Peripheral Neuropathic Pain: From Experimental Models to Potential Therapeutic Targets in Dorsal Root Ganglion Neurons

Abstract
:1. Introduction
1.1. Overview of Neuropathic Pain
1.2. Etiology and Patholoigcal Approches
2. Neuropathic Pain Models
2.1. Injury Induced
2.2. Systemic Exposure Neuropathic Pain Model
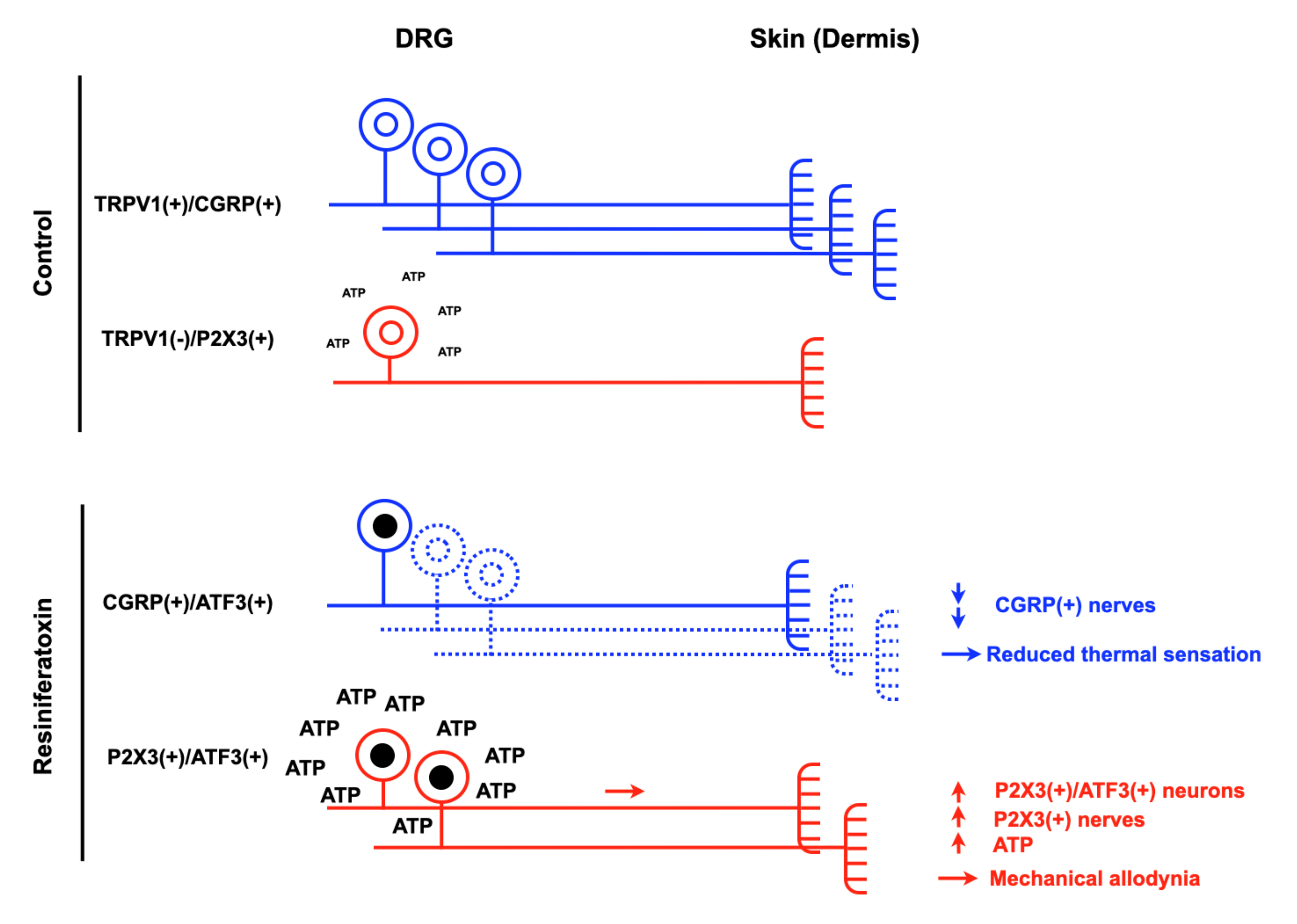
2.2.1. Resiniferatoxin
2.2.2. Acrylamide
2.2.3. Chemotherapy-Induced Peripheral Neuropathy (CIPN)
3. Molecular Targets in Dorsal Root Ganglia
3.1. Purinergic P2X Receptors (P2Xs)
3.1.1. Transient Receptor Potential Receptors
3.1.2. Voltage-Gated Sodium Channels (NaVs)
3.1.3. Voltage-Gated Calcium Channels
4. Epigenetic, Transcriptional, and Post-Translational Modification in Neuropathic Pain
4.1. DNA Methyltransferase (DNAMT) and Histone Deacetylase (HDAC)
4.1.1. DNA Methylation and Demethylation
4.1.2. Histone Deacetylase Inhibitors
4.2. p-ERK and Transcriptional Factors
4.3. Ubiquitination and Deubiquitination
5. Current Challenges to Manipulate on DRG in In Vivo Models
6. Conclusions

{kind=link}
| DRG | P2Xs | TRPs | NaVs | CaVs |
|---|---|---|---|---|
| Injury-Induced | ||||
| SNL | P2X3 ↑ (IR: d7, mRNA: d1) [55,56,57] P2X3 ↓ (IR: d14, mRNA: d7) [55,57] P2X3—(IR: d3, mRNA: d14) [56] P2X4—(mRNA: d1, d7) [57] P2X5 ↑ (mRNA: d1, d7) [57] P2X6 ↓ (mRNA: d7) [57] P2X6—(mRNA: d1) [57] | TRPV1 ↑ (IR: d3, mRNA: d1-28) [56] TRPA1 ↑ (IR: d11, mRNA) [131] | NaV1.7 ↓ (L5, IR: d7, mRNA: d7) [132] NaV1.7 ↑ (L4, IR: d7, mRNA: d7) [132] NaV1.3 ↑ [133] NaV1.1 ↓ [133] NaV1.6 ↓ [133] NaV1.7 ↓ [133] NaV1.8 ↓ [133] NaV1.9 ↓ [133] | α2δ-1 subunit ↑ (mRNA: d2) [81] |
| CCI | P2X2—(IR: d18) [134] P2X2 ↑ (mRNA: d14) [135] P2X3 ↑ (IR: d14, d18, mRNA: d14) [51,52,53,58,135] P2X4—(IR: d18) [134] P2X4 ↓ (IR:d18, mRNA: d14) [51,53] P2X5—(mRNA: d14) [134] P2X6—(IR: d18) [134] P2X6 ↑ (IR: d18) [53] | TRPM8 ↑ (IR: d4, d7, d10, d14, mRNA: d14) [64,136] TPRMP—(mRNA: d7) [136] TRPA1 ↑ (mRNA: d7, d14, d16) [136,137] TRPV1 ↑ (mRNA: d16) [137] TRPV1—(mRNA: d7, d14) [136] | NaV1.7 ↑ (IR: d7, mRNA: d7) [132] | α 1 ↓ (mRNA: d7) [82] |
| SNI | P2X3—(IR: d7) [135] | TRPM8 ↓ [138] TRPV1 ↓ [138] TRPA1 ↓ [138] | Rats (protein) [139] NaV1.3 ↑ NaV1.7 ↓ NaV1.8 ↓ NaV1.9 ↓ Mice (mRNA) [140] NaV1.1 ↓ NaV1.2 − NaV1.3 ↓ NaV1.6 ↓ NaV1.7 ↓ NaV1.8 ↓ NaV1.9 ↓ | Cav3.2 ↑ [80] |
| DRG | P2Xs | TRPs | NaVs | CaVs |
|---|---|---|---|---|
| Systemic Small Fiber Neuropathy | ||||
| RTX | P2X3 ↑ (IR: d7) [59] | TRPV1 ↓ (mRNA: d3) [53,59,136] TRPA1 ↓ (mRNA: d3) [136,141] TRPM8—(mRNA: d3) [136] | N/A | N/A |
| Cisplatin | P2X3 ↑ (mRNA: d14) [60] | TRPV1 ↑ [142] TRPA1 ↑ [142] TRPM8—[142] | N/A | CaV2.2 ↑ (IR: d3) [143] |
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van Hecke, O.; Austin, S.K.; Khan, R.A.; Smith, B.H.; Torrance, N. Neuropathic pain in the general population: A systematic review of epidemiological studies. Pain 2014, 155, 654–662. [Google Scholar] [CrossRef]
- Colloca, L.; Ludman, T.; Bouhassira, D.; Baron, R.; Dickenson, A.H.; Yarnitsky, D.; Freeman, R.; Truini, A.; Attal, N.; Finnerup, N.B.; et al. Neuropathic pain. Nat. Rev. Dis. Primers 2017, 3, 17002. [Google Scholar] [CrossRef] [Green Version]
- Hansson, P.T.; Attal, N.; Baron, R.; Cruccu, G. Toward a definition of pharmacoresistant neuropathic pain. Eur. J. Pain 2009, 13, 439–440. [Google Scholar] [CrossRef]
- Omori, S.; Isose, S.; Misawa, S.; Watanabe, K.; Sekiguchi, Y.; Shibuya, K.; Beppu, M.; Amino, H.; Kuwabara, S. Pain-related evoked potentials after intraepidermal electrical stimulation to Adelta and C fibers in patients with neuropathic pain. Neurosci. Res. 2017, 121, 43–48. [Google Scholar] [CrossRef]
- Lacomis, D. Small-fiber neuropathy. Muscle Nerve 2002, 26, 173–188. [Google Scholar] [CrossRef]
- Abram, S.E.; Yi, J.; Fuchs, A.; Hogan, Q.H. Permeability of injured and intact peripheral nerves and dorsal root ganglia. Anesthesiology 2006, 105, 146–153. [Google Scholar] [CrossRef]
- Jacobs, J.M.; Macfarlane, R.M.; Cavanagh, J.B. Vascular leakage in the dorsal root ganglia of the rat, studied with horseradish peroxidase. J. Neurol. Sci. 1976, 29, 95–107. [Google Scholar] [CrossRef]
