Fluorinated Galactoses Inhibit Galactose-1-Phosphate Uridyltransferase and Metabolically Induce Galactosemia-like Phenotypes in HEK-293 Cells

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fluorinated Galactose Derivatives and Other Materials
2.2. Preparation of 2-fluoro-2-deoxy-galactose-1-phosphate (F-Gal-1-P)
2.3. Preparation of 2,2′-difluoro-2-deoxy-galactose (F2-Gal) and 2,2′-difluoro-2-deoxy-galactose-1-phosphate (F2-Gal-1-P) and Its per-O-acetylated Derivative
2.4. Cells and Cell Culture
2.5. Recombinant Expression and Purification of Human GALT in E. coli
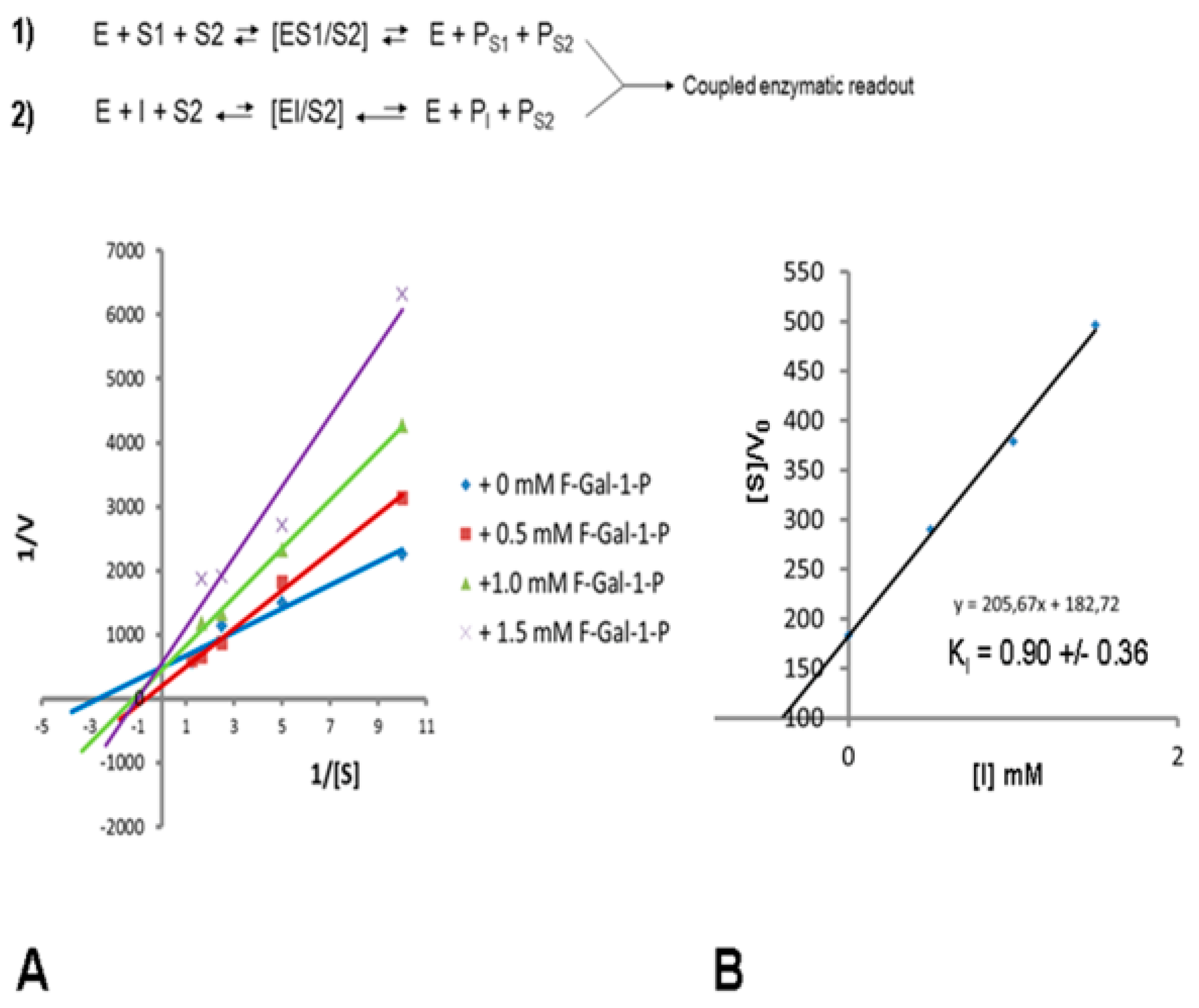
2.6. Kinetic Studies of Recombinant Human GALT by Coupled Enzyme Assay
2.7. Analysis of Relative Conversion Rates of Gal-1-P and F-Gal-1-P
2.8. Differential N-glycosylation Analyses of Secretory Chicken Netrin-1 Formed in the Presence or Absence of F-Gal
2.9. Fluorescence-Activated Cell Sorting (FACS) of HEK293 Cells and Human Fibroblasts
2.10. Electrophoresis and Western Blot Analysis
2.11. Differential Label-Free Proteomics
3. Results
3.1. Kinetic Studies of Recombinant hGALT Activity in the Presence of Fluorinated Galactose Inhibitors
3.2. N-Glycosylation of Recombinant Probes in the Presence of Fluorinated Galactose
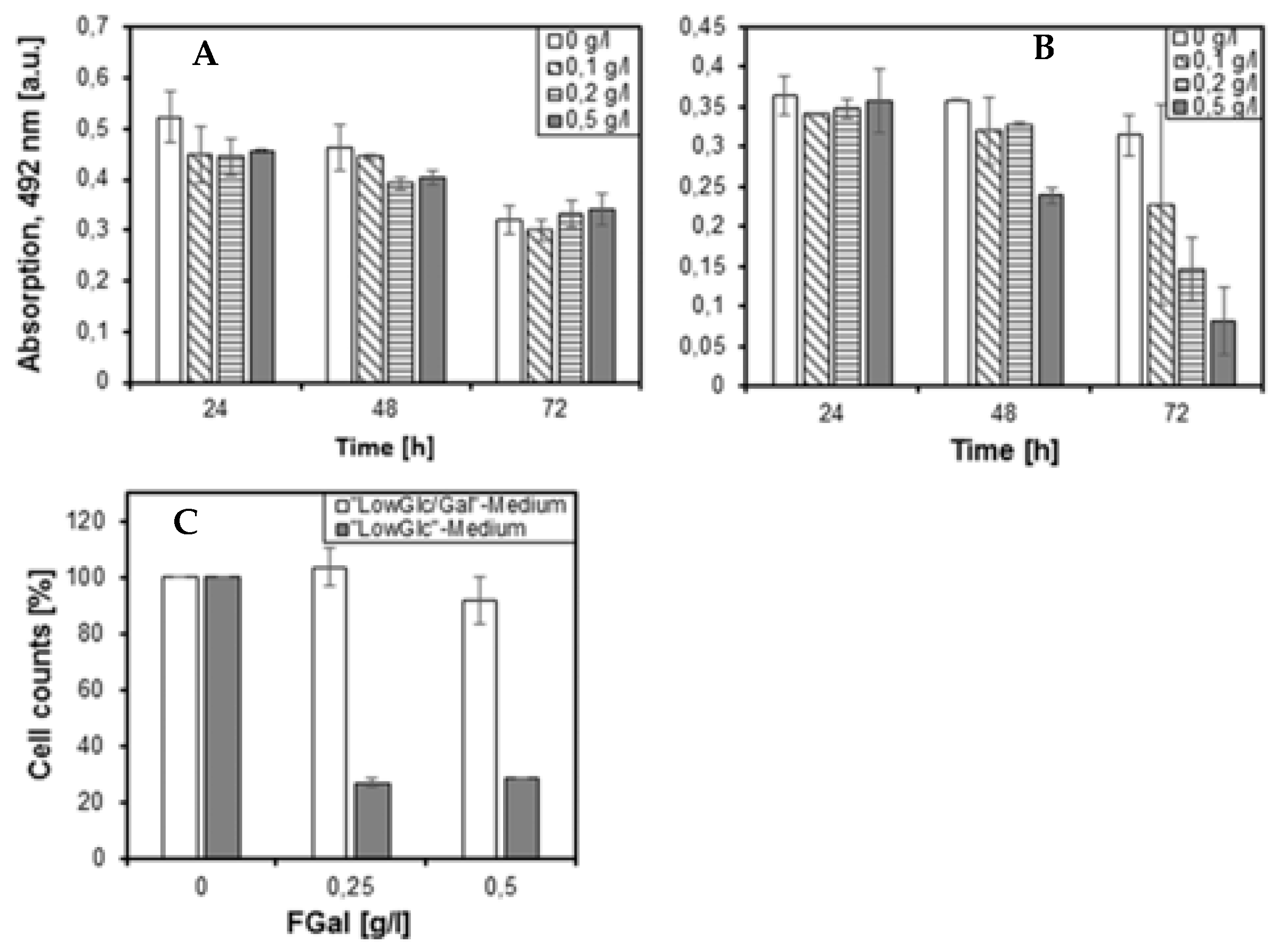
3.3. Establishment of the Epithelial Cell Model (HEK293 and HEK293-MUC1)
3.4. FACS Reveals Altered Plasma Membrane Localisation of Glycoprotein Markers Induced by F-Gal and F2-Gal
3.5. Differential Proteomics of Lipid Rafts from MIG-HEK293 and Control Cells
3.6. Differential Proteomics of Lipid Rafts from Galactosemic and Healthy Control Fibroblasts
4. Discussion
4.1. General Discussion
4.2. HEK-293 Cells Grown in the Presence of Fluorinated Galactose Show a Metabolically Induced Galactosemic Phenotype (MIG)
4.3. Molecular Pathways Affected in Classical Galactosemia
