Preclinical Comparison of Stem Cells Secretome and Levodopa Application in a 6-Hydroxydopamine Rat Model of Parkinson’s Disease

 , and
, and 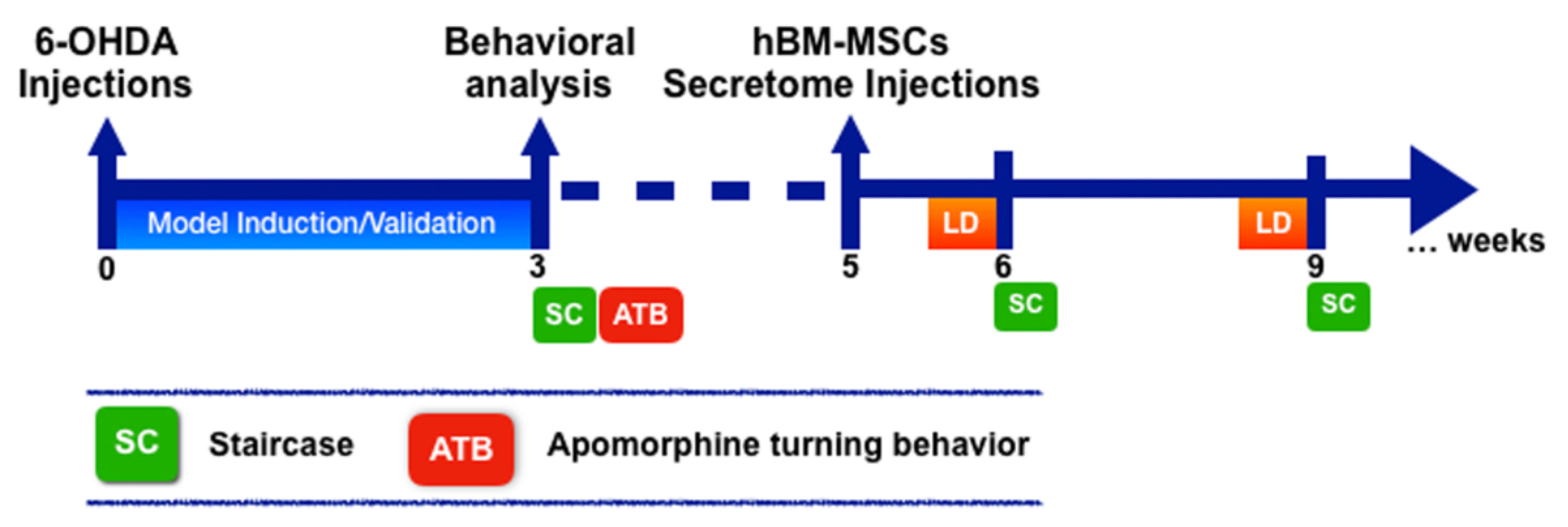{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
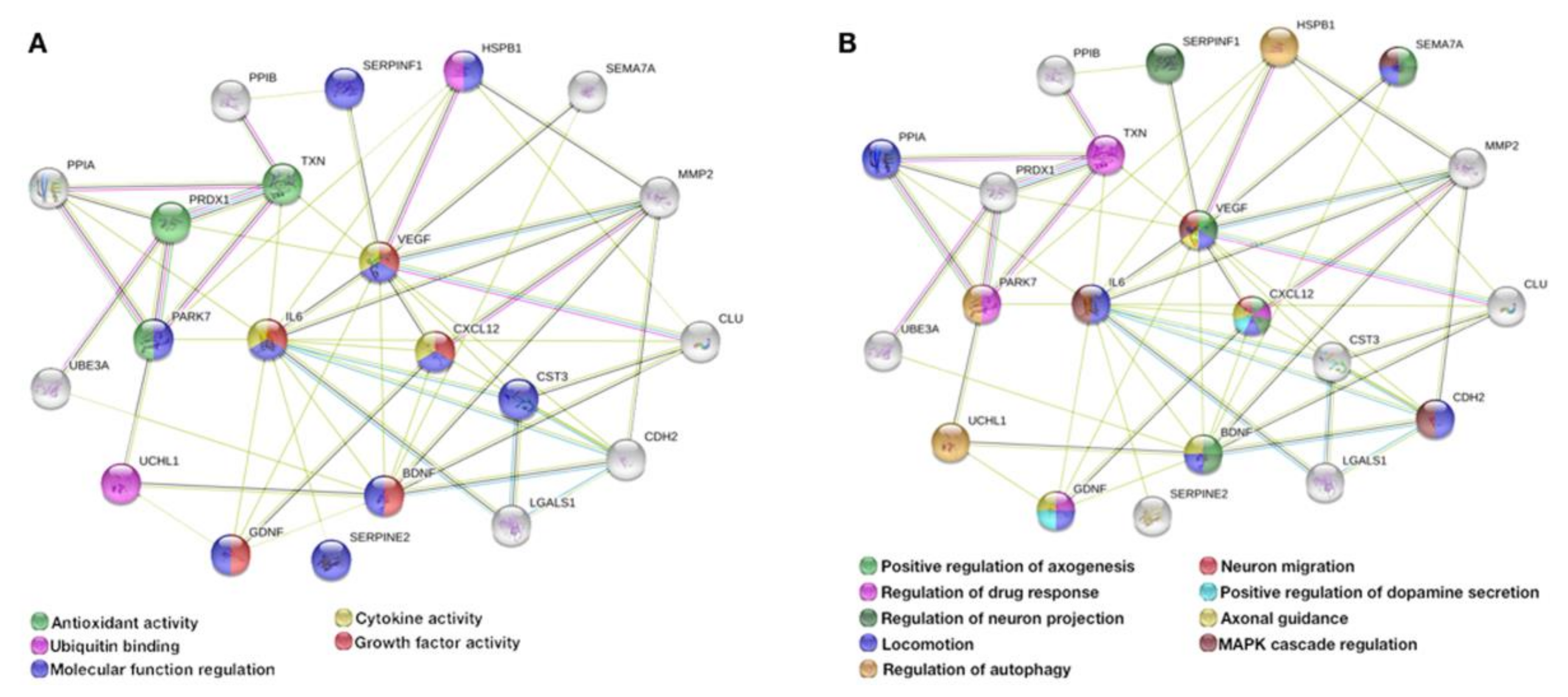
Abstract
:1. Introduction
2. Methods
2.1. Expansion of Human Bone Marrow MSCs (hBM-MSCs) and Secretome Collection
2.2. 6-Hydroxydopamine (6-OHDA)-Induced Lesions and hBM-MSCs Secretome and Levodopa Administration
2.3. Staircase Test
2.4. Apomorphine Turning Behavior
2.5. Tyrosine Hydroxylase (TH) Immunostaining and Striatal Fiber Density Measurement
2.6. Statistical Analysis
3. Results
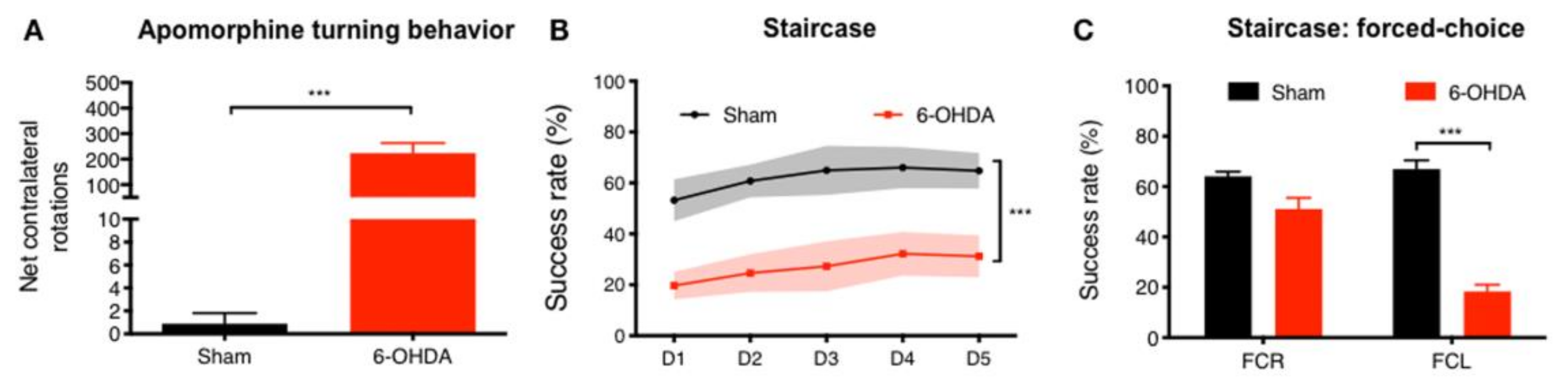
3.1. 6-OHDA PD Model Phenotypic Validation
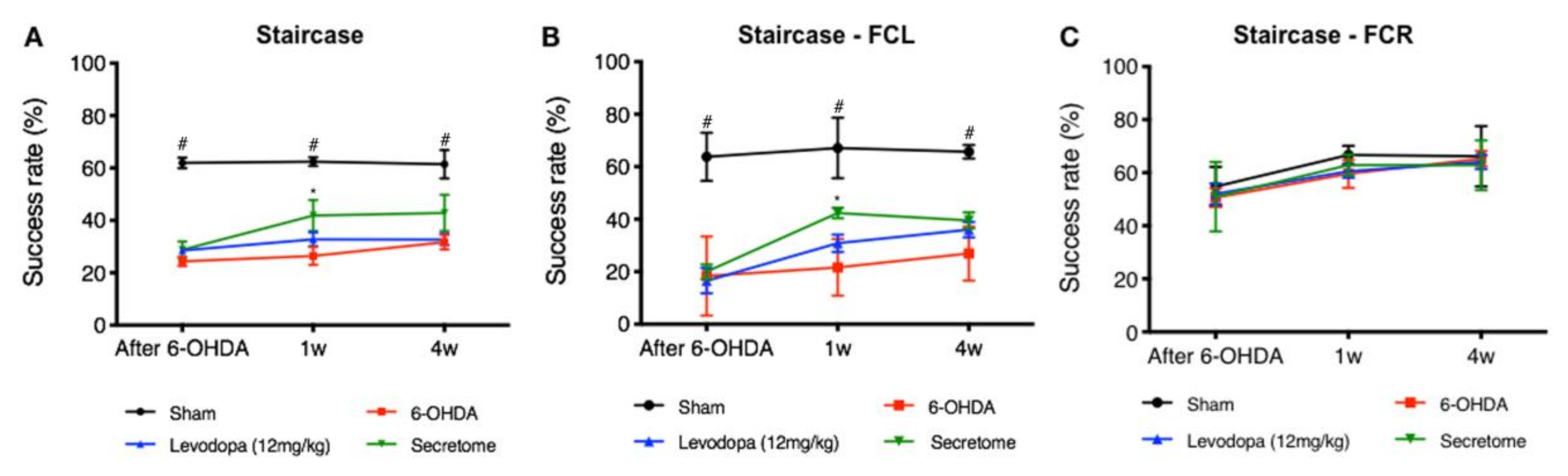
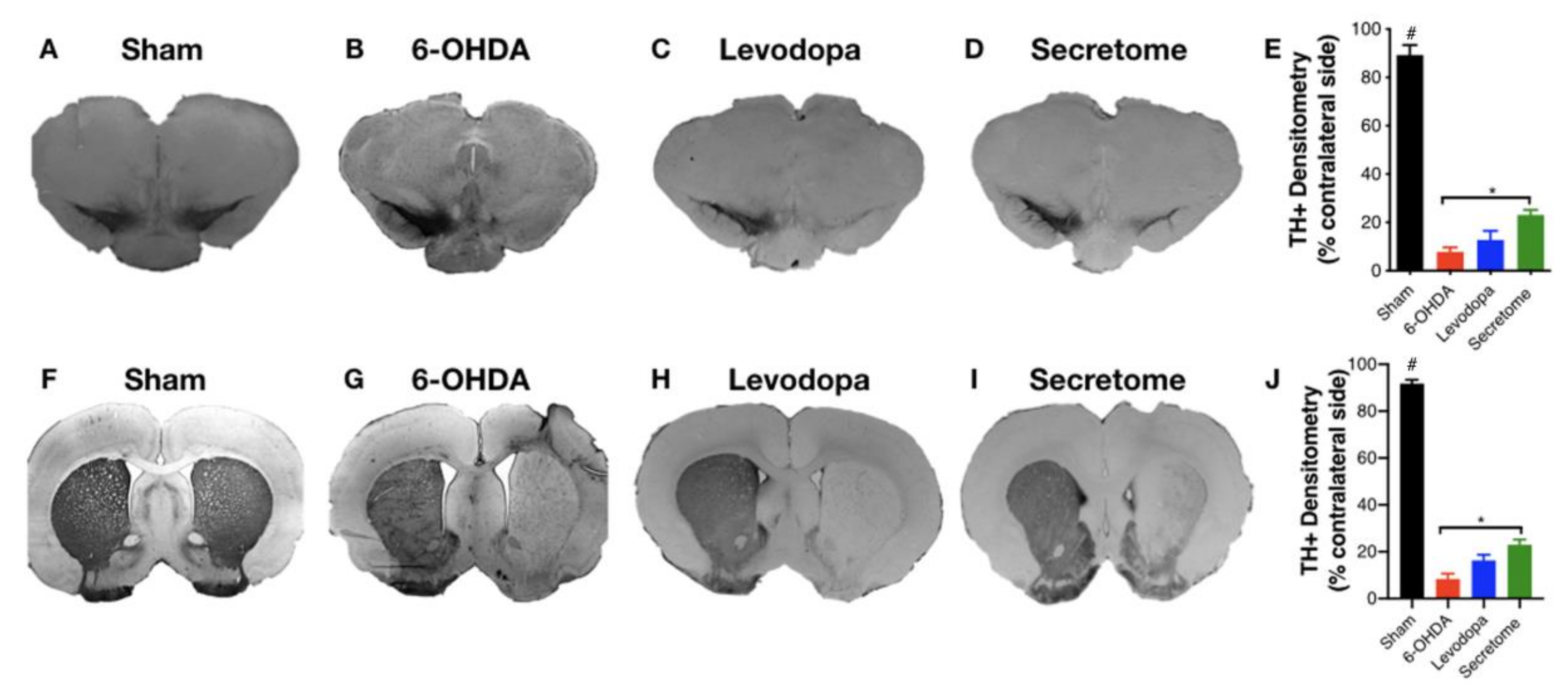
3.2. hBM-MSCs Secretome Improves 6-OHDA PD Motor and Histological Deficits
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Teixeira, F.G.; Carvalho, M.M.; Sousa, N.; Salgado, A.J. Mesenchymal stem cells secretome: A new paradigm for central nervous system regeneration? Cell. Mol. Life Sci. 2013, 70, 3871–3882. [Google Scholar] [CrossRef] [PubMed]
- Pires, A.O.; Teixeira, F.G.; Mendes-Pinheiro, B.; Serra, S.C.; Sousa, N.; Salgado, A.J. Old and New Challenges in Parkinson’s Disease Therapeutics. Prog. Neurobiol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Yacoubian, T.A.; Standaert, D.G. Targets for neuroprotection in Parkinson’s disease. Biochim. Biophys. Acta 2009, 1792, 676–687. [Google Scholar] [CrossRef] [PubMed]
- Nomoto, M.; Nagai, M.; Nishikawa, N.; Ando, R.; Kagamiishi, Y.; Yano, K.; Saito, S.; Takeda, A. Pharmacokinetics and safety/efficacy of levodopa pro-drug ONO-2160/carbidopa for Parkinson’s disease. eNeurologicalSci 2018, 13, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Pillay, V.; Choonara, Y.E. Advances in the treatment of Parkinson’s disease. Prog. Neurobiol. 2007, 81, 29–44. [Google Scholar] [CrossRef]
- Maiti, P.; Manna, J.; Dunbar, G.L. Current understanding of the molecular mechanisms in Parkinson’s disease: Targets for potential treatments. Transl. Neurodegener. 2017, 6, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, T.; Pu, J.; Si, X.; Ye, R.; Zhang, B. An update on potential therapeutic strategies for Parkinson’s disease based on pathogenic mechanisms. Expert. Rev. Neurother. 2016, 16, 711–722. [Google Scholar] [CrossRef]
- Fox, S.H.; Lang, A.E. ‘Don’t delay, start today’: Delaying levodopa does not delay motor complications. Brain 2014, 137, 2628–2630. [Google Scholar] [CrossRef] [Green Version]
- Blandini, F.; Cova, L.; Armentero, M.T.; Zennaro, E.; Levandis, G.; Bossolasco, P.; Calzarossa, C.; Mellone, M.; Giuseppe, B.; Deliliers, G.L.; et al. Transplantation of undifferentiated human mesenchymal stem cells protects against 6-hydroxydopamine neurotoxicity in the rat. Cell Transplant. 2010, 19, 203–217. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Squillaro, T.; Peluso, G.; Galderisi, U. Clinical Trials With Mesenchymal Stem Cells: An Update. Cell Transplant 2016, 25, 829–848. