Phenol-Soluble Modulin-Mediated Aggregation of Community-Associated Methicillin-Resistant Staphylococcus Aureus in Human Cerebrospinal Fluid


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Bacterial Strains and Cell Cultures
2.3. CSF-Induced Cell Aggregation Assay
2.4. Aggregation Image Analysis
2.5. PSM Quantification
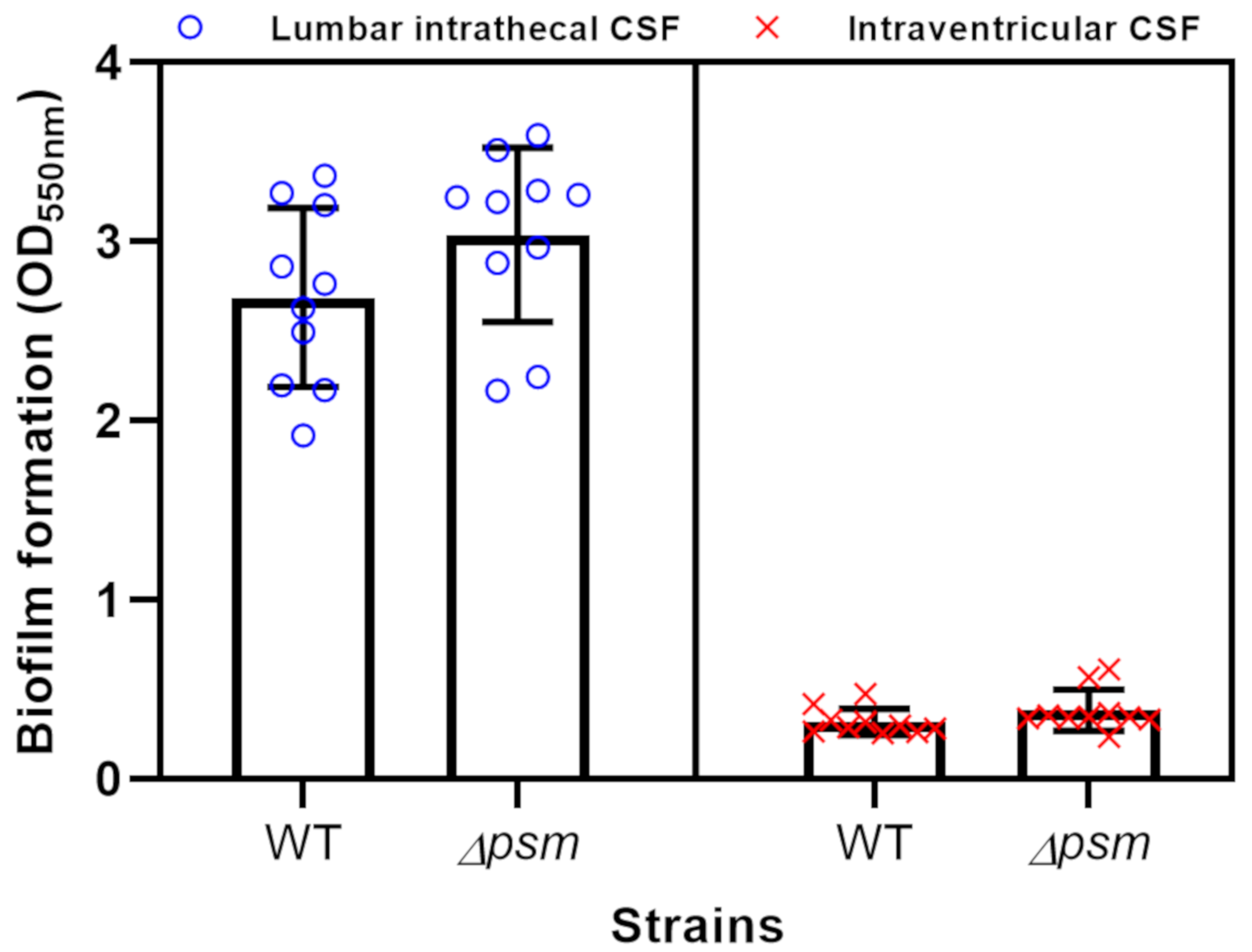
2.6. Biofilm Assay
2.7. Statistical Analyses
3. Results
3.1. Patient Demographics
3.2. Difference in Cell Aggregation According to CSF Sampling Site
3.3. Difference in Cell Aggregation According to PSM Presence
3.4. Biofilm Formation
4. Discussion
4.1. CSF as a Spokesman for the CNS Environment
4.2. Difference in Cell Aggregation According to CSF Sampling Site
4.3. Difference in Cell Aggregation According to PSM Presence
4.4. The Effect of Antibiotics
4.5. The Role of PSMs on Biofilm Formation in Human CSF
4.6. Study Limitations
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G., Jr. Methicillin-resistant Staphylococcus aureus: An overview of basic and clinical research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Community-associated MRSA: What makes them special? Int. J. Med. Microbiol. 2013, 303, 324–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, E.F.; Johnson, J.K.; Jabra-Rizk, M.A. Community-Associated Methicillin-Resistant Staphylococcus aureus: An Enemy amidst Us. PLoS Pathog. 2016, 12, e1005837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Bayer, A.; Cosgrove, S.E.; Daum, R.S.; Fridkin, S.K.; Gorwitz, R.J.; Kaplan, S.L.; Karchmer, A.W.; Levine, D.P.; Murray, B.E.; et al. Clinical practice guidelines by the infectious diseases society of america for the treatment of methicillin-resistant Staphylococcus aureus infections in adults and children: Executive summary. Clin. Infect. Dis. 2011, 52, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Haste, N.M.; Perera, V.R.; Maloney, K.N.; Tran, D.N.; Jensen, P.; Fenical, W.; Nizet, V.; Hensler, M.E. Activity of the streptogramin antibiotic etamycin against methicillin-resistant Staphylococcus aureus. J. Antibiot. (Tokyo) 2010, 63, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Cheung, G.Y.; Wang, R.; Khan, B.A.; Sturdevant, D.E.; Otto, M. Role of the accessory gene regulator agr in community-associated methicillin-resistant Staphylococcus aureus pathogenesis. Infect. Immun. 2011, 79, 1927–1935. [Google Scholar] [CrossRef] [Green Version]
- Fridkin, S.K.; Hageman, J.C.; Morrison, M.; Sanza, L.T.; Como-Sabetti, K.; Jernigan, J.A.; Harriman, K.; Harrison, L.H.; Lynfield, R.; Farley, M.M. Methicillin-resistant Staphylococcus aureus disease in three communities. N. Engl. J. Med. 2005, 352, 1436–1444. [Google Scholar] [CrossRef]