- Sapunar, D.; Ljubkovic, M.; Lirk, P.; McCallum, J.B.; Hogan, Q.H. Distinct membrane effects of spinal nerve ligation on injured and adjacent dorsal root ganglion neurons in rats. Anesthesiology 2005, 103, 360–376. [Google Scholar] [CrossRef]
- Chen, C.J.; Liu, D.Z.; Yao, W.F.; Gu, Y.; Huang, F.; Hei, Z.Q.; Li, X. Identification of key genes and pathways associated with neuropathic pain in uninjured dorsal root ganglion by using bioinformatic analysis. J. Pain Res. 2017, 10, 2665–2674. [Google Scholar] [CrossRef] [Green Version]
- Maratou, K.; Wallace, V.C.; Hasnie, F.S.; Okuse, K.; Hosseini, R.; Jina, N.; Blackbeard, J.; Pheby, T.; Orengo, C.; Dickenson, A.H.; et al. Comparison of dorsal root ganglion gene expression in rat models of traumatic and HIV-associated neuropathic pain. Eur. J. Pain 2009, 13, 387–398. [Google Scholar] [CrossRef] [Green Version]
- Komori, N.; Takemori, N.; Kim, H.K.; Singh, A.; Hwang, S.H.; Foreman, R.D.; Chung, K.; Chung, J.M.; Matsumoto, H. Proteomics study of neuropathic and nonneuropathic dorsal root ganglia: Altered protein regulation following segmental spinal nerve ligation injury. Physiol. Genom. 2007, 29, 215–230. [Google Scholar] [CrossRef] [Green Version]
- Puljak, L.; Kojundzic, S.L.; Hogan, Q.H.; Sapunar, D. Targeted delivery of pharmacological agents into rat dorsal root ganglion. J. Neurosci. Methods 2009, 177, 397–402. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.P.; Zhou, W.; Kang, L.M.; Yan, H.; Zhang, L.; Xu, B.H.; Cai, W.H. Intrathecal miR-96 inhibits Nav1.3 expression and alleviates neuropathic pain in rat following chronic construction injury. Neurochem. Res. 2014, 39, 76–83. [Google Scholar] [CrossRef]
- Xu, X.; Fu, S.; Shi, X.; Liu, R. Microglial BDNF, PI3K, and p-ERK in the Spinal Cord Are Suppressed by Pulsed Radiofrequency on Dorsal Root Ganglion to Ease SNI-Induced Neuropathic Pain in Rats. Pain Res. Manag. 2019, 2019, 5948686. [Google Scholar] [CrossRef]
- Wang, Y.; Xue, M.; Xia, Y.; Jiang, Q.; Huang, Z.; Huang, C. Electroacupuncture treatment upregulates alpha7nAChR and inhibits JAK2/STAT3 in dorsal root ganglion of rat with spared nerve injury. J. Pain Res. 2019, 12, 1947–1955. [Google Scholar] [CrossRef] [Green Version]
- Liem, L.; Russo, M.; Huygen, F.J.; Van Buyten, J.P.; Smet, I.; Verrills, P.; Cousins, M.; Brooker, C.; Levy, R.; Deer, T.; et al. One-year outcomes of spinal cord stimulation of the dorsal root ganglion in the treatment of chronic neuropathic pain. Neuromodulation 2015, 18, 41–48. [Google Scholar] [CrossRef]
- Van Buyten, J.P.; Smet, I.; Liem, L.; Russo, M.; Huygen, F. Stimulation of dorsal root ganglia for the management of complex regional pain syndrome: A prospective case series. Pain Pract. 2015, 15, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Chiang, M.C.; Tseng, M.T.; Pan, C.L.; Chao, C.C.; Hsieh, S.T. Progress in the treatment of small fiber peripheral neuropathy. Expert Rev. Neurother. 2015, 15, 305–313. [Google Scholar] [CrossRef]
- McCarthy, B.G.; Hsieh, S.T.; Stocks, A.; Hauer, P.; Macko, C.; Cornblath, D.R.; Griffin, J.W.; McArthur, J.C. Cutaneous innervation in sensory neuropathies: Evaluation by skin biopsy. Neurology 1995, 45, 1848–1855. [Google Scholar] [CrossRef]
- Herrmann, D.N.; Griffin, J.W.; Hauer, P.; Cornblath, D.R.; McArthur, J.C. Epidermal nerve fiber density and sural nerve morphometry in peripheral neuropathies. Neurology 1999, 53, 1634–1640. [Google Scholar] [CrossRef]
- Kim, S.H.; Chung, J.M. An experimental model for peripheral neuropathy produced by segmental spinal nerve ligation in the rat. Pain 1992, 50, 355–363. [Google Scholar] [CrossRef]
- Bennett, G.J.; Xie, Y.K. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain 1988, 33, 87–107. [Google Scholar] [CrossRef]
- Decosterd, I.; Woolf, C.J. Spared nerve injury: An animal model of persistent peripheral neuropathic pain. Pain 2000, 87, 149–158. [Google Scholar] [CrossRef]