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, F.G.; Salgado, A.J. Mesenchymal stem cells secretome: Current trends and future challenges. Neural Regen. Res. 2020, 15, 75–77. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.R.; Teixeira, G.Q.; Santos, S.G.; Barbosa, M.A.; Almeida-Porada, G.; Goncalves, R.M. Mesenchymal Stromal Cell Secretome: Influencing Therapeutic Potential by Cellular Pre-conditioning. Front. Immunol. 2018, 9, 2837. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, F.G.; Carvalho, M.M.; Neves-Carvalho, A.; Panchalingam, K.M.; Behie, L.A.; Pinto, L.; Sousa, N.; Salgado, A.J. Secretome of mesenchymal progenitors from the umbilical cord acts as modulator of neural/glial proliferation and differentiation. Stem Cell Rev. 2015, 11, 288–297. [Google Scholar] [CrossRef]
- Teixeira, F.G.; Panchalingam, K.M.; Assuncao-Silva, R.; Serra, S.C.; Mendes-Pinheiro, B.; Patricio, P.; Jung, S.; Anjo, S.I.; Manadas, B.; Pinto, L.; et al. Modulation of the Mesenchymal Stem Cell Secretome Using Computer-Controlled Bioreactors: Impact on Neuronal Cell Proliferation, Survival and Differentiation. Sci. Rep. 2016, 6, 27791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, F.G.; Carvalho, M.M.; Panchalingam, K.M.; Rodrigues, A.J.; Mendes-Pinheiro, B.; Anjo, S.; Manadas, B.; Behie, L.A.; Sousa, N.; Salgado, A.J. Impact of the Secretome of Human Mesenchymal Stem Cells on Brain Structure and Animal Behavior in a Rat Model of Parkinson’s Disease. Stem Cells Transl. Med. 2017, 6, 634–646. [Google Scholar] [CrossRef]
- Carvalho, M.M.; Campos, F.L.; Coimbra, B.; Pego, J.M.; Rodrigues, C.; Lima, R.; Rodrigues, A.J.; Sousa, N.; Salgado, A.J. Behavioral characterization of the 6-hydroxidopamine model of Parkinson’s disease and pharmacological rescuing of non-motor deficits. Mol. Neurodegener. 2013, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Paxinos, G.; Watson, C. Rat Brain in Stereotaxic Coordinates, 5th ed.; Academic Press: San Diego, CA, USA, 2004. [Google Scholar]
- Carvalho, M.M.; Campos, F.L.; Marques, M.; Soares-Cunha, C.; Kokras, N.; Dalla, C.; Leite-Almeida, H.; Sousa, N.; Salgado, A.J. Effect of Levodopa on Reward and Impulsivity in a Rat Model of Parkinson’s Disease. Front. Behav. Neurosci. 2017, 11, 145. [Google Scholar] [CrossRef] [Green Version]
- Campos, F.L.; Carvalho, M.M.; Cristovao, A.C.; Je, G.; Baltazar, G.; Salgado, A.J.; Kim, Y.S.; Sousa, N. Rodent models of Parkinson’s disease: Beyond the motor symptomatology. Front. Behav. Neurosci. 2013, 7, 175. [Google Scholar] [CrossRef] [Green Version]
- Bibbiani, F.; Costantini, L.C.; Patel, R.; Chase, T.N. Continuous dopaminergic stimulation reduces risk of motor complications in parkinsonian primates. Exp. Neurol. 2005, 192, 73–78. [Google Scholar] [CrossRef]
- Poewe, W.; Wenning, G.K. Apomorphine: An underutilized therapy for Parkinson’s disease. Mov. Disord. Off. J. Mov. Disord. Soc. 2000, 15, 789–794. [Google Scholar] [CrossRef]