- David, M.Z.; Daum, R.S. Community-associated methicillin-resistant Staphylococcus aureus: Epidemiology and clinical consequences of an emerging epidemic. Clin. Microbiol. Rev. 2010, 23, 616–687. [Google Scholar] [CrossRef] [Green Version]
- Naesens, R.; Ronsyn, M.; Druwe, P.; Denis, O.; Ieven, M.; Jeurissen, A. Central nervous system invasion by community-acquired meticillin-resistant Staphylococcus aureus. J. Med. Microbiol. 2009, 58, 1247–1251. [Google Scholar] [CrossRef]
- Peschel, A.; Otto, M. Phenol-soluble modulins and staphylococcal infection. Nat. Rev. Microbiol. 2013, 11, 667–673. [Google Scholar] [CrossRef]
- Wang, R.; Braughton, K.R.; Kretschmer, D.; Bach, T.H.; Queck, S.Y.; Li, M.; Kennedy, A.D.; Dorward, D.W.; Klebanoff, S.J.; Peschel, A.; et al. Identification of novel cytolytic peptides as key virulence determinants for community-associated MRSA. Nat. Med. 2007, 13, 1510–1514. [Google Scholar] [CrossRef] [PubMed]
- Dastgheyb, S.S.; Villaruz, A.E.; Le, K.Y.; Tan, V.Y.; Duong, A.C.; Chatterjee, S.S.; Cheung, G.Y.; Joo, H.S.; Hickok, N.J.; Otto, M. Role of phenol-soluble modulins in formation of Staphylococcus aureus biofilms in synovial fluid. Infect. Immun. 2015, 83, 2966–2975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longhurst, W.D.; Sheele, J.M. Spontaneous methicillin-resistant Staphylococcus aureus (MRSA) meningitis. Am. J. Emerg. Med. 2018, 36, 909. [Google Scholar] [CrossRef] [PubMed]
- Pintado, V.; Pazos, R.; Jiménez-Mejías, M.E.; Rodríguez-Guardado, A.; Gil, A.; García-Lechuz, J.M.; Cabellos, C.; Chaves, F.; Domingo, P.; Ramos, A.; et al. Methicillin-resistant Staphylococcus aureus meningitis in adults: A multicenter study of 86 cases. Medicine 2012, 91, 10–17. [Google Scholar] [CrossRef]
- Fortuna, D.; Hooper, D.C.; Roberts, A.L.; Harshyne, L.A.; Nagurney, M.; Curtis, M.T. Potential role of CSF cytokine profiles in discriminating infectious from non-infectious CNS disorders. PLoS ONE 2018, 13, e0205501. [Google Scholar] [CrossRef] [Green Version]
- Joo, H.S.; Cheung, G.Y.; Otto, M. Antimicrobial activity of community-associated methicillin-resistant Staphylococcus aureus is caused by phenol-soluble modulin derivatives. J. Biol. Chem. 2011, 286, 8933–8940. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Joo, H.S.; Otto, M. The isolation and analysis of phenol-soluble modulins of Staphylococcus epidermidis. Methods Mol. Biol. 2014, 1106, 93–100. [Google Scholar]
- Piewngam, P.; Zheng, Y.; Nguyen, T.H.; Dickey, S.W.; Joo, H.S.; Villaruz, A.E.; Glose, K.A.; Fisher, E.L.; Hunt, R.L.; Li, B.; et al. Pathogen elimination by probiotic Bacillus via signalling interference. Nature 2018, 562, 532–537. [Google Scholar] [CrossRef]