- Challa, S.R. Surgical animal models of neuropathic pain: Pros and Cons. Int. J. Neurosci. 2015, 125, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Tseng, T.J.; Chen, C.C.; Hsieh, Y.L.; Hsieh, S.T. Effects of decompression on neuropathic pain behaviors and skin reinnervation in chronic constriction injury. Exp. Neurol. 2007, 204, 574–582. [Google Scholar] [CrossRef]
- Tseng, T.J.; Hsieh, Y.L.; Hsieh, S.T. Reversal of ERK activation in the dorsal horn after decompression in chronic constriction injury. Exp. Neurol. 2007, 206, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.F.; Hsieh, J.H.; Chiang, H.; Kan, H.W.; Huang, C.M.; Chellis, L.; Lin, B.S.; Miaw, S.C.; Pan, C.L.; Chao, C.C.; et al. Effective gene expression in the rat dorsal root ganglia with a non-viral vector delivered via spinal nerve injection. Sci. Rep. 2016, 6, 35612. [Google Scholar] [CrossRef] [PubMed]
- Hovaguimian, A.; Gibbons, C.H. Diagnosis and treatment of pain in small-fiber neuropathy. Curr. Pain Headache Rep. 2011, 15, 193–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cliffer, K.D.; Siuciak, J.A.; Carson, S.R.; Radley, H.E.; Park, J.S.; Lewis, D.R.; Zlotchenko, E.; Nguyen, T.; Garcia, K.; Tonra, J.R.; et al. Physiological characterization of Taxol-induced large-fiber sensory neuropathy in the rat. Ann. Neurol. 1998, 43, 46–55. [Google Scholar] [CrossRef]
- Davies, M.; Brophy, S.; Williams, R.; Taylor, A. The prevalence, severity, and impact of painful diabetic peripheral neuropathy in type 2 diabetes. Diabetes Care 2006, 29, 1518–1522. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, Y.L.; Chiang, H.; Tseng, T.J.; Hsieh, S.T. Enhancement of cutaneous nerve regeneration by 4-methylcatechol in resiniferatoxin-induced neuropathy. J. Neuropathol. Exp. Neurol. 2008, 67, 93–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.C.; Lu, S.C.; Hsieh, Y.L. Establishing a Mouse Model of a Pure Small Fiber Neuropathy with the Ultrapotent Agonist of Transient Receptor Potential Vanilloid Type 1. J. Vis. Exp. 2018. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, T.H.; Fu, Y.S.; Ho, W.Y.; Chen, T.H.; Hsieh, Y.L. Promotion of thermal analgesia and neuropeptidergic skin reinnervation by 4-methylcatechol in resiniferatoxin-induced neuropathy. Kaohsiung J. Med. Sci. 2013, 29, 405–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, P.S.; Schaumburg, H.H. Nervous system degeneration produced by acrylamide monomer. Environ. Health Perspect. 1975, 11, 129–133. [Google Scholar] [CrossRef]
- Suzuki, K.; Pfaff, L.D. Acrylamide neuropathy in rats. An electron microscopic study of degeneration and regeneration. Acta Neuropathol. 1973, 24, 197–213. [Google Scholar] [CrossRef] [PubMed]
- Ko, M.H.; Chen, W.P.; Hsieh, S.T. Neuropathology of skin denervation in acrylamide-induced neuropathy. Neurobiol. Dis. 2002, 11, 155–165. [Google Scholar] [CrossRef] [Green Version]
- Ling, B.; Authier, N.; Balayssac, D.; Eschalier, A.; Coudore, F. Assessment of nociception in acrylamide-induced neuropathy in rats. Pain 2005, 119, 104–112. [Google Scholar] [CrossRef]
- Yousef, M.I.; El-Demerdash, F.M. Acrylamide-induced oxidative stress and biochemical perturbations in rats. Toxicology 2006, 219, 133–141. [Google Scholar] [CrossRef]
- Yousefsani, B.S.; Akbarizadeh, N.; Pourahmad, J. The antioxidant and neuroprotective effects of Zolpidem on acrylamide-induced neurotoxicity using Wistar rat primary neuronal cortical culture. Toxicol. Rep. 2020, 7, 233–240. [Google Scholar] [CrossRef]
- Oda, S.S. Metformin Protects against Experimental Acrylamide Neuropathy in Rats. Drug Dev. Res. 2017, 78, 349–359. [Google Scholar] [CrossRef]