- Trenkwalder, C.; Chaudhuri, K.R.; Garcia Ruiz, P.J.; LeWitt, P.; Katzenschlager, R.; Sixel-Doring, F.; Henriksen, T.; Sesar, A.; Poewe, W. Expert Consensus Group report on the use of apomorphine in the treatment of Parkinson’s disease—Clinical practice recommendations. Parkinsonism Relat. Disord. 2015, 21, 1023–1030. [Google Scholar] [CrossRef]
- Febbraro, F.; Andersen, K.J.; Sanchez-Guajardo, V.; Tentillier, N.; Romero-Ramos, M. Chronic intranasal deferoxamine ameliorates motor defects and pathology in the alpha-synuclein rAAV Parkinson’s model. Exp. Neurol. 2013, 247, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Vilaca-Faria, H.; Salgado, A.J.; Teixeira, F.G. Mesenchymal Stem Cells-derived Exosomes: A New Possible Therapeutic Strategy for Parkinson’s Disease? Cells 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Mendes-Pinheiro, B.; Teixeira, F.G.; Anjo, S.I.; Manadas, B.; Behie, L.A.; Salgado, A.J. Secretome of Undifferentiated Neural Progenitor Cells Induces Histological and Motor Improvements in a Rat Model of Parkinson’s Disease. Stem Cells Transl. Med. 2018, 7, 829–838. [Google Scholar] [CrossRef] [Green Version]
- Pires, A.O.; Mendes-Pinheiro, B.; Teixeira, F.G.; Anjo, S.I.; Ribeiro-Samy, S.; Gomes, E.D.; Serra, S.C.; Silva, N.A.; Manadas, B.; Sousa, N.; et al. Unveiling the Differences of Secretome of Human Bone Marrow Mesenchymal Stem Cells, Adipose Tissue-Derived Stem Cells, and Human Umbilical Cord Perivascular Cells: A Proteomic Analysis. Stem Cells Dev. 2016, 25, 1073–1083. [Google Scholar] [CrossRef] [PubMed]
- Maulik, N.; Das, D.K. Emerging potential of thioredoxin and thioredoxin interacting proteins in various disease conditions. Biochim. Biophys. Acta 2008, 1780, 1368–1382. [Google Scholar] [CrossRef] [PubMed]
- Arodin, L.; Miranda-Vizuete, A.; Swoboda, P.; Fernandes, A.P. Protective effects of the thioredoxin and glutaredoxin systems in dopamine-induced cell death. Free Radic. Biol. Med. 2014, 73, 328–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, X.S.; Jia, J.J.; Kwon, Y.; Wang, S.D.; Bai, J. The role of thioredoxin-1 in suppression of endoplasmic reticulum stress in Parkinson disease. Free Radic. Biol. Med. 2014, 67, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Silva-Adaya, D.; Gonsebatt, M.E.; Guevara, J. Thioredoxin system regulation in the central nervous system: Experimental models and clinical evidence. Oxid. Med. Cell Longev. 2014, 2014, 590808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasterkamp, R.J.; Peschon, J.J.; Spriggs, M.K.; Kolodkin, A.L. Semaphorin 7A promotes axon outgrowth through integrins and MAPKs. Nature 2003, 424, 398–405. [Google Scholar] [CrossRef]
- Kalaani, J.; Roche, J.; Hamade, E.; Badran, B.; Jaber, M.; Gaillard, A.; Prestoz, L. Axon guidance molecule expression after cell therapy in a mouse model of Parkinson’s disease. Restor. Neurol. Neurosci. 2016, 34, 877–895. [Google Scholar] [CrossRef] [PubMed]
