication. Antivir. Res. 2013, 100, 662–672. [Google Scholar] [CrossRef]
- Gotz, V.; Magar, L.; Dornfeld, D.; Giese, S.; Pohlmann, A.; Höper, D.; Kong, B.-W.; Jans, D.A.; Beer, M.; Haller, O.; et al. Influenza A viruses escape from MxA restriction at the expense of efficient nuclear vRNP import. Sci. Rep. 2016, 6, 23138. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.N.Y.; Atkinson, S.C.; Wang, C.; Lee, A.; Bogoyevitch, M.A.; Borg, N.A.; Jans, D.A. The broad spectrum antiviral ivermectin targets the host nuclear transport importin α/β1 heterodimer. Antivir. Res. 2020, 177, 104760. [Google Scholar] [CrossRef]
- Mastrangelo, E.; Pezzullo, M.; De Burghgraeve, T.; Kaptein, S.; Pastorino, B.; Dallmeier, K.; de Lamballerie, X.; Neyts, J.; Hanson, A.M.; Frick, D.N.; et al. Lvermectin is a potent inhibitor of flavivirus replication specifically targeting NS3 helicase activity: New prospects for an old drug. J. Antimicrob. Chemother. 2012, 67, 1884–1894. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, S.; Audsley, M.; Lieu, K.; Marsh, G.; Thomas, D.R.; Heaton, S.; Paxman, J.; Wagstaff, K.M.; Buckle, A.G.; Moseley, G.; et al. Recognition by host nuclear transport proteins drives disorder-to-order transition in Hendra virus V. Sci. Rep. 2018, 8, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jans, D.A.; Martin, A.J.; Wagstaff, K.M. Inhibitors of nuclear transport. Curr. Opin. Cell Biol. 2019, 58, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Varghese, F.S.; Kaukinen, P.; Glaesker, S.; Bespalov, M.; Hanski, L.; Wennerberg, K.; Kümmerer, B.M.; Ahola, T. Discovery of berberine, abamectin and ivermectin as antivirals against chikungunya and other alphaviruses. Antivir. Res. 2016, 126, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.R.; Lundberg, L.; Pinkhan, C.; Shechter, S.; Debono, A.; Baell, J.; Wagstaff, K.M.; Hick, C.A.; Kehn-Hall, K.; Jans, D.A. Identification of novel antivirals inhibiting recognition of Venezuelan equine encephalitis virus capsid protein by the Importin α/β1 heterodimer through high-throughput screening. Antivir. Res. 2018, 151, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Caly, L.; Druce, J.D.; Catton, M.G.; Jans, D.A.; Wagstaff, K.M. The FDA-approved drug ivermectin inhibits the replication of SARS-CoV-2 in vitro. Antivir. Res. 2020, 178, 104787. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Liu, W.; Wang, B.; Dang, R.; Qiu, L.; Ren, J.; Yan, C.; Yang, Z.; Wang, X. Ivermectin inhibits DNA polymerase UL42 of Pseudorabies virus entrance into the nucleus and proliferation of the virus in vitro and vivo. Antivir. Res. 2018, 159, 55–62. [Google Scholar] [CrossRef]
- Bennett, S.M.; Zhao, L.; Bosard, C.; Imperiale, M.J. Role of a nuclear localization signal on the minor capsid proteins VP2 and VP3 in BKPyV nuclear entry. Virology 2015, 474, 110–116. [Google Scholar] [CrossRef] [Green Version]
- King, C.R.; Tessier, T.M.; Dodge, M.J.; Weinberg, J.B.; Mymryk, J.S. Lnhibition of human adenovirus replication by the importin α/β1 nuclear import inhibitor ivermectin. J. Virol. 2020, in press. [Google Scholar] [CrossRef]
- Changeux, J.-P.; Amoura, Z.; Rey, F.A.; Miyara, M. A nicotinic hypothesis for Covid-19 with preventive and therapeutic implications. Comptes Rendus Biol. 2020, 343, 33–39. [Google Scholar] [CrossRef]
- Krause, R.M.; Buisson, B.; Bertrand, S.; Corringer, P.J.; Galzi, J.L.; Changeux., J.P.; Bertrand, D. Lvermectin: A positive allosteric effector of the alpha7 neuronal nicotinic acetylcholine receptor. Mol. Pharmacol. 1998, 53, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Guzzo, C.A.; Furtek, C.I.; Porras, A.G.; Chen, C.; Tipping, R.; Clineschmidt, C.M.; Sciberras, D.G.; Hsieh, J.Y.K.; Lasseter, K.C. Safety, tolerability, and pharmacokinetics of escalating high doses of ivermectin in healthy adult subjects. J. Clin. Pharmacol. 2002, 42, 1122–1133. [Google Scholar] [CrossRef] [PubMed]
- Loukas, A.; Hotez, P.J. Chemotherapy of helminth infections. In Goodman & Gilman’s The Pharmacological Basis of Therapeutics, 11th ed.; Brunton, L.L., Lazo, J.S., Parker, K.L., Eds.; McGraw Hill: New York, NY, USA, 2006; pp. 1073–1093. [Google Scholar]
- Navarro, M.; Camprubí, D.; Requena-Méndez, A.; Buonfrate, D.; Giorli, G.; Kamgno, J.; Gardon, J.; Muñoz, J.; Krolewiecki, A. Safety of high-dose ivermectin: A systematic review and meta-analysis. J. Antimicrob. Chemother. 2020, 75, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Twum-Danso, N.A.Y.; Meredith, S.E.O. Variation in incidence of serious adverse events after onchocerciasis treatment with ivermectin in areas of Cameroon co-endemic for loiasis. Trop. Med. Int. Health 2003, 8, 820–831. [Google Scholar] [CrossRef]