- Seretny, M.; Currie, G.L.; Sena, E.S.; Ramnarine, S.; Grant, R.; MacLeod, M.R.; Colvin, L.A.; Fallon, M. Incidence, prevalence, and predictors of chemotherapy-induced peripheral neuropathy: A systematic review and meta-analysis. Pain 2014, 155, 2461–2470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balayssac, D.; Ferrier, J.; Descoeur, J.; Ling, B.; Pezet, D.; Eschalier, A.; Authier, N. Chemotherapy-induced peripheral neuropathies: From clinical relevance to preclinical evidence. Expert Opin. Drug Saf. 2011, 10, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Yang, I.H.; Siddique, R.; Hosmane, S.; Thakor, N.; Hoke, A. Compartmentalized microfluidic culture platform to study mechanism of paclitaxel-induced axonal degeneration. Exp. Neurol. 2009, 218, 124–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravula, S.K.; Wang, M.S.; McClain, M.A.; Asress, S.A.; Frazier, B.; Glass, J.D. Spatiotemporal localization of injury potentials in DRG neurons during vincristine-induced axonal degeneration. Neurosci. Lett. 2007, 415, 34–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, E.S.; Windebank, A.J. Cisplatin-induced apoptosis of DRG neurons involves bax redistribution and cytochrome c release but not fas receptor signaling. Neurobiol. Dis. 2002, 9, 220–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, Y.; Li, Y.; Segal, R.A. A Mechanistic Understanding of Axon Degeneration in Chemotherapy-Induced Peripheral Neuropathy. Front. Neurosci. 2017, 11, 481. [Google Scholar] [CrossRef] [PubMed]
- Boehmerle, W.; Huehnchen, P.; Peruzzaro, S.; Balkaya, M.; Endres, M. Electrophysiological, behavioral and histological characterization of paclitaxel, cisplatin, vincristine and bortezomib-induced neuropathy in C57Bl/6 mice. Sci. Rep. 2014, 4, 6370. [Google Scholar] [CrossRef] [PubMed]
- Chiang, H.Y.; Chien, H.F.; Shen, H.H.; Yang, J.D.; Chen, Y.H.; Chen, J.H.; Hsieh, S.T. Reinnervation of muscular targets by nerve regeneration through guidance conduits. J. Neuropathol. Exp. Neurol. 2005, 64, 576–587. [Google Scholar] [CrossRef] [Green Version]
- Chine, V.B.; Au, N.P.B.; Kumar, G.; Ma, C.H.E. Targeting Axon Integrity to Prevent Chemotherapy-Induced Peripheral Neuropathy. Mol. Neurobiol. 2019, 56, 3244–3259. [Google Scholar] [CrossRef]
- Calvo, M.; Davies, A.J.; Hebert, H.L.; Weir, G.A.; Chesler, E.J.; Finnerup, N.B.; Levitt, R.C.; Smith, B.H.; Neely, G.G.; Costigan, M.; et al. The Genetics of Neuropathic Pain from Model Organisms to Clinical Application. Neuron 2019, 104, 637–653. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.W.; Yue, K.; Ru, Q.; Xiong, Q.; Ma, B.M.; Tian, X.; Li, C.Y. Differential expression of ATP-gated P2X receptors in DRG between chronic neuropathic pain and visceralgia rat models. Purinergic Signal. 2016, 12, 79–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novakovic, S.D.; Kassotakis, L.C.; Oglesby, I.B.; Smith, J.A.; Eglen, R.M.; Ford, A.P.; Hunter, J.C. Immunocytochemical localization of P2X3 purinoceptors in sensory neurons in naive rats and following neuropathic injury. Pain 1999, 80, 273–282. [Google Scholar] [CrossRef]
- Leng, C.; Chen, L.; Li, C. Alteration of P2X1-6 receptor expression in retrograde Fluorogold-labeled DRG neurons from rat chronic neuropathic pain model. Biomed. Rep. 2019, 10, 225–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barclay, J.; Patel, S.; Dorn, G.; Wotherspoon, G.; Moffatt, S.; Eunson, L.; Abdel’al, S.; Natt, F.; Hall, J.; Winter, J.; et al. Functional downregulation of P2X3 receptor subunit in rat sensory neurons reveals a significant role in chronic neuropathic and inflammatory pain. J. Neurosci. 2002, 22, 8139–8147. [Google Scholar] [CrossRef] [Green Version]
- Kage, K.; Niforatos, W.; Zhu, C.Z.; Lynch, K.J.; Honore, P.; Jarvis, M.F. Alteration of dorsal root ganglion P2X3 receptor expression and function following spinal nerve ligation in the rat. Exp. Brain Res. 2002, 147, 511–519. [Google Scholar] [CrossRef]
- Fukuoka, T.; Tokunaga, A.; Tachibana, T.; Dai, Y.; Yamanaka, H.; Noguchi, K. VR1, but not P2X(3), increases in the spared L4 DRG in rats with L5 spinal nerve ligation. Pain 2002, 99, 111–120. [Google Scholar] [CrossRef]
- Kim, C.; Chung, J.M.; Chung, K. Changes in the gene expression of six subtypes of P2X receptors in rat dorsal root ganglion after spinal nerve ligation. Neurosci. Lett. 2003, 337, 81–84. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Guo, Q.L.; Zou, W.Y.; Huang, C.S.; Yan, J.Q. Cyclooxygenase inhibitors suppress the expression of P2X(3) receptors in the DRG and attenuate hyperalgesia following chronic constriction injury in rats. Neurosci. Lett. 2010, 478, 77–81. [Google Scholar] [CrossRef]