- Pacelli, C.; Giguere, N.; Bourque, M.J.; Levesque, M.; Slack, R.S.; Trudeau, L.E. Elevated Mitochondrial Bioenergetics and Axonal Arborization Size Are Key Contributors to the Vulnerability of Dopamine Neurons. Curr. Biol. 2015, 25, 2349–2360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardona, F.; Perez-Tur, J. Other Proteins Involved in Parkinson’s Disease and Related Disorders. Curr. Protein Pept. Sci. 2017, 18, 765–778. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Huo, H.; Yang, C.; Zhang, T.; Chu, Y.; Liu, Y. Ubiquitin C-Terminal Hydrolase L1 regulates autophagy by inhibiting autophagosome formation through its deubiquitinating enzyme activity. Biochem. Biophys. Res. Commun. 2018, 497, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Lunati, A.; Lesage, S.; Brice, A. The genetic landscape of Parkinson’s disease. Rev. Neurol. 2018, 174, 628–643. [Google Scholar] [CrossRef] [PubMed]
- Bifsha, P.; Landry, K.; Ashmarina, L.; Durand, S.; Seyrantepe, V.; Trudel, S.; Quiniou, C.; Chemtob, S.; Xu, Y.; Gravel, R.A.; et al. Altered gene expression in cells from patients with lysosomal storage disorders suggests impairment of the ubiquitin pathway. Cell Death Differ. 2007, 14, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.Y.; Zhou, H.Y.; Wang, Z.Q.; Chen, S.D. Endoplasmic reticulum stress contributes to the cell death induced by UCH-L1 inhibitor. Mol. Cell. Biochem. 2008, 318, 109–115. [Google Scholar] [CrossRef]
- Falk, T.; Gonzalez, R.T.; Sherman, S.J. The yin and yang of VEGF and PEDF: Multifaceted neurotrophic factors and their potential in the treatment of Parkinson’s Disease. Int. J. Mol. Sci. 2010, 11, 2875–2900. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, T.; Fukuda-Tani, M.; Nihira, T.; Wada, K.; Hattori, N.; Mizuno, Y.; Mochizuki, H. Correlation between levels of pigment epithelium-derived factor and vascular endothelial growth factor in the striatum of patients with Parkinson’s disease. Exp. Neurol. 2007, 206, 308–317. [Google Scholar] [CrossRef]
- Allen, S.J.; Watson, J.J.; Shoemark, D.K.; Barua, N.U.; Patel, N.K. GDNF, NGF and BDNF as therapeutic options for neurodegeneration. Pharmacol. Ther. 2013, 138, 155–175. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Chen, W.; Liu, X.; Qiao, D.; Zhou, F.M. Exercise-Induced Neuroprotection of the Nigrostriatal Dopamine System in Parkinson’s Disease. Front. Aging Neurosci. 2017, 9, 358. [Google Scholar] [CrossRef] [Green Version]
- Fujii, H.; Matsubara, K.; Sakai, K.; Ito, M.; Ohno, K.; Ueda, M.; Yamamoto, A. Dopaminergic differentiation of stem cells from human deciduous teeth and their therapeutic benefits for Parkinsonian rats. Brain Res. 2015, 1613, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Baquet, Z.C.; Bickford, P.C.; Jones, K.R. Brain-derived neurotrophic factor is required for the establishment of the proper number of dopaminergic neurons in the substantia nigra pars compacta. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 6251–6259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Edman, L.C.; Sanchez-Alcaniz, J.A.; Fritz, N.; Bonilla, S.; Hecht, J.; Uhlen, P.; Pleasure, S.J.; Villaescusa, J.C.; Marin, O.; et al. Cxcl12/Cxcr4 signaling controls the migration and process orientation of A9-A10 dopaminergic neurons. Development 2013, 140, 4554–4564. [Google Scholar] [CrossRef] [Green Version]