- Fulcher, A.; Jans, D.A. Regulation of nucleocytoplasmic trafficking of viral proteins; an integral role in pathogenesis ? Biochem. Biophys. Acta Mol. Cell Res. 2011, 1813, 2176–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caly, L.; Wagstaff, K.M.; Jans, D.A. Nuclear trafficking of proteins from RNA viruses: Potential target for anti-virals? Antivir. Res. 2012, 95, 202–206. [Google Scholar] [CrossRef]
- Yang, S.N.Y.; Atkinson, S.C.; Fraser, J.E.; Wang, C.; Maher, B.; Roman, N.; Forwood, J.K.; Wagstaff, K.M.; Borg, N.A.; Jans, D.A. Novel Flavivirus Antiviral That Targets The Host Nuclear Transport Importin α/β1 Heterodimer. Cells 2019, 8, 281. [Google Scholar] [CrossRef] [Green Version]
- Pryor, M.J.; Rawlinson, S.M.; Butcher, R.E.; Barton, C.L.; Waterhouse, T.A.; Vasudevan, S.G.; Bardin, P.G.; Wright, P.J.; Jans, D.A.; Davidson, A.D. Nuclear localization of dengue virus nonstructural protein 5 through its importin alpha/beta-recognized nuclear localization sequences is integral to viral infection. Traffic 2007, 8, 795–807. [Google Scholar] [CrossRef] [PubMed]
- Fraser, J.E.; Watanabe, S.; Wang, C.; Chan, W.K.; Maher, B.; Lopez-Denman, A.; Hick, C.; Wagstaff, K.M.; Mackenzie, J.M.; Sexton, P.M.; et al. A nuclear transport inhibitor that modulates the unfolded protein response and provides in vivo protection against lethal dengue virus infection. J. Infect. Dis. 2014, 210, 1780–1791. [Google Scholar] [CrossRef] [Green Version]
- Frieman, M.; Yount, B.; Heise, M.; Kopecky-Bromberg, S.A.; Palese, P.; Baric, R.S. Severe Acute Respiratory Syndrome Coronavirus ORF6 Antagonizes STAT1 Function by Sequestering Nuclear Import Factors on the Rough Endoplasmic Reticulum/Golgi Membrane. J. Virol. 2007, 81, 9812–9824. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Ye1, F.; Zhu, N.; Wang, W.; Deng, Y.; Zhao, Z.; Tan, W. Middle East respiratory syndrome coronavirus ORF4b protein inhibits type I interferon production through both cytoplasmic and nuclear targets. Sci. Rep. 2015, 5, 17554. [Google Scholar] [CrossRef] [Green Version]
- Wagstaff, K.M.; Rawlinson, S.M.; Hearps, A.C.; Jans, D.A. An AlphaScreen(R)-based assay for high-throughput screening for specific inhibitors of nuclear import. J. Biomol. Screen. 2011, 16, 192–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagstaff, K.M.; Headey, S.; Telwatte, S.; Tyssen, D.; Hearps, A.C.; Thomas, D.R.; Tachedjian, G.; Jans, D.A. Molecular dissection of an inhibitor targeting the HIV integrase dependent preintegration complex nuclear import. Cell. Microbiol. 2018, e12953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Denman, A.J.; Russo, A.; Wagstaff, K.M.; White, P.A.; Jans, D.A.; Mackenzie, J.M. Nucleocytoplasmic shuttling of the West Nile virus RNA-dependent RNA polymerase NS5 is critical to infection. Cell. Microbiol. 2018, 20, e12848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ci, X.; Li, H.; Yu, Q.; Zhang, X.; Yu, L.; Chen, N.; Song, Y.; Deng, X. Avermectin exerts anti-inflammatory effect by downregulating the nuclear transcription factor kappa-B and mitogen-activated protein kinase activation pathway. Fundam. Clin. Pharmacol. 2009, 23, 449–455. [Google Scholar] [CrossRef]
- Yu, Z.-H.; Chan, H.C. Gossypol as a male antifertility agent—Why studies should have been continued. Int. J. Androl. 1998, 21, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Porat, O. Effects of Gossypol on the Motility of Mammalian Sperm. Mol. Reprod. Dev. 1990, 21, 400–408. [Google Scholar] [CrossRef]
- Yamasmith, E.; Saleh-arong, F.A.; Avirutnan, P.; Angkasekwinai, N.; Mairiang, D.; Wongsawat, E.; Tanrumluk, S.; Fongsri, U.; Suputtamongkol, Y. Efficacy and Safety of Ivermectin against Dengue Infection: A Phase III, Randomized, Double-blind, Placebo-controlled Trial. In Proceedings of the 34th Annual Meeting The Royal College of Physicians of Thailand—‘Internal Medicine and One Health’, Pattaya, Thailand, 26–28 April 2018. [Google Scholar]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef]
- Brady, O.J.; Gething, P.W.; Bhatt, S.; Messina, J.P.; Brownstein, J.S.; Hoen, A.G.; Moyes, C.L.; Farlow, A.W.; Scott, T.W.; Hay, S.I. Refining the Global Spatial Limits of Dengue Virus Transmission by Evidence-Based Consensus. PLoS Negl. Trop. Dis. 2012, 6, e1760. [Google Scholar] [CrossRef]
- Boulware, D.R.; Pullen, M.F.; Bangdiwala, A.S.; Pastick, K.A.; Lofgren, S.M.; Okafor, E.C.; Skipper, C.P.; Nascene, A.A.; Nicol, M.R.; Abassi, M.; et al. A Randomized Trial of Hydroxychloroquine as Postexposure Prophylaxis for Covid-19. N. Engl. J. Med. 2020, 383, 517–525. [Google Scholar] [CrossRef]
- US Food and Drug Administration. Coronavirus (COVID-19) Update: FDA Revokes Emergency Use Authorization for Chloroquine and Hydroxychloroquine. 15 June 2020. Available online: https://www.fda.gov/media/138945 (accessed on 11 September 2020).