- Hsieh, Y.L.; Chiang, H.; Lue, J.H.; Hsieh, S.T. P2X3-mediated peripheral sensitization of neuropathic pain in resiniferatoxin-induced neuropathy. Exp. Neurol. 2012, 235, 316–325. [Google Scholar] [CrossRef]
- Hori, K.; Ozaki, N.; Suzuki, S.; Sugiura, Y. Upregulations of P2X(3) and ASIC3 involve in hyperalgesia induced by cisplatin administration in rats. Pain 2010, 149, 393–405. [Google Scholar] [CrossRef]
- Watanabe, M.; Ueda, T.; Shibata, Y.; Kumamoto, N.; Ugawa, S. The role of TRPV1 channels in carrageenan-induced mechanical hyperalgesia in mice. Neuroreport 2015, 26, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J.; Leffler, A.; Malmberg, A.B.; Martin, W.J.; Trafton, J.; Petersen-Zeitz, K.R.; Koltzenburg, M.; Basbaum, A.I.; Julius, D. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 2000, 288, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Story, G.M.; Peier, A.M.; Reeve, A.J.; Eid, S.R.; Mosbacher, J.; Hricik, T.R.; Earley, T.J.; Hergarden, A.C.; Andersson, D.A.; Hwang, S.W.; et al. ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell 2003, 112, 819–829. [Google Scholar] [CrossRef] [Green Version]
- Su, L.; Shu, R.; Song, C.; Yu, Y.; Wang, G.; Li, Y.; Liu, C. Downregulations of TRPM8 expression and membrane trafficking in dorsal root ganglion mediate the attenuation of cold hyperalgesia in CCI rats induced by GFRalpha3 knockdown. Brain Res. Bull. 2017, 135, 8–24. [Google Scholar] [CrossRef]
- Dib-Hajj, S.D.; Cummins, T.R.; Black, J.A.; Waxman, S.G. Sodium channels in normal and pathological pain. Annu. Rev. Neurosci. 2010, 33, 325–347. [Google Scholar] [CrossRef] [Green Version]
- Su, L.; Wang, C.; Yu, Y.H.; Ren, Y.Y.; Xie, K.L.; Wang, G.L. Role of TRPM8 in dorsal root ganglion in nerve injury-induced chronic pain. BMC Neurosci. 2011, 12, 120. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.H.; Lin, J.P.; Huang, L.E.; Yang, Y.; Chen, C.Q.; Li, N.N.; Su, M.Y.; Zhao, X.; Zhu, S.M.; Yao, Y.X. Silencing of spinal Trpv1 attenuates neuropathic pain in rats by inhibiting CAMKII expression and ERK2 phosphorylation. Sci. Rep. 2019, 9, 2769. [Google Scholar] [CrossRef] [Green Version]
- Black, J.A.; Dib-Hajj, S.; McNabola, K.; Jeste, S.; Rizzo, M.A.; Kocsis, J.D.; Waxman, S.G. Spinal sensory neurons express multiple sodium channel alpha-subunit mRNAs. Brain Res. Mol. Brain Res. 1996, 43, 117–131. [Google Scholar] [CrossRef]
- Dib-Hajj, S.D.; Tyrrell, L.; Black, J.A.; Waxman, S.G. NaN, a novel voltage-gated Na channel, is expressed preferentially in peripheral sensory neurons and down-regulated after axotomy. Proc. Natl. Acad. Sci. USA 1998, 95, 8963–8968. [Google Scholar] [CrossRef] [Green Version]
- Waxman, S.G. Transcriptional channelopathies: An emerging class of disorders. Nat. Rev. Neurosci. 2001, 2, 652–659. [Google Scholar] [CrossRef]
- Chattopadhyay, M.; Mata, M.; Fink, D.J. Continuous delta-opioid receptor activation reduces neuronal voltage-gated sodium channel (NaV1.7) levels through activation of protein kinase C in painful diabetic neuropathy. J. Neurosci. 2008, 28, 6652–6658. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Morrow, T.J.; Paulson, P.E.; Isom, L.L.; Wiley, J.W. Early painful diabetic neuropathy is associated with differential changes in tetrodotoxin-sensitive and -resistant sodium channels in dorsal root ganglion neurons in the rat. J. Biol. Chem. 2004, 279, 29341–29350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laedermann, C.J.; Cachemaille, M.; Kirschmann, G.; Pertin, M.; Gosselin, R.D.; Chang, I.; Albesa, M.; Towne, C.; Schneider, B.L.; Kellenberger, S.; et al. Dysregulation of voltage-gated sodium channels by ubiquitin ligase NEDD4-2 in neuropathic pain. J. Clin. Investig. 2013, 123, 3002–3013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catterall, W.A.; Perez-Reyes, E.; Snutch, T.P.; Striessnig, J. International Union of Pharmacology. XLVIII. Nomenclature and structure-function relationships of voltage-gated calcium channels. Pharmacol. Rev. 2005, 57, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Iftinca, M.C. Neuronal T-type calcium channels: What’s new? Iftinca: T-type channel regulation. J. Med. Life 2011, 4, 126–138. [Google Scholar] [PubMed]