- Skrzydelski, D.; Guyon, A.; Dauge, V.; Rovere, C.; Apartis, E.; Kitabgi, P.; Nahon, J.L.; Rostene, W.; Parsadaniantz, S.M. The chemokine stromal cell-derived factor-1/CXCL12 activates the nigrostriatal dopamine system. J. Neurochem. 2007, 102, 1175–1183. [Google Scholar] [CrossRef]
- Kim, H.; Oh, Y.; Kim, K.; Jeong, S.; Chon, S.; Kim, D.; Jung, M.H.; Pak, Y.K.; Ha, J.; Kang, I.; et al. Cyclophilin A regulates JNK/p38-MAPK signaling through its physical interaction with ASK1. Biochem. Biophys. Res. Commun. 2015, 464, 112–117. [Google Scholar] [CrossRef]
- Liu, J.; Mu, C.; Yue, W.; Li, J.; Ma, B.; Zhao, L.; Liu, L.; Chen, Q.; Yan, C.; Liu, H.; et al. A diterpenoid derivate compound targets selenocysteine of thioredoxin reductases and induces Bax/Bak-independent apoptosis. Free Radic. Biol. Med. 2013, 63, 485–494. [Google Scholar] [CrossRef]
- Mathews, P.M.; Levy, E. Cystatin C in aging and in Alzheimer’s disease. Ageing Res. Rev. 2016, 32, 38–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Gonzalez, R.; Sahoo, S.; Gauthier, S.A.; Kim, Y.; Li, M.; Kumar, A.; Pawlik, M.; Benussi, L.; Ghidoni, R.; Levy, E. Neuroprotection mediated by cystatin C-loaded extracellular vesicles. Sci. Rep. 2019, 9, 11104. [Google Scholar] [CrossRef] [Green Version]
- Zou, J.; Chen, Z.; Wei, X.; Chen, Z.; Fu, Y.; Yang, X.; Chen, D.; Wang, R.; Jenner, P.; Lu, J.H.; et al. Cystatin C as a potential therapeutic mediator against Parkinson’s disease via VEGF-induced angiogenesis and enhanced neuronal autophagy in neurovascular units. Cell Death Dis. 2017, 8, e2854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Sheng, J.; Tang, Z.; Wu, X.; Yu, Y.; Guo, H.; Shen, Y.; Zhou, C.; Paraoan, L.; Zhou, J. Cystatin C prevents degeneration of rat nigral dopaminergic neurons: In vitro and in vivo studies. Neurobiol. Dis. 2005, 18, 152–165. [Google Scholar] [CrossRef] [PubMed]
- Yasuhara, T.; Shingo, T.; Muraoka, K.; Kameda, M.; Agari, T.; Wen Ji, Y.; Hayase, H.; Hamada, H.; Borlongan, C.V.; Date, I. Neurorescue effects of VEGF on a rat model of Parkinson’s disease. Brain Res. 2005, 1053, 10–18. [Google Scholar] [CrossRef]
- Repici, M.; Giorgini, F. DJ-1 in Parkinson’s Disease: Clinical Insights and Therapeutic Perspectives. J. Clin. Med. 2019, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, W.M.; Golczak, M.; Choe, K.; Curran, P.L.; Miller, O.G.; Yao, C.; Wang, W.; Lin, J.; Milkovic, N.M.; Ray, A.; et al. Regulation of DJ-1 by Glutaredoxin 1 in Vivo: Implications for Parkinson’s Disease. Biochemistry 2016, 55, 4519–4532. [Google Scholar] [CrossRef] [Green Version]
- Chien, C.H.; Lee, M.J.; Liou, H.C.; Liou, H.H.; Fu, W.M. Microglia-Derived Cytokines/Chemokines Are Involved in the Enhancement of LPS-Induced Loss of Nigrostriatal Dopaminergic Neurons in DJ-1 Knockout Mice. PLoS ONE 2016, 11, e0151569. [Google Scholar] [CrossRef]
- Chen, R.; Park, H.A.; Mnatsakanyan, N.; Niu, Y.; Licznerski, P.; Wu, J.; Miranda, P.; Graham, M.; Tang, J.; Boon, A.J.W.; et al. Parkinson’s disease protein DJ-1 regulates ATP synthase protein components to increase neuronal process outgrowth. Cell Death Dis. 2019, 10, 469. [Google Scholar] [CrossRef] [Green Version]