- Matsumae, M.; Sato, O.; Hirayama, A.; Hayashi, N.; Takizawa, K.; Atsumi, H.; Sorimachi, T. Research into the physiology of cerebrospinal fluid reaches a new horizon: Intimate exchange between cerebrospinal fluid and interstitial fluid may contribute to maintenance of homeostasis in the central nervous system. Neurol. Med. Chir. (Tokyo) 2016, 56, 416–441. [Google Scholar] [CrossRef] [Green Version]
- Griffin, D.E. Emergence and re-emergence of viral diseases of the central nervous system. Prog. Neurobiol. 2010, 91, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Kothur, K.; Wienholt, L.; Brilot, F.; Dale, R.C. CSF cytokines/chemokines as biomarkers in neuroinflammatory CNS disorders: A systematic review. Cytokine 2016, 77, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Kwon, B.K.; Stammers, A.M.; Belanger, L.M.; Bernardo, A.; Chan, D.; Bishop, C.M.; Slobogean, G.P.; Zhang, H.; Umedaly, H.; Giffin, M.; et al. Cerebrospinal fluid inflammatory cytokines and biomarkers of injury severity in acute human spinal cord injury. J. Neurotrauma 2010, 27, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Berezovsky, D.E.; Bruce, B.B.; Vasseneix, C.; Peragallo, J.H.; Newman, N.J.; Biousse, V. Cerebrospinal fluid total protein in idiopathic intracranial hypertension. J. Neurol. Sci. 2017, 381, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Basis of virulence in community-associated methicillin-resistant Staphylococcus aureus. Annu. Rev. Microbiol. 2010, 64, 143–162. [Google Scholar] [CrossRef]
- Periasamy, S.; Joo, H.S.; Duong, A.C.; Bach, T.H.; Tan, V.Y.; Chatterjee, S.S.; Cheung, G.Y.; Otto, M. How Staphylococcus aureus biofilms develop their characteristic structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1281–1286. [Google Scholar] [CrossRef] [Green Version]
- Spaulding, A.R.; Satterwhite, E.A.; Lin, Y.C.; Chuang-Smith, O.N.; Frank, K.L.; Merriman, J.A.; Schaefers, M.M.; Yarwood, J.M.; Peterson, M.L.; Schlievert, P.M. Comparison of Staphylococcus aureus strains for ability to cause infective endocarditis and lethal sepsis in rabbits. Front. Cell. Infect. Microbiol. 2012, 2, 18. [Google Scholar] [CrossRef] [Green Version]
- Davido, B.; Saleh-Mghir, A.; Laurent, F.; Danel, C.; Couzon, F.; Gatin, L.; Vandenesch, F.; Rasigade, J.P.; Cremieux, A.C. Phenol-soluble modulins contribute to early sepsis dissemination not late local USA300-osteomyelitis severity in rabbits. PLoS ONE 2016, 11, e0157133. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, K.; Syed, A.K.; Stephenson, R.E.; Rickard, A.H.; Boles, B.R. Functional amyloids composed of phenol soluble modulins stabilize Staphylococcus aureus biofilms. PLoS Pathog. 2012, 8, e1002744. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | N (Percentage) or Median [IQR] |