- US Food and Drug Administration. FDA Cautions against Use of Hydroxychloroquine or Chloroquine for COVID-19 Outside of the Hospital Setting or a Clinical Trial due to Risk of Heart Rhythm Problems. 1 July 2020. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/nda/2020/OSE%20Review_Hydroxychloroquine-Cholorquine%20-%2019May2020_Redacted.pdf (accessed on 11 September 2020).
- Cao, B.; Wang, Y.; Wen, D.; Liu, W.; Wang, J.; Fan, G.; Ruan, R.; Song, B.; Cai, C.; Wei, M.; et al. A Trial of Lopinavir–Ritonavir in Adults Hospitalized with Severe Covid-19. N. Engl. J. Med. 2020, 382, 1787–1799. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, D.; Du, G.; Du, R.; Zhao, J.; Jin, Y.; Fu, S.; Gao, L.; Cheng, Z.; Lu, Q.; et al. Remdesivir in adults with severe COVID-19: A randomised, double-blind, placebo-controlled, multicentre trial. Lancet 2020, 395, 1569–1578. [Google Scholar] [CrossRef]
- Arshad, U.; Pertinez, H.; Box, H.; Tatham, L.; Rajoli, R.K.R.; Curley, P.; Neary, M.; Sharp, J.; Liptrott, N.J.; Valentijn, A.; et al. Prioritisation of Anti-SARS-Cov-2 Drug Repurposing Opportunities Based on Plasma and Target Site Concentrations Derived From Their Established Human Pharmacokinetics. Clin. Pharm. Ther. 2020. [Google Scholar] [CrossRef] [PubMed]
- Schmith, V.D.; Zhou, J.J.; Lohmer, L.R. The Approved Dose of Ivermectin Alone is not the Ideal Dose for the Treatment of COVID-19. Clin. Pharm. Ther. 2020. [Google Scholar] [CrossRef] [PubMed]
- Trial Site News. Eurnekian Public Hospital Argentina’s IVERCAR Ivermectin & Carrageenan Study Shows Positive Results Targeting COVID-19. 3 September 2020. Available online: https://www.trialsitenews.com/argentinas-ivercar-ivermectin-carrageenan-study-shows-positive-results-targeting-covid-19/ (accessed on 11 September 2020).
- Alam, M.; Murshed, R.; Bhiuyan, E.; Saber, S.; Alam, R.F.; Robin, R.C. A Case Series of 100 COVID-19 Positive Patients Treated with Combination of Ivermectin and Doxycycline. Bangladesh Coll. Phys. Surg. 2020, 38, 10–15. [Google Scholar] [CrossRef]
- Rahman, M.A.; Iqbal, S.A.; Islam, M.A.; Niaz, M.K.; Hussain, T.; Siddiquee, T.H. Comparison of Viral Clearance between Ivermectin with Doxycycline and Hydroxychloroquine with Azithromycin in COVID-19 Patients. Bangladesh Coll. Phys. Surg. 2020, 38, 5–9. [Google Scholar] [CrossRef]
- Republica Del Peru Ministerio De Salud. RM_270-2020-MINSA. 8 May 2020. Available online: https://cdn.www.gob.pe/uploads/document/file/694719/RM_270-2020-MINSA.PDF (accessed on 11 September 2020).
- The Gobierno del Estate Plurinacional de Bolivia Misterio de Salud. Ministerial Resolution No 259 from the Gobierno del Estate Plurinacional de Bolivia Misterio de Salud. 20 May 2020. Available online: https://www.minsalud.gob.bo/component/jdownloads/send/27-comunicado-oficial/425-resolucion-ministerial-n-0259 (accessed on 11 September 2020).