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bosch, A. Classical galactosaemia revisited. J. Inherit. Metab. Dis. 2006, 29, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Charlwood, J.; Clayton, P.; Keir, G.; Mian, N.; Winchester, B. Defective galactosylation of serum transferrin in galactosemia. Glycobiology 1998, 8, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Staubach, S.; Schadewaldt, P.; Wendel, U.; Nohroudi, K.; Hanisch, F.G. Differential glycomics of epithelial membrane glycoproteins from urinary exovesicles reveals shifts toward complex-type N-glycosylation in classical galactosemia. J. Proteome Res. 2012, 11, 906–916. [Google Scholar] [CrossRef] [PubMed]
- Staubach, S.; Pekmez, M.; Hanisch, F.G. Differential proteomics of urinary exovesicles from classical galactosemic patients reveals subclinical kidney insuffiency. J. Proteome Res. 2016, 15, 1754–1761. [Google Scholar] [CrossRef] [PubMed]
- Staubach, S.; Müller, S.; Pekmez, M.; Hanisch, F.G. Classical galactosemia: Insight into molecular pathomechanisms by differential membrane proteomics of fibroblasts under galactose stress. J. Proteome Res. 2017, 16, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Slepak, T.I.; Tang, M.; Slepak, V.Z.; Lai, K. Involvement of endoplasmic reticulum stress in a novel Classic Galactosemia model. Mol. Genet. Metab. 2007, 92, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Potter, B.A.; Hughey, R.P.; Weisz, O.A. Role of N- and O-glycans in polarized biosynthetic sorting. Am. J. Physiol. Cell. Physiol. 2006, 290, C1–C10. [Google Scholar] [CrossRef] [Green Version]
- Shaw, G.; Morse, S.; Ararat, M.; Graham, F.L. Preferential transformation of human neuronal cells by human adenoviruses and the origin of HEK 293 cells. FASEB J. 2002. [Google Scholar] [CrossRef]
- Wahl, C.; Hirtz, D.; Elling, L. Multiplexed capillary electrophoresis as analytical tool for fast optimization of multi-enzyme cascade reactions—Synthesis of nucleotide sugars. Biotechnol. J. 2016, 11, 1298–1308. [Google Scholar] [CrossRef]