|---|---|
| Age (years) | 63.0 [49.0, 75.0] |
| Sex | |
| Male | 30 (44.1%) |
| Female | 38 (55.9%) |
| CSF sampling site | |
| Intraventricular | 15 (22.1%) |
| Intrathecal | 53 (77.9%) |
| Antibiotics use before sampling | 8 (11.8%) |
| Diagnosis | |
| Normal pressure hydrocephalus | 19 (27.9%) |
| Migraine, Tension-type headache | 18 (26.5%) |
| Infectious disease | 12 (17.6%) |
| Cerebrovascular disease | 11 (16.2%) |
| Neurodegenerative disease | 6 (8.8%) |
| Epilepsy | 1 (1.5%) |
| Myopathy | 1 (1.5%) |
| CSF analysis | |
| Normal CSF | 40 (58.8%) |
| Glucose (mg/dL) | 64.0 [54.0, 79.2] |
| Protein (mg/dL) | 36.0 [26.2, 56.2] |
| Increased RBC (≥1) | 23 (33.8%) |
| Increased WBC (≥5) | 11 (16.2%) |
| Cell aggregation pattern | |
| Active cell aggregation | 54 (79.4%) |
| Centeroid | 40 (74.0%) |
| Oval scattered | 7 (13.0%) |
| Streaming | 7 (13.0%) |
| Robust bacterial culture | 9 (13.2%) |
| Neither aggregation nor bacterial culture | 5 (7.4%) |
| Variable | Intraventricular (N = 15) | Intrathecal (N = 53) | p Value |
|---|---|---|---|
| Age (years) | 64.0 [56.5, 75.0] | 61.0 [40.0, 73.0] | 0.311 |
| Sex | 0.603 | ||
| Male | 8 (53.5%) | 22 (41.5%) | |
| Female | 7 (46.7%) | 31 (58.5%) | |
| Antibiotics Use before Sampling | 1 (6.7%) | 7 (13.2%) | 0.674 |
| CSF analysis | |||
| Normal CSF | 8 (53.3%) | 32 (60.4%) | 0.848 |
| Glucose (mg/dL) | 63.0 [55.0, 69.0] | 64.0 [54.0, 82.0] | 0.446 |
| Protein (mg/dL) | 17.4 [13.4, 28.0] | 42.7 [29.7, 62.9] | <0.001 |
| Increased RBC (≥1) | 7 (46.7%) | 16 (30.2%) | 0.378 |
| Increased WBC (≥5) | 0 (0%) | 11 (20.8%) | 0.105 |
| WT aggregation area (%) | 0.2 [0.0, 0.9] | 6.7 [4.5, 9.9] | <0.001 |
| Δpsm aggregation area (%) | 0.0 [0.0, 0.2] | 1.2 [0.5, 2.2] | <0.001 |
| Variable | WT/ΔPSM ≥ 2 (N = 43) | WT/ΔPSM < 2 (N = 25) | p Value |
|---|---|---|---|
| Age (years) | 64.0 [53.5, 72.5] | 57.0 [40.0, 75.0] | 0.620 |
| Sex | 0.456 | ||
| Male | 17 (39.5%) | 13 (52.0%) | |
| Female | 7 (46.7%) | 31 (58.5%) | |
| CSF sampling site | 0.002 | ||
| Intraventricular | 4 (9.3%) | 11 (44.0%) | |
| Intrathecal | 39 (90.7%) | 14 (56.0%) | |
| Antibiotics use before sampling | 3 (7.0%) | 5 (20.0%) | 0.133 |
| CSF analysis | |||
| Normal CSF | 28 (65.1%) | 12 (48.0%) | 0.260 |
| Glucose (mg/dL) | 68.0 [55.0, 82.0] | 56.0 [53.0, 67.0] | 0.059 |
| Protein (mg/dL) | 34.2 [27.2, 47.4] | 48.4 [20.6, 81.4] | 0.457 |
| Increased RBC (≥1) | 13 (30.2%) | 10 (40.0%) | 0.579 |
| Increased WBC (≥5) | 5 (11.6%) | 6 (24.0%) | 0.305 |