{kind=link}
| Compound | Documented Action on IMPα 3 | Antiviral Against 4 | Inhibitory Concentration (Assay)/Fold reduction 5 |
|---|---|---|---|
| Ivermectin 1 | Coronavirus SARS-CoV-2 HIV-1 (VSV-G-pseudotyped NL4-3.Luc.R-E-HIV) Influenza VLPs (avian influenza A/MxA escape mutants) Flavivirus: YFV (17D) DENV1 (EDEN-1) DENV2 (NGC) DENV2 (EDEN-2) DENV3 (EDEN-3) DENV4 (EDEN-4) WNV (NY99) WNV (MRM61C) ZIKV (Asian/Cook Islands/2014) Alphavirus: Chikungunya virus (CHIKV-Rluc) Sindbis (HR) Semliki forest virus VEEV (TC83) Hendra (Hendra virus/Australia/Horse/1994) DNA viruses Adenovirus (HAdV-C5) Adenovirus (HAdV-B3) BK polyomavirus (BKPyV) Pseudorabies | EC50 = 2.2/2.8 μM (qPCR/released/cell-associated virus) [17] 5000-fold 50 μM > 2-fold (luciferase) [7] 10 μM total inhibition (luciferase) [10] EC50 = 5/0.5 nM (CPE/qPCR) [12] 3 μM > 50,000-fold (pfu) [15] EC50 = 2.3/3 μM (CFI, 2 hosts) [8] EC50 = 0.7 μM (qPCR) [12] EC50 = 0.4/0.6 μM (pfu/qPCR) [11] 50 μM total inhibition (pfu) [7] EC50 = 2.1/1.7 μM (CFI, 2 hosts) [8] EC50 = 1.7 μM (CFI) [8] EC50 = 1.9 μM (CFI) [8] EC50 = 4 μM (qPCR) [12] EC50 = 1/0.5 μM (pfu/qPCR) [11] EC50 = 1.3/1.6 μM (pfu/qPCR) [11] EC50 = 1.9/0.6 μM (luciferase, 2 hosts) 3 μM > 5000-fold (pfu) [15] 3 μM > 1000-fold (pfu) [15] 3 μM > 200-fold (pfu) [15] 1 μM c. 20-fold (pfu) [9] est. EC50 = 2 μM (TCID/luciferase) [13] EC50 = c. 2.5 μM; 10 μM 20-fold (qPCR) [20] 10 μM c. 8-fold (qPCR) [20] Est. EC50 1.5 μM (PFU/CPE/qPCR) [19] Est. EC50 c. 0.8 μM 1000-fold [18] | |
| Gossypol 2 | WNV (MRM61C) Hendra (Hendra virus/Australia/Horse/1994) | 10 μM 100-fold (pfu) [36] 10 μM (TCID/luciferase) [14] | |
| GW5074 |
| DENV2 NGC ZIKV (Asian/Cook Islands/2014) WNV (MRM61C) | EC50 = 0.5/1 (pfu/PCR) [29] EC50 = 0.3/0.6 (pfu/PCR) [29] EC50 = 5/7 (pfu/PCR) [29] |
| Title, URL | Status 1 | N 2 | Interventions 3 | Start | Locations | |
|---|---|---|---|---|---|---|
| 1 | Ivermectin Effect on SARS-CoV-2 Replication in Patients With COVID-19; https://ClinicalTrials.gov/show/NCT04381884 | R | 45 | Ivermectin 0.6 mg/kg QD plus SC vs. SC | 18.5.20 | CEMIC, Buenos Aires, Ciudad De Buenos Aires, Argentina |
| 2 | Ivermectin and Nitazoxanide Combination Therapy for COVID-19; https://ClinicalTrials.gov/show/NCT04360356 | NY | 100 | Ivermectin 0.2 mg/kg once plus NZX 500 mg BID for 6 days vs. SC | 20.5.20 | Tanta University, Egypt |
| 3 | Ivermectin vs. Placebo for the Treatment of Patients With Mild to Moderate COVID-19; https://ClinicalTrials.gov/show/NCT04429711 | R | 100 | Ivermectin 12–15 mg/day for 3 days vs. Placebo | 12.5.20 | Sheba Medical Center, Ramat-Gan, Israel |
| 4 | Hydroxychloroquine and Ivermectin for the Treatment of COVID-19 Infection; https://ClinicalTrials.gov/show/NCT04391127 | R | 200 | Ivermectin 12 mg (<80 kg) or 18 mg (>80 kg) once vs. HCQ 400 mg BID for 1 day then 200 mg BID for 4 days vs. Placebo | 4.5.20 | Jose Manuel Arreola Guerra, Aguascalientes, Mexico |
| 5 | Efficacy of Ivermectin in Adult Patients With Early Stages of COVID-19. https://ClinicalTrials.gov/show/NCT04405843 | NY | 400 | Ivermectin 0.3 mg/kg daily for 5 days vs. Placebo | 20.6.20 | Colombia |
| 6 | Ivermectin In Treatment of COVID 19 Patients. https://ClinicalTrials.gov/show/NCT04425707 | R | 100 | Ivermectin (dose unlisted) vs. SC vs. Ivermectin (dose unlisted) plus SC | 9.6.20 | Isolation and referral hospitals for COVID 19 patients, Cairo, Egypt |
| 7 | Efficacy and Safety of Ivermectin and Doxycycline in Combination or IVE Alone in Patients With COVID-19 Infection; https://ClinicalTrials.gov/show/NCT04407130 | E | 72 | Ivermectin 0.2 mg/kg once plus 200 mg DOC day 1 followed by 100 mg DOC BID for 4 days vs. Ivermectin 0.2 mg/kg QD for 5 days vs. Placebo | 16.6.20 | Icddr, B, Dhaka, Bangladesh |
| 8 | Efficacy of Ivermectin as Add on Therapy in COVID19 Patients. https://ClinicalTrials.gov/show/NCT04343092 | C | 100 | Ivermectin 0.2 mg/kg once weekly plus HCQ 400 mg QD plus ATM 500 mg QD vs. HCQ 400 mg QD plus ATM 500 mg QD | 18.4.20 | General Directorate of Medical City, Bagdad, Baghdad, Iraq |