- Dolphin, A.C. Voltage-gated calcium channel alpha 2delta subunits: An assessment of proposed novel roles. F1000Res 2018, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talley, E.M.; Cribbs, L.L.; Lee, J.H.; Daud, A.; Perez-Reyes, E.; Bayliss, D.A. Differential distribution of three members of a gene family encoding low voltage-activated (T-type) calcium channels. J. Neurosci. 1999, 19, 1895–1911. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.B.; Martinez-Salgado, C.; Heppenstall, P.A.; Lewin, G.R. A T-type calcium channel required for normal function of a mammalian mechanoreceptor. Nat. Neurosci. 2003, 6, 724–730. [Google Scholar] [CrossRef]
- Rose, K.E.; Lunardi, N.; Boscolo, A.; Dong, X.; Erisir, A.; Jevtovic-Todorovic, V.; Todorovic, S.M. Immunohistological demonstration of CaV3.2 T-type voltage-gated calcium channel expression in soma of dorsal root ganglion neurons and peripheral axons of rat and mouse. Neuroscience 2013, 250, 263–274. [Google Scholar] [CrossRef] [Green Version]
- Kang, X.J.; Chi, Y.N.; Chen, W.; Liu, F.Y.; Cui, S.; Liao, F.F.; Cai, J.; Wan, Y. Increased expression of CaV3.2 T-type calcium channels in damaged DRG neurons contributes to neuropathic pain in rats with spared nerve injury. Mol. Pain 2018, 14. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.D.; Chaplan, S.R.; Higuera, E.S.; Sorkin, L.S.; Stauderman, K.A.; Williams, M.E.; Yaksh, T.L. Upregulation of dorsal root ganglion (alpha)2(delta) calcium channel subunit and its correlation with allodynia in spinal nerve-injured rats. J. Neurosci. 2001, 21, 1868–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.S.; Yoon, C.H.; Lee, S.J.; Park, S.Y.; Yoo, H.J.; Cho, H.J. Changes in voltage-gated calcium channel alpha(1) gene expression in rat dorsal root ganglia following peripheral nerve injury. Brain Res. Mol. Brain Res. 2001, 96, 151–156. [Google Scholar] [CrossRef]
- Lotem, J.; Sachs, L. Epigenetics and the plasticity of differentiation in normal and cancer stem cells. Oncogene 2006, 25, 7663–7672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atlasi, Y.; Stunnenberg, H.G. The interplay of epigenetic marks during stem cell differentiation and development. Nat. Rev. Genet. 2017, 18, 643–658. [Google Scholar] [CrossRef]
- Penas, C.; Navarro, X. Epigenetic Modifications Associated to Neuroinflammation and Neuropathic Pain After Neural Trauma. Front. Cell. Neurosci. 2018, 12, 158. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.Y.; Penninger, J.M. Transcriptional mechanisms underlying neuropathic pain: DREAM, transcription factors and future pain management? Expert Rev. Neurother. 2002, 2, 677–689. [Google Scholar] [CrossRef]
- Moss, A.; Blackburn-Munro, G.; Garry, E.M.; Blakemore, J.A.; Dickinson, T.; Rosie, R.; Mitchell, R.; Fleetwood-Walker, S.M. A role of the ubiquitin-proteasome system in neuropathic pain. J. Neurosci. 2002, 22, 1363–1372. [Google Scholar] [CrossRef] [Green Version]
- Vogenberg, F.R.; Isaacson Barash, C.; Pursel, M. Personalized medicine: Part 1: Evolution and development into theranostics. Pharm. Ther. 2010, 35, 560–576. [Google Scholar]
- Khorasanizadeh, S. The nucleosome: From genomic organization to genomic regulation. Cell 2004, 116, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Jenuwein, T.; Allis, C.D. Translating the histone code. Science 2001, 293, 1074–1080. [Google Scholar] [CrossRef] [Green Version]