- Zondler, L.; Miller-Fleming, L.; Repici, M.; Goncalves, S.; Tenreiro, S.; Rosado-Ramos, R.; Betzer, C.; Straatman, K.R.; Jensen, P.H.; Giorgini, F.; et al. DJ-1 interactions with alpha-synuclein attenuate aggregation and cellular toxicity in models of Parkinson’s disease. Cell Death Dis. 2014, 5, e1350. [Google Scholar] [CrossRef] [Green Version]
- D’Anglemont de Tassigny, X.; Pascual, A.; Lopez-Barneo, J. GDNF-based therapies, GDNF-producing interneurons, and trophic support of the dopaminergic nigrostriatal pathway. Implications for Parkinson’s disease. Front. Neuroanat. 2015, 9, 10. [Google Scholar] [CrossRef] [Green Version]
- Migliore, M.M.; Ortiz, R.; Dye, S.; Campbell, R.B.; Amiji, M.M.; Waszczak, B.L. Neurotrophic and neuroprotective efficacy of intranasal GDNF in a rat model of Parkinson’s disease. Neuroscience 2014, 274, 11–23. [Google Scholar] [CrossRef]
- Garea-Rodriguez, E.; Eesmaa, A.; Lindholm, P.; Schlumbohm, C.; Konig, J.; Meller, B.; Krieglstein, K.; Helms, G.; Saarma, M.; Fuchs, E. Comparative Analysis of the Effects of Neurotrophic Factors CDNF and GDNF in a Nonhuman Primate Model of Parkinson’s Disease. PLoS ONE 2016, 11, e0149776. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.C.; Lee, E.H. Neuroprotective mechanism of glial cell line-derived neurotrophic factor on dopamine neurons: Role of antioxidation. Neuropharmacology 1999, 38, 913–916. [Google Scholar] [CrossRef]
- Sullivan, A.M.; O’Keeffe, G.W. Neurotrophic factor therapy for Parkinson’s disease: Past, present and future. Neural. Regen. Res. 2016, 11, 205–207. [Google Scholar] [CrossRef] [PubMed]
- Zuo, T.; Qin, J.Y.; Chen, J.; Shi, Z.; Liu, M.; Gao, X.; Gao, D. Involvement of N-cadherin in the protective effect of glial cell line-derived neurotrophic factor on dopaminergic neuron damage. Int. J. Mol. Med. 2013, 31, 561–568. [Google Scholar] [CrossRef] [Green Version]
- Sakane, F.; Miyamoto, Y. N-cadherin regulates the proliferation and differentiation of ventral midbrain dopaminergic progenitors. Dev. Neurobiol. 2013, 73, 518–529. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Ishihara, K.; Hibi, M. Roles of STAT3 in mediating the cell growth, differentiation and survival signals relayed through the IL-6 family of cytokine receptors. Oncogene 2000, 19, 2548–2556. [Google Scholar] [CrossRef] [PubMed]
- Erta, M.; Quintana, A.; Hidalgo, J. Interleukin-6, a major cytokine in the central nervous system. Int. J. Biol. Sci. 2012, 8, 1254–1266. [Google Scholar] [CrossRef]
- Bolin, L.M.; Strycharska-Orczyk, I.; Murray, R.; Langston, J.W.; Di Monte, D. Increased vulnerability of dopaminergic neurons in MPTP-lesioned interleukin-6 deficient mice. J. Neurochem. 2002, 83, 167–175. [Google Scholar] [CrossRef]
- Akaneya, Y.; Takahashi, M.; Hatanaka, H. Interleukin-1 beta enhances survival and interleukin-6 protects against MPP+ neurotoxicity in cultures of fetal rat dopaminergic neurons. Exp. Neurol. 1995, 136, 44–52. [Google Scholar] [CrossRef]
- Lee, Y.P.; Kim, D.W.; Kang, H.W.; Hwang, J.H.; Jeong, H.J.; Sohn, E.J.; Kim, M.J.; Ahn, E.H.; Shin, M.J.; Kim, D.S.; et al. PEP-1-heat shock protein 27 protects from neuronal damage in cells and in a Parkinson’s disease mouse model. FEBS J. 2012, 279, 1929–1942. [Google Scholar] [CrossRef]