| 9 | COVidIVERmectin: Ivermectin for Treatment of Covid-19 (COVER). https://ClinicalTrials.gov/show/NCT04438850 | NY | 102 | Ivermectin 0.6 mg/kg QD for 5 days vs. Ivermectin 1.2 mg/kg QD for 5 days vs. Placebo | 20.6.20 | Negrar, Verona, Italy; Bologna, Italy; Milan, Italy; Rovereto, Italy; Turin, Italy; Barcelona, Spain; Madrid, Spain |
| 10 | Efficacy, Safety and Tolerability of Ivermectin in Subjects Infected With SARS-CoV-2 With or Without Symptoms (SILVERBULLET). https://ClinicalTrials.gov/show/NCT04407507 | NY | 66 | Ivermectin 12 mg/day for 3 days plus paracetamol 500 mg QID for 14 days vs. Placebo plus paracetamol 500 mg QID for 14 days | 20.6.20 | Investigacion Biomedica para el Desarrollo de Farmacos S.A. de C.V., Mexico |
| 11 | Sars-CoV-2/COVID-19 Ivermectin Navarra-ISGlobal Trial (SAINT). https://ClinicalTrials.gov/show/NCT04390022 | R | 24 | Ivermectin 0.4 mg/kg once vs. Placebo | 14.5.20 | Clinica Universidad de Navarra, Pamplona, Navarra, Spain |
| 12 | A Comparative Study on Ivermectin and Hydroxychloroquine on the COVID19 Patients in Bangladesh. https://ClinicalTrials.gov/show/NCT04434144 | C | 116 | Ivermectin 0.2 mg/kg once plus DOC 100 mg BID for 10 days vs. HCQ 400 mg day 1 then 200 mg BID for 9 days plus ATM 500 mg/day for 5 days | 2.5.20 | Chakoria Upazilla Health Complex, Cox’s Bazar, Bangladesh |
| 13 | Ivermectin vs Combined Hydroxychloroquine and Antiretroviral Drugs (ART) Among Asymptomatic COVID-19 Infection (IDRA-COVID19). https://ClinicalTrials.gov/show/NCT04435587 | NY | 80 | Ivermectin 0,6 mg/kg daily for 3 days vs. HCQ 400 mg BID Day 1 then 200 mg BID for 4 days plus Darunavir/ritonavir (400 mg/100 mg) BID for 5 days | 20.7.20 | Siriraj Hospital, Bangkok Noi, Bangkok, Thailand |
| 14 | IVERMECTIN Aspirin Dexametasone and Enoxaparin as Treatment of Covid 19. https://ClinicalTrials.gov/show/NCT04425863 | A | 100 | Ivermectin 5 mg/mL oral to be repeated 1 week later (dose unlisted) | 1.5.20 | Hospital Eurnekian, Buenos Aires, Argentina |
| 15 | A Preventive Treatment for Migrant Workers at High-risk of Covid-19. https://ClinicalTrials.gov/show/NCT04446104 | R | 5000 | Ivermectin 12 mg once vs. HCQ 400 mg day 1 then 200 mg/day for 42 days vs. Zinc 80 mg/day plus vitamin C 500 mg/day for 42 days vs. Povidone-iodine throat spray TID for 42 days vs. Vitamin C 500 mg/day for 42 days | 13.5.20 | Tuas South Dormitory, Singapore, Singapore |
| 16 | New Antiviral Drugs for Treatment of COVID-19. https://ClinicalTrials.gov/show/NCT04392427 | NY | 100 | Ivermectin (dose unlisted) plus NZX (dose unlisted) plus ribavirin 200 mg or 400 mg vs. Control (untreated) | 20.5.20 | Mansoura University, Mansoura, Select A State Or Province, Egypt |
| 17 | Early Treatment With Ivermectin and LosarTAN for Cancer Patients With COVID-19 Infection (TITAN). https://ClinicalTrials.gov/show/NCT04447235 | NY | 176 | Ivermectin 12 mg once plus losartan 50 mg/day for 15 days vs. Placebo | 20.7.20 | Instituto do Cancer do Estado de São Paulo, Brazil |
| 18 | Ivermectin in Treatment of COVID-19. https://ClinicalTrials.gov/show/NCT04445311 | R | 100 | Ivermectin daily (dose unlisted) for 3 days plus SC vs. SC | 31.5.20 | Waheed Shouman, Zagazig, Sharkia, Egypt |
| 19 | Efficacy of Ivermectin in COVID-19. https://ClinicalTrials.gov/show/NCT04392713 | R | 100 | Ivermectin 12 mg once plus SC vs. SC | 15.4.20 | Combined Military Hospital Lahore, Lahore, Punjab, Pakistan |
| 20 | Ivermectin and Doxycycine in COVID-19 Treatment. https://ClinicalTrials.gov/show/NCT04403555 | NY | 40 | Ivermectin (dose unlisted) plus DOC (dose unlisted) vs. CQ (dose unlisted) | 1.6.20 | Sherief Abd-Elsalam, Tanta, Egypt |
| 21 | The Efficacy of Ivermectin and Nitazoxanide in COVID-19 Treatment. https://ClinicalTrials.gov/show/NCT04351347 | R | 300 | Ivermectin (dose unlisted) vs. Ivermectin (dose unlisted) plus NZX (dose unlisted) vs. Ivermectin (dose unlisted) plus CQ (dose unlisted) | 16.6.20 | Tanta University, Tanta, Egypt |