- Wahl, C.; Spiertz, M.; Elling, L. Characterization of a new UDP-sugar pyrophosphorylase from Hordeum vulgare (barley). Biotechnol. J. 2017, 258, 51–55. [Google Scholar] [CrossRef]
- Fischöder, T.; Laaf, D.; Dey, C.; Elling, L. Enzymatic Synthesis of N-Acetyllactosamine (LacNAc) Type 1 Oligomers and Characterization as Multivalent Galectin Ligand. Molecules 2017, 22, 1320. [Google Scholar] [CrossRef] [PubMed]
- Francisco, C.G.; Gonzalez, C.C.; Kennedy, A.R.; Paz, N.R.; Suarez, E. Fragmentation of carbohydrate anomeric alkoxyl radicals: New synthesis of chiral 1-fluoro-1-halo-1-iodoalditols. Chem. Eur. J. 2008, 14, 6704–6712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, S.P.; Tikad, A. β-Selective One-Pot Fluorophosphorylation of d,d-Heptosylglycals Mediated by Selectfluor. Isr. J. Chem. 2015, 55, 392–397. [Google Scholar] [CrossRef]
- Dumitrescu, L.; Eppe, G.; Tikad, A.; El Bkassiny, S.; Pan, W.; Gurcha, S.; Besra, G.; Arda, A.; Jiménez-Barbero, J.; Vincent, S.P. Selectfluor and NFSI exo-glycal fluorination strategies applied to the enhancement of the binding affinity of galactofuranosyl transferase GalfT2 inhibitors. Chem. Eur. J. 2014, 20, 15208–15215. [Google Scholar] [CrossRef]
- Dohi, H.; Périon, R.; Durka, M.; Bosco, M.; Roué, Y.; Moreau, F.; Grizot, S.; Ducruix, A.; Escaich, S.; Vincent, S.P. Stereoselective glycal fluorophosphorylation: Synthesis of ADP-2-fluoroheptose, an inhibitor of the LPS biosynthesis. Chem. Eur. J. 2008, 14, 9530–9539. [Google Scholar] [CrossRef]
- Engelmann, K.; Kinlough, C.L.; Müller, S.; Razawi, H.; Baldus, S.E.; Hughey, R.P.; Hanisch, F.G. Transmembrane and secreted MUC1 probes show trafficking-dependent changes in O-glycan core profiles. Glycobiology 2005, 15, 1111–1124. [Google Scholar] [CrossRef] [Green Version]
- Crews, C.; Wilkinson, K.D.; Wells, L.; Perkins, C.; Fridovich-Keil, J.L. Functional consequence of substitutions at residue 171 in human galactose-1-phosphate uridylyltransferase. J. Biol. Chem. 2000, 275, 22847–22853. [Google Scholar] [CrossRef] [Green Version]