- Cox, D.; Whiten, D.R.; Brown, J.W.P.; Horrocks, M.H.; San Gil, R.; Dobson, C.M.; Klenerman, D.; van Oijen, A.M.; Ecroyd, H. The small heat shock protein Hsp27 binds alpha-synuclein fibrils, preventing elongation and cytotoxicity. J. Biol. Chem. 2018, 293, 4486–4497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.M.; Park, S.H.; Shin, D.I.; Hwang, J.Y.; Park, B.; Park, Y.J.; Lee, T.H.; Chae, H.Z.; Jin, B.K.; Oh, T.H.; et al. Oxidative modification of peroxiredoxin is associated with drug-induced apoptotic signaling in experimental models of Parkinson disease. J. Biol. Chem. 2008, 283, 9986–9998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goemaere, J.; Knoops, B. Peroxiredoxin distribution in the mouse brain with emphasis on neuronal populations affected in neurodegenerative disorders. J. Comp. Neurol. 2012, 520, 258–280. [Google Scholar] [CrossRef]
- Farmer, L.; Sommer, J.; Monard, D. Glia-derived nexin potentiates neurite extension in hippocampal pyramidal cells in vitro. Dev. Neurosci. 1990, 12, 73–80. [Google Scholar] [CrossRef]
- Hoffmann, M.C.; Nitsch, C.; Scotti, A.L.; Reinhard, E.; Monard, D. The prolonged presence of glia-derived nexin, an endogenous protease inhibitor, in the hippocampus after ischemia-induced delayed neuronal death. Neuroscience 1992, 49, 397–408. [Google Scholar] [CrossRef]
- Pucci, S.; Mazzarelli, P.; Missiroli, F.; Regine, F.; Ricci, F. Neuroprotection: VEGF, IL-6, and clusterin: The dark side of the moon. Prog. Brain Res. 2008, 173, 555–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, Y.; Jeong, K.; Kim, K.; Lee, Y.S.; Jeong, S.; Kim, S.S.; Yoon, K.S.; Ha, J.; Kang, I.; Choe, W. Cyclophilin B protects SH-SY5Y human neuroblastoma cells against MPP( + )-induced neurotoxicity via JNK pathway. Biochem. Biophys. Res. Commun. 2016, 478, 1396–1402. [Google Scholar] [CrossRef]
- Starossom, S.C.; Mascanfroni, I.D.; Imitola, J.; Cao, L.; Raddassi, K.; Hernandez, S.F.; Bassil, R.; Croci, D.O.; Cerliani, J.P.; Delacour, D.; et al. Galectin-1 deactivates classically activated microglia and protects from inflammation-induced neurodegeneration. Immunity 2012, 37, 249–263. [Google Scholar] [CrossRef] [Green Version]
- Mulherkar, S.A.; Jana, N.R. Loss of dopaminergic neurons and resulting behavioural deficits in mouse model of Angelman syndrome. Neurobiol. Dis. 2010, 40, 586–592. [Google Scholar] [CrossRef]
- Oh, S.H.; Kim, H.N.; Park, H.J.; Shin, J.Y.; Kim, D.Y.; Lee, P.H. The Cleavage Effect of Mesenchymal Stem Cell and Its Derived Matrix Metalloproteinase-2 on Extracellular alpha-Synuclein Aggregates in Parkinsonian Models. Stem Cells Transl. Med. 2017, 6, 949–961. [Google Scholar] [CrossRef]
- Colamartino, M.; Santoro, M.; Duranti, G.; Sabatini, S.; Ceci, R.; Testa, A.; Padua, L.; Cozzi, R. Evaluation of levodopa and carbidopa antioxidant activity in normal human lymphocytes in vitro: Implication for oxidative stress in Parkinson’s disease. Neurotox. Res. 2015, 27, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Thirugnanasambandam, N.; Grundey, J.; Paulus, W.; Nitsche, M.A. Dose-dependent nonlinear effect of L-DOPA on paired associative stimulation-induced neuroplasticity in humans. J. Neurosci. 2011, 31, 5294–5299. [Google Scholar] [CrossRef]