| 22 | Prophylactic Ivermectin in COVID-19 Contacts. https://ClinicalTrials.gov/show/NCT04422561 | C 4 | 304 | Ivermectin 15 mg (40–60 kg), 18 mg (60–80 kg) or 24 mg (> 80 kg) per day, 2 doses 72 h apart vs. Control (untreated) | 31.5.20 | Zagazig University, Zagazig, Sharkia, Egypt |
| 23 | Max Ivermectin- COVID 19 Study Versus Standard of Care Treatment for COVID 19 Cases. A Pilot Study. https://ClinicalTrials.gov/show/NCT04373824 | R | 50 | Ivermectin 0.2 mg/kg daily for 2 days plus SC vs. SC | 25.4.20 | Max Super Speciality hospital, Saket (A unit of Devki Devi Foundation), New Delhi, Delhi, India |
| 24 | A Study to Compare the Efficacy and Safety of Different Doses of Ivermectin for COVID-19 (IFORS) https://ClinicalTrials.gov/show/NCT04431466 | NY | 64 | Ivermectin 0.1 mg/kg once vs. Ivermectin 0.1 mg/kg day 1 and repeated after 72 h vs. Ivermectin 0.2 m/kg once vs. Ivermectin 0.2 mg/kg day 1 and repeated after 72 h vs. SC | 1.7.20 | Hospital Univeristário da Universidade Federal de São Carlos (HU-UFSCar), São Carlos, São Paulo, Brazil |
| 25 | Novel Agents for Treatment of High-risk COVID-19 Positive Patients https://ClinicalTrials.gov/show/NCT04374019 | R | 240 | Ivermectin 12 mg (<75 kg) or 15 mg (>75 kg) daily for 2 days vs. HCQ 600 mg/day for 14 days plus ATM 500 mg day 1 then 250 mg/day for 4 days vs. Camostat Mesilate 200 mg TID for 14 days vs. Artemesia annua 50 mg TID for 14 days | 1.5.20 | University of Kentucky Markey Cancer Center, Lexington, Kentucky, United States |
| 26 | Ivermectin-Azithromycin-Cholecalciferol (IvAzCol) Combination Therapy for COVID-19 (IvAzCol) https://ClinicalTrials.gov/show/NCT04399746 | R | 30 | Ivermectin 6 mg/day on days 0, 1, 7 and 8 plus ATM 500 mg/day 4 days plus Cholecalciferol 400 IU BID for 30 days vs. Control (untreated) | 15.3.20 | Outpatient treatment, Mexico City, Mexico |
| 27 | USEFULNESS of Topic Ivermectin and Carrageenan to Prevent Contagion of Covid 19 (IVERCAR) https://ClinicalTrials.gov/show/NCT04425850 | A 5 | 1195 | Ivermectin (topical for oral mucosae) plus iota carrageenan (topical for oral mucosae) 5 times per day plus PPE vs. PPE only | 1.6.20 | Hospital Eurnekian, Buenos Aires, Argentina |
| 28 | Novel Regimens in COVID-19 Treatment https://ClinicalTrials.gov/show/NCT04382846 | NY | 80 | Ivermectin plus CQ (dose unlisted) vs. Ivermectin plus NZX (dose unlisted) vs. Ivermectin plus NZX plus ATM (dose unlisted) vs. NZX and ATM (dose unlisted) | 8.5.20 | Tanta University, Egypt |
| 29 | Anti-Androgen Treatment for COVID-19 https://ClinicalTrials.gov/show/NCT04446429 | NY | 254 | Ivermectin 0.2 mg/kg QD plus ATM 500 mg QD vs. Ivermectin 0.2 mg/kg QD plus ATM 500 mg QD plus Dutasteride 0.5 mg QD | 26.6.20 | Corpometria Institute, Brasilia, Brazil |
| 30 | A Real-life Experience on Treatment of Patients With COVID 19 https://ClinicalTrials.gov/show/NCT04345419 | R | 120 | Ivermectin (dose unlisted) vs. CQ (dose unlisted) vs. Favipiravir (dose unlisted) vs. NZX (dose unlisted) vs. Niclosamide (dose unlisted) vs. other drugs (oseltamivir or combination of above, dose unlisted) | 16.6.20 | Tanta university hospital, Tanta, Egypt |
| 31 | Worldwide Trends on COVID-19 Research After the Declaration of COVID-19 Pandemic (observational) https://ClinicalTrials.gov/show/NCT04460547 | NY | 200 | Completed interventional vs. completed observational studies on Ivermectin, Convalescent Plasma, HCQ, DAS181, or Interferon 1A | 25.7.20 | Qassim University, Saudi Arabia |
| 32 | Trial of Combination Therapy to Treat COVID-19 Infection https://ClinicalTrials.gov/show/NCT04482686 | NY | 300 | Ivermectin (dose unlisted) day 1 and 4 plus DOC (dose unlisted) for 10 days plus Zinc for 10 days plus Vitamin D3 for 10 days plus Vitamin C for 10 days vs. Placebo | 22.7.20 | ProgenaBiome, California, USA |
| 33 | Randomised clinical trial of ivermectin for treatment and prophylaxis of COVID-19 https://www.clinicaltrialsregister.eu/ctr-search/trial/2020-001994-66/ES | O | 266 | Ivermectin (dose unlisted) vs. Placebo | 8.5.20 | Fundació Assistencial Mútua Terrassa, Spain |