- Vihar, B.; Hanisch, F.G.; Baumgartner, W. Neutral glycans from sandfish skin can reduce friction of polymers. J. R. Soc. Interface 2016, 13, 20160103. [Google Scholar] [CrossRef] [Green Version]
- Loch, N.; Geilen, C.C.; Spörndle, I.; Oberdorfer, F.; Keppler, D.; Tauber, R.; Reutter, W. 2-Deoxy-2-fluoro-D-galactose protein N-glycosylation. FEBS Lett. 1991, 294, 217–220. [Google Scholar] [CrossRef] [Green Version]
- Gross, V.; Hull, W.E.; Berger, U.; Andus, T.; Kreisel, W.; Gerok, W.; Keppler, D. Inhibition of protein N-glycosylation by 2-deoxy-2-fluoro-D-galactose. Biochem. J. 1992, 285, 821–826. [Google Scholar] [CrossRef]
- De-Souza, E.A.; Pimentel, F.S.A.; Machado, C.M.; Martins, L.S.; da-Silva, W.S.; Montero-Lomeli, M.; Masuda, C.A. The unfolded protein response has a protective role in yeast models of classic galactosemia. Dis. Models Mech. 2014, 7, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kushner, R.F.; Ryan, E.L.; Sefton, J.M.I.; Sanders, R.D.; Lucioni, P.J.; Moberg, K.H.; Fridovich-Keil, J.L. A Drosophila Melanogaster model of classic galactosemia. Dis. Models Mech. 2010, 3, 618–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, M.; Sidiqqi, A.; Witt, B.; Yuzyuk, T.; Johnson, B.; Fraser, N.; Chen, W.; Rascon, R.; Yin, X.; Goli, H.; et al. Subfertility and growth restriction in a new galactose-1-phosphate uridyltransferase (GALT)-deficient mouse model. Eur. J. Hum. Genet. 2014, 22, 1172–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, A.I.; Rubio-Gozalbo, M.E.; Vicente, J.B.; Rivera, I. Sweet and sour: An update on classic galactosemia. J. Inherit. Metab. Dis. 2017, 40, 325–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demetriades, C.; Doumpas, N.; Teleman, A.A. Regulation of TORC1 in response to amino acid starvation via lysosomal recruitment of TSC2. Cell 2014, 156, 786–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loewith, R.; Hall, M.N. Target of rapamycin (TOR) in nutrient signaling and growth control. Genetics 2011, 189, 1177–1201. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.W.; Jin, F.; Hwang, H.; Hwang, S.; Anand, V.; Duncan, M.C.; Huang, J. Insights into TOR function and rapamycin response: Chemical genomic profiling by using a high-density cell array method. Proc. Natl. Acad. Sci. USA 2005, 102, 7215–7220. [Google Scholar] [CrossRef] [Green Version]
- Mannaa, A.; Hanisch, F.-G. Redox proteomes in human physiology and disease mechanisms. J. Proteome Res. 2020, 19, 1–17. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecular Mass | Glycan Composition | Structure Model | Presence in Sample | |
|---|---|---|---|---|
| MNa/MH-32 | Control | F-Gal | ||
| 1580/1526 | H5N2 |  | ++ | ++ |
| 1866/1812 | H4N4 |  | + | + |
| 2040/1986 | F1H4N4 |  | + | + |
| 2070/2016 | H5N4 |  | ++ | ++ |
| 2244/2190 | F1H5N4 |  | +++ | +++ |
| 2285/2231 | F1H4N5 |  | ++ | +++ |
| 2431/2377 | S1H5N4 |  | +++ | ++ |
| 2605/2551 | S1F1H5N4 |  | ++ | ++ |
| 2967/2913 | S2F1H5N4 |  | (+) | (+) |
| 3055/3001 | S1F1H6N5 |  | (+) | (+) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janes, V.; Grabany, S.; Delbrouck, J.; Vincent, S.P.; Gottschalk, J.; Elling, L.; Hanisch, F.-G. Fluorinated Galactoses Inhibit Galactose-1-Phosphate Uridyltransferase and Metabolically Induce Galactosemia-like Phenotypes in HEK-293 Cells. Cells 2020, 9, 607. https://doi.org/10.3390/cells9030607
Janes V, Grabany S, Delbrouck J, Vincent SP, Gottschalk J, Elling L, Hanisch F-G. Fluorinated Galactoses Inhibit Galactose-1-Phosphate Uridyltransferase and Metabolically Induce Galactosemia-like Phenotypes in HEK-293 Cells. Cells. 2020; 9(3):607. https://doi.org/10.3390/cells9030607
Chicago/Turabian StyleJanes, Verena, Simona Grabany, Julien Delbrouck, Stephane P. Vincent, Johannes Gottschalk, Lothar Elling, and Franz-Georg Hanisch. 2020. "Fluorinated Galactoses Inhibit Galactose-1-Phosphate Uridyltransferase and Metabolically Induce Galactosemia-like Phenotypes in HEK-293 Cells" Cells 9, no. 3: 607. https://doi.org/10.3390/cells9030607