- Brod, L.S.; Aldred, J.L.; Nutt, J.G. Are high doses of carbidopa a concern? A randomized, clinical trial in Parkinson’s disease. Mov. Disord. 2012, 27, 750–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tayarani-Binazir, K.A.; Jackson, M.J.; Fisher, R.; Zoubiane, G.; Rose, S.; Jenner, P. The timing of administration, dose dependence and efficacy of dopa decarboxylase inhibitors on the reversal of motor disability produced by L-DOPA in the MPTP-treated common marmoset. Eur. J. Pharm. 2010, 635, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Picconi, B.; Paille, V.; Ghiglieri, V.; Bagetta, V.; Barone, I.; Lindgren, H.S.; Bernardi, G.; Angela Cenci, M.; Calabresi, P. l-DOPA dosage is critically involved in dyskinesia via loss of synaptic depotentiation. Neurobiol. Dis. 2008, 29, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, F.G.; Gago, M.F.; Marques, P.; Moreira, P.S.; Magalhaes, R.; Sousa, N.; Salgado, A.J. Safinamide: A new hope for Parkinson’s disease? Drug Discov. Today 2018. [Google Scholar] [CrossRef]
- Fabbri, M.; Coelho, M.; Abreu, D.; Guedes, L.C.; Rosa, M.M.; Costa, N.; Antonini, A.; Ferreira, J.J. Do patients with late-stage Parkinson’s disease still respond to levodopa? Parkinsonism Relat. Disord. 2016, 26, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Calabresi, P.; Ghiglieri, V.; Mazzocchetti, P.; Corbelli, I.; Picconi, B. Levodopa-induced plasticity: A double-edged sword in Parkinson’s disease? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370. [Google Scholar] [CrossRef] [Green Version]
- Lane, E.L. L-DOPA for Parkinson’s disease-a bittersweet pill. Eur. J. Neurosci. 2019, 49, 384–398. [Google Scholar] [CrossRef]
- Cooper, J.F.; Van Raamsdonk, J.M. Modeling Parkinson’s Disease in C. elegans. J. Parkinsons Dis. 2018, 8, 17–32. [Google Scholar] [CrossRef] [Green Version]
- Van der Schyf, C.J.; Geldenhuys, W.J. Multimodal drugs and their future for Alzheimer’s and Parkinson’s disease. Int. Rev. Neurobiol. 2011, 100, 107–125. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teixeira, F.G.; Vilaça-Faria, H.; Domingues, A.V.; Campos, J.; Salgado, A.J. Preclinical Comparison of Stem Cells Secretome and Levodopa Application in a 6-Hydroxydopamine Rat Model of Parkinson’s Disease. Cells 2020, 9, 315. https://doi.org/10.3390/cells9020315
Teixeira FG, Vilaça-Faria H, Domingues AV, Campos J, Salgado AJ. Preclinical Comparison of Stem Cells Secretome and Levodopa Application in a 6-Hydroxydopamine Rat Model of Parkinson’s Disease. Cells. 2020; 9(2):315. https://doi.org/10.3390/cells9020315
Chicago/Turabian StyleTeixeira, Fábio G., Helena Vilaça-Faria, Ana V. Domingues, Jonas Campos, and António J. Salgado. 2020. "Preclinical Comparison of Stem Cells Secretome and Levodopa Application in a 6-Hydroxydopamine Rat Model of Parkinson’s Disease" Cells 9, no. 2: 315. https://doi.org/10.3390/cells9020315