| 34 | Multicenter, randomized, double-blind, placebo-controlled study investigating efficacy, safety and tolerability of ivermectin HUVE-19 in patients with proven SARS-CoV-2 infection (COVID-19) and manifested clinical symptoms. https://www.clinicaltrialsregister.eu/ctr-search/trial/2020-002091-12/BG | O | 120 | Ivermectin 0.4 mg/kg plus SC vs. Placebo plus SC | 5.5.20 | Bulgaria (9 sites) |
| 35 | Efficacy of hydroxychloroquine, ciclesonide and ivermectin in treatment of moderate covid-19 illness: an open-label randomised controlled study (EHYCIVER-COVID) http://ctri.nic.in/ClinicaltrialsCTRI/2020/04/024948 | NY | 120 | Ivermectin 12 mg/day for 7 days vs. Ciclesonide 0.2 mg/kg BID for 7 days vs. HCQ 400 mg BID Day 1 then 200 mg BID for 6 days vs. SC | 15.5.20 | New Delhi, India |
| 36 | A Phase IIB open label randomized controlled trial to evaluate the efficacy and safety of Ivermectin in reducing viral loads in patients with hematological disorders who are admitted with COVID 19 infection http://ctri.nic.in/ClinicaltrialsCTRI/2020/04/025068 | NY | 50 | Ivermectin 3 mg (15–24 kg) or 6 mg (25–35 kg) or 9 mg (36–50 kg) or 12 mg (51–65 kg) or 15 mg (66–79 kg) or 0.2 mg/kg (80 kg) once vs. SC | 27.5.20 | Christian Medical College Vellore, TAMIL NADU, India |
| 37 | Interventional study to assess the efficacy of Ivermectin with standard of care treatment versus standard of care in patients of COVID-19 at R D Gardi Medical College, Ujjain, India http://ctri.nic.in/ClinicaltrialsCTRI/2020/04/025224 | NY | 50 | Ivermectin 12 mg/day for 2 days plus SC vs. SC | 24.5.20 | R D Gardi Medical College, Ujjain, Madhya Pradesh, India |
| 38 | Study to assess the efficacy of Ivermectin as prophylaxis of COVID 19 among health care workers and COVID 19 contacts in Ujjain, India; http://ctri.nic.in/ClinicaltrialsCTRI/2020/04/025333 | NY | 2000 | Ivermectin 12 mg/day (adult) or 6 mg/day (children) for 2 days vs. Control | 27.5.20 | R D Gardi Medical College, Ujjain, Madhya Pradesh, India |
| 39 | Randomised Controlled Trial of Ivermectin in hospitalised patients with COVID19 (RIVET-COV). http://ctri.nic.in/ClinicaltrialsCTRI/2020/04/026001 | NY | 60 | Ivermectin single dosing of 0.2 mg/kg vs. Ivermectin 0.4 mg/kg vs. Ivermectin 0.8 mg/kg vs. Ivermectin 1.6 mg/kg vs. Ivermectin 2 mg/kg vs. SC | 25.6.20 | New Delhi, India |
| 40 | A Prospective, randomized, single centred, open labelled, two arm, placebo-controlled trial to evaluate efficacy and safety of Ivermectin drug in patients infected with SARS-CoV-2 virus; http://ctri.nic.in/ClinicaltrialsCTRI/2020/04/025960 | NY | 100 | Ivermectin 12 mg/day for 3 days vs. SC | 18.6.20 | Symbiosis University Hospital and Research Centre, Maharashtra, India |
| 41 | A Clinical Trial to Study the Efficacy of “Ivermectin” in the prevention of Covid-19. A Single Arm Study. http://ctri.nic.in/ClinicaltrialsCTRI/2020/04/026232 | NY | 50 | Ivermectin 0.2 mg/kg once | 10.7.20 | DVFM, Andhra Pradesh, India |
| 42 | Ivermectin Nasal Spray for COVID19 Patients. https://ClinicalTrials.gov/show/NCT04510233 | NY | 60 | Ivermectin nasal spray (1 mL) in each nostril BID vs. Ivermectin oral (6 mg) TID vs. SC | 10.8.20 | Tanta University, Tanta, Egypt |
| 43 | Outpatient use of ivermectin in COVID-19. https://ClinicalTrials.gov/show/NCT04530474 | NY | 200 | Ivermectin 0.15–0.2 mg/kg (max 12 mg) once vs. Placebo | 26.8.20 | Temple University Hospital, Philadelphia, USA |
| 44 | Ivermectin to prevent hospitilizations in COIVD-19. https://ClinicalTrials.gov/show/NCT04529525 | R | 500 | Ivermectin 12 mg (48–80 kg) or 18 mg (80–110 kg) or 24 mg (>100 kg) at inclusion and again at 24h vs. Placebo | 21.8.20 | Ministry of Public Health, Province of Corrientes, Argentina |
| 45 | Clinical trial of ivermectin plus doxycycline for the treatment of confirmed Covid-19 infection. https://ClinicalTrials.gov/show/NCT04523831 | R | 400 | Ivermectin 6 mg and doxycycline 100 mg BID for 5 days vs. Placebo | 19.8.20 | Dhaka Medical College, Dhaka Bangladesh |