- Shao, C.; Gao, Y.; Jin, D.; Xu, X.; Tan, S.; Yu, H.; Zhao, Q.; Zhao, L.; Wang, W.; Wang, D. DNMT3a methylation in neuropathic pain. J. Pain Res. 2017, 10, 2253–2262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garriga, J.; Laumet, G.; Chen, S.R.; Zhang, Y.; Madzo, J.; Issa, J.J.; Pan, H.L.; Jelinek, J. Nerve Injury-Induced Chronic Pain Is Associated with Persistent DNA Methylation Reprogramming in Dorsal Root Ganglion. J. Neurosci. 2018, 38, 6090–6101. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.H.; Hu, J.; Zhou, Y.L.; Qin, X.; Song, Z.Y.; Yang, P.P.; Hu, S.; Jiang, X.; Xu, G.Y. Promoted Interaction of Nuclear Factor-kappaB With Demethylated Purinergic P2X3 Receptor Gene Contributes to Neuropathic Pain in Rats With Diabetes. Diabetes 2015, 64, 4272–4284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.Y.; Liang, L.; Gu, X.; Li, Z.; Wu, S.; Sun, L.; Atianjoh, F.E.; Feng, J.; Mo, K.; Jia, S.; et al. DNA methyltransferase DNMT3a contributes to neuropathic pain by repressing Kcna2 in primary afferent neurons. Nat. Commun. 2017, 8, 14712. [Google Scholar] [CrossRef] [PubMed]
- Chuang, D.M.; Leng, Y.; Marinova, Z.; Kim, H.J.; Chiu, C.T. Multiple roles of HDAC inhibition in neurodegenerative conditions. Trends Neurosci. 2009, 32, 591–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, J.S.; Haggarty, S.J.; Giacometti, E.; Dannenberg, J.H.; Joseph, N.; Gao, J.; Nieland, T.J.; Zhou, Y.; Wang, X.; Mazitschek, R.; et al. HDAC2 negatively regulates memory formation and synaptic plasticity. Nature 2009, 459, 55–60. [Google Scholar] [CrossRef]
- Sando, R., III; Gounko, N.; Pieraut, S.; Liao, L.; Yates, J., III; Maximov, A. HDAC4 governs a transcriptional program essential for synaptic plasticity and memory. Cell 2012, 151, 821–834. [Google Scholar] [CrossRef] [Green Version]
- Gu, P.; Pan, Z.; Wang, X.M.; Sun, L.; Tai, L.W.; Cheung, C.W. Histone deacetylase 5 (HDAC5) regulates neuropathic pain through SRY-related HMG-box 10 (SOX10)-dependent mechanism in mice. Pain 2018, 159, 526–539. [Google Scholar] [CrossRef]
- Matsushita, Y.; Araki, K.; Omotuyi, O.; Mukae, T.; Ueda, H. HDAC inhibitors restore C-fibre sensitivity in experimental neuropathic pain model. Br. J. Pharmacol. 2013, 170, 991–998. [Google Scholar] [CrossRef] [Green Version]
- Denk, F.; Huang, W.; Sidders, B.; Bithell, A.; Crow, M.; Grist, J.; Sharma, S.; Ziemek, D.; Rice, A.S.; Buckley, N.J.; et al. HDAC inhibitors attenuate the development of hypersensitivity in models of neuropathic pain. Pain 2013, 154, 1668–1679. [Google Scholar] [CrossRef] [Green Version]
- Ji, R.R.; Baba, H.; Brenner, G.J.; Woolf, C.J. Nociceptive-specific activation of ERK in spinal neurons contributes to pain hypersensitivity. Nat. Neurosci. 1999, 2, 1114–1119. [Google Scholar] [CrossRef] [PubMed]
- Samour, M.S.; Shaikh, S.M.; Mahns, D.A.; Shortland, P.J. Noxious, but not innocuous, thermal stimuli evoke pERK expression in dorsal horn neurons after spared nerve injury in adult rats. Neurosci. Lett. 2017, 654, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.J.; Jia, L.; Zhang, X.; Wei, H.; Yue, S.W. MAPK Pathways Are Involved in Neuropathic Pain in Rats with Chronic Compression of the Dorsal Root Ganglion. Evid. Based Complement. Alternat. Med. 2016, 2016, 6153215. [Google Scholar] [CrossRef] [PubMed]
- Chuderland, D.; Konson, A.; Seger, R. Identification and characterization of a general nuclear translocation signal in signaling proteins. Mol. Cell 2008, 31, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Impey, S.; Obrietan, K.; Wong, S.T.; Poser, S.; Yano, S.; Wayman, G.; Deloulme, J.C.; Chan, G.; Storm, D.R. Cross talk between ERK and PKA is required for Ca2+ stimulation of CREB-dependent transcription and ERK nuclear translocation. Neuron 1998, 21, 869–883. [Google Scholar] [CrossRef] [Green Version]
- Monje, P.; Hernandez-Losa, J.; Lyons, R.J.; Castellone, M.D.; Gutkind, J.S. Regulation of the transcriptional activity of c-Fos by ERK. A novel role for the prolyl isomerase PIN1. J. Biol. Chem. 2005, 280, 35081–35084. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.J.; Ji, R.R. c-Fos and pERK, which is a better marker for neuronal activation and central sensitization after noxious stimulation and tissue injury? Open Pain J. 2009, 2, 11–17. [Google Scholar] [CrossRef]
- Seijffers, R.; Mills, C.D.; Woolf, C.J. ATF3 increases the intrinsic growth state of DRG neurons to enhance peripheral nerve regeneration. J. Neurosci. 2007, 27, 7911–7920. [Google Scholar] [CrossRef] [Green Version]