| WT aggregation area (%) | 6.9 [5.0, 9.7] | 1.0 [0.0, 6.2] | <0.001 |
| Δpsm aggregation area (%) | 1.2 [0.5, 1.5] | 0.0 [0.0, 3.7] | 0.050 |
| WT/Δpsm | 5.5 [4.0, 12.1] | 0.0 [0.0, 1.3] | <0.001 |
| Parameter | Crude OR [95%CI] | Crude p Value | Adjusted OR [95%CI] | Adjusted p Value |
|---|---|---|---|---|
| Age | 0.99 [0.07–1.38] | 0.641 | 0.99 [0.96–1.03] | 0.699 |
| Male | 0.6 [0.22–1.63] | 0.32 | 0.54 [0.15–1.92] | 0.341 |
| Intraventricular CSF sampling Antibiotics use | 0.13 [0.04–0.48] 0.3 [0.07–1.38] | 0.002 0.123 | 0.1 [0.02–0.44] 0.51 [0.06–4.52] | 0.002 0.545 |
| Normal CSF | 2.02 [0.74–5.52] | 0.169 | 1.55 [0.08–31.03] | 0.773 |
| Increased RBC | 0.65 [0.23–1.82] | 0.413 | 1.23 [0.08–18] | 0.878 |
| Increased WBC | 0.42 [0.11–1.54] | 0.19 | 0.41 [0.03–5.66] | 0.504 |
| Protein | 1 [0.99–1.01] | 0.978 | 1 [0.99–1.01] | 0.971 |
| Glucose | 0.98 [0.97–1.02] | 0.057 | 0.98 [0.95–1.01] | 0.182 |
| Variable | WT/ΔPSM ≥ 2 (N = 39) | WT/ΔPSM < 2 (N = 14) | p Value |
|---|---|---|---|
| Age (years) | 64.0 [53.5, 74.0] | 51.0 [33.2, 55.0] | 0.161 |
| Sex | 0.663 | ||
| Male | 15 (38.5%) | 7 (50.0%) | |
| Female | 24 (61.5%) | 7 (50.0%) | |
| Antibiotics use before sampling | 3 (7.7%) | 4 (28.6%) | 0.133 |
| CSF analysis | |||
| Normal CSF | 27 (69.2%) | 5 (35.7%) | 0.060 |
| Glucose (mg/dL) | 68.0 [55.0, 83.2] | 54.5 [50.0, 64.2] | 0.072 |
| Protein (mg/dL) | 36.0 [28.9, 48.6] | 68.1 [48.5, 94.8] | 0.002 |
| Increased RBC (≥1) | 10 (25.6%) | 6 (42.9%) | 0.311 |
| Increased WBC (≥5) | 5 (12.8%) | 6 (42.9%) | 0.049 |
| WT aggregation area (%) | 6.9 [5.2, 10.3] | 4.4 [1.3, 7.8] | 0.023 |
| Δpsm aggregation area (%) | 1.2 [0.7, 1.5] | 2.2 [0.1, 5.3] | 0.518 |
| WT/Δpsm | 5.5 [4.3, 14.2] | 1.1 [0.2, 1.4] | <0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.-r.; Lee, Y.; Kim, H.-k.; Kim, W.; Kim, Y.-G.; Yang, Y.-H.; Kim, J.-S.; Joo, H.-S. Phenol-Soluble Modulin-Mediated Aggregation of Community-Associated Methicillin-Resistant Staphylococcus Aureus in Human Cerebrospinal Fluid. Cells 2020, 9, 788. https://doi.org/10.3390/cells9030788
Kim D-r, Lee Y, Kim H-k, Kim W, Kim Y-G, Yang Y-H, Kim J-S, Joo H-S. Phenol-Soluble Modulin-Mediated Aggregation of Community-Associated Methicillin-Resistant Staphylococcus Aureus in Human Cerebrospinal Fluid. Cells. 2020; 9(3):788. https://doi.org/10.3390/cells9030788
Chicago/Turabian StyleKim, Deok-ryeong, Yeonhee Lee, Hyeon-kyeong Kim, Wooseong Kim, Yun-Gon Kim, Yung-Hun Yang, Jae-Seok Kim, and Hwang-Soo Joo. 2020. "Phenol-Soluble Modulin-Mediated Aggregation of Community-Associated Methicillin-Resistant Staphylococcus Aureus in Human Cerebrospinal Fluid" Cells 9, no. 3: 788. https://doi.org/10.3390/cells9030788