| 46 | Pilot study to evaluate the potential of ivermectin to reduce COVID-19 transmission. https://www.clinicaltrialsregister.eu/ctr-search/trial/2020-001474-29/ES | O | 24 | Ivermectin (dose unlisted) vs. Placebo | 8.5.20 | Clinica Universidad de Navarra, Pamplona, Spain |
| 47 | Dose-Finding study of Ivermectin treatment on patients infected with Covid-19: A clinical trial. https://en.irct.ir/trial/47012 | A | 125 | Ivermectin 0.2 mg/kg single dose plus SC vs. Ivermectin 0.2 mg/kg day 1, 2, 5 plus SC vs. Placebo plus SC vs. Ivermectin 0.4 mg/kg day 1 and 0.2 mg/kg day 2, 5 vs. SC | 4.5.20 | Qazvin University of Medical Sciences, Qazvin, Iran |
| 48 | In vivo use of ivermectin (IVR) for treatment for corona virus infected patients: a randomized controlled trial. http://www.chictr.org.cn/showprojen.aspx?proj=54707 | NY | 60 | Ivermectin single dose 0.2 mg/kg vs. Placebo | 10.6.20 | Rayak Hospital, Riyaq, Lebanon |
| 49 | A randomized clinical trial study, comparison of the therapeutic effects of Ivermectin, Kaletra and Chloroquine with Kaletra and Chloroquine in the treatment of patients with coronavirus 2019 (COVID-19). http://en.irct.ir/trial/48444 | A | 60 | Ivermectin 0.15–0.2 mg/kg single dose day 1 plus HCQ 200 mg day 1 plus Lopinavir/Ritonavir 400/100 mg days 2–6 vs. HCQ 200 mg day 1 plus Lopinavir/Ritonavir 400/100 mg days 2-6 | 30.5.20 | Ahvaz Razi Hospital, Ahvaz, Iran |
| 50 | A double-blind clinical trial to repurpose and assess the efficacy and safety of ivermectin in COVID-19. http://isrctn.com/ISRCTN40302986 | R | 45 | Ivermectin 6 mg every 3.5 days for 2 weeks vs. Ivermectin 12 mg every 3.5 days for 2 weeks vs. Placebo | 23.4.20 | Lagos University Teaching Hospital, Lagos, Nigeria |
| 51 | Effectiveness of Ivermectin in the Treatment of Coronavirus Infection in Patients admitted to Educational Hospitals of Mazandaran in 2020. https://en.irct.ir/trial/49174 | R | 60 | Ivermectin 0.2 mg/kg once plus SC vs. SC | 21.5.20 | Bouali Hospital, Sari, Iran |
| 52 | Subcutaneous Ivermectin in Combination With and Without Oral Zinc and Nigella Sativa: a Placebo Randomized Control Trial on Mild to Moderate COVID-19 Patients. https://clinicaltrials.gov/ct2/show/study/NCT04472585 | R | 40 | Ivermectin 0.2 mg/kg subcutaneous injection every 2 days plus SC vs. Ivermectin 0.2 mg/kg subcutaneous injection every 2 days plus 80 mg/kg Nigella Sativa oral QD plus SC vs. Ivermectin 0.2 mg/kg subcutaneous injection every 2 days plus 20 mg Zinc Sulfate oral TID plus SC vs. Placebo plus SC | 14.7.20 | Shaikh Zayed Hospital, Lahore, Pakistan |
| 53 | Pragmatic study “CORIVER”: Ivermectin as antiviral treatment for patients infected by SARS-COV2 (COVID-19). https://www.clinicaltrialsregister.eu/ctr-search/trial/2020-001971-33/ES | O | 45 | Ivermectin 0.2–0.4 mg/kg (regime unlisted) vs. HCQ 400 mg vs. ATM 500 mg vs. Placebo | 22.7.20 | Hospital Universitario Virgen de las Nieves, Granada, Spain |
| 54 | Effectiveness and Safety of Ivermectin for the Prevention of Covid-19 Infection in Colombian Health Personnel at All Levels of Care, During the 2020 Pandemic: A Randomized Clinical Controled Trial. https://clinicaltrials.gov/ct2/show/record/NCT04527211 | NY | 550 | Ivermectin 0.2 mg/kg weekly for 7 weeks vs. Placebo | 7.9.20 | Pontificia Universidad Javeriana, Valle Del Cauca, Colombia |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jans, D.A.; Wagstaff, K.M. Ivermectin as a Broad-Spectrum Host-Directed Antiviral: The Real Deal? Cells 2020, 9, 2100. https://doi.org/10.3390/cells9092100
Jans DA, Wagstaff KM. Ivermectin as a Broad-Spectrum Host-Directed Antiviral: The Real Deal? Cells. 2020; 9(9):2100. https://doi.org/10.3390/cells9092100
Chicago/Turabian StyleJans, David A., and Kylie M. Wagstaff. 2020. "Ivermectin as a Broad-Spectrum Host-Directed Antiviral: The Real Deal?" Cells 9, no. 9: 2100. https://doi.org/10.3390/cells9092100