- Tsujino, H.; Kondo, E.; Fukuoka, T.; Dai, Y.; Tokunaga, A.; Miki, K.; Yonenobu, K.; Ochi, T.; Noguchi, K. Activating transcription factor 3 (ATF3) induction by axotomy in sensory and motoneurons: A novel neuronal marker of nerve injury. Mol. Cell. Neurosci. 2000, 15, 170–182. [Google Scholar] [CrossRef]
- Ding, S.; Yu, Q.; Wang, J.; Zhu, L.; Li, T.; Guo, X.; Zhang, X. Activation of ATF3/AP-1 signaling pathway is required for P2X3-induced endometriosis pain. Hum. Reprod. 2020, 35, 1130–1144. [Google Scholar] [CrossRef]
- Staub, O.; Dho, S.; Henry, P.; Correa, J.; Ishikawa, T.; McGlade, J.; Rotin, D. WW domains of Nedd4 bind to the proline-rich PY motifs in the epithelial Na+ channel deleted in Liddle’s syndrome. EMBO J. 1996, 15, 2371–2380. [Google Scholar] [CrossRef] [PubMed]
- Younger, J.M.; Ren, H.Y.; Chen, L.; Fan, C.Y.; Fields, A.; Patterson, C.; Cyr, D.M. A foldable CFTR{Delta}F508 biogenic intermediate accumulates upon inhibition of the Hsc70-CHIP E3 ubiquitin ligase. J. Cell Biol. 2004, 167, 1075–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altier, C.; Garcia-Caballero, A.; Simms, B.; You, H.; Chen, L.; Walcher, J.; Tedford, H.W.; Hermosilla, T.; Zamponi, G.W. The Cavbeta subunit prevents RFP2-mediated ubiquitination and proteasomal degradation of L-type channels. Nat. Neurosci. 2011, 14, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Caballero, A.; Gadotti, V.M.; Stemkowski, P.; Weiss, N.; Souza, I.A.; Hodgkinson, V.; Bladen, C.; Chen, L.; Hamid, J.; Pizzoccaro, A.; et al. The deubiquitinating enzyme USP5 modulates neuropathic and inflammatory pain by enhancing Cav3.2 channel activity. Neuron 2014, 83, 1144–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nascimento, A.I.; Mar, F.M.; Sousa, M.M. The intriguing nature of dorsal root ganglion neurons: Linking structure with polarity and function. Prog. Neurobiol. 2018, 168, 86–103. [Google Scholar] [CrossRef] [PubMed]
- Haberberger, R.V.; Barry, C.; Dominguez, N.; Matusica, D. Human Dorsal Root Ganglia. Front. Cell. Neurosci. 2019, 13, 271. [Google Scholar] [CrossRef] [PubMed]
- Puljak, L.; Kojundzic, S.L.; Hogan, Q.H.; Sapunar, D. Lidocaine injection into the rat dorsal root ganglion causes neuroinflammation. Anesth. Analg. 2009, 108, 1021–1026. [Google Scholar] [CrossRef] [Green Version]
- Fischer, G.; Kostic, S.; Nakai, H.; Park, F.; Sapunar, D.; Yu, H.; Hogan, Q. Direct injection into the dorsal root ganglion: Technical, behavioral, and histological observations. J. Neurosci. Methods 2011, 199, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.J.; Walla, B.C.; Diaz, M.F.; Fuller, G.N.; Gutstein, H.B. Intermittent lumbar puncture in rats: A novel method for the experimental study of opioid tolerance. Anesth. Analg. 2006, 103, 714–720. [Google Scholar] [CrossRef]
- Haggerty, D.L.; Grecco, G.G.; Reeves, K.C.; Atwood, B. Adeno-Associated Viral Vectors in Neuroscience Research. Mol. Ther. Methods Clin. Dev. 2020, 17, 69–82. [Google Scholar] [CrossRef]
- Daya, S.; Berns, K.I. Gene therapy using adeno-associated virus vectors. Clin. Microbiol. Rev. 2008, 21, 583–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, M.R.; Ehlert, E.M.; Eggers, R.; Pool, C.W.; Hermening, S.; Huseinovic, A.; Timmermans, E.; Blits, B.; Verhaagen, J. Comparison of AAV serotypes for gene delivery to dorsal root ganglion neurons. Mol. Ther. 2010, 18, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Haery, L.; Deverman, B.E.; Matho, K.S.; Cetin, A.; Woodard, K.; Cepko, C.; Guerin, K.I.; Rego, M.A.; Ersing, I.; Bachle, S.M.; et al. Adeno-Associated Virus Technologies and Methods for Targeted Neuronal Manipulation. Front. Neuroanat. 2019, 13, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mingozzi, F.; High, K.A. Immune responses to AAV vectors: Overcoming barriers to successful gene therapy. Blood 2013, 122, 23–36. [Google Scholar] [CrossRef]
- Russell, S.; Bennett, J.; Wellman, J.A.; Chung, D.C.; Yu, Z.F.; Tillman, A.; Wittes, J.; Pappas, J.; Elci, O.; McCague, S.; et al. Efficacy and safety of voretigene neparvovec (AAV2-hRPE65v2) in patients with RPE65-mediated inherited retinal dystrophy: A randomised, controlled, open-label, phase 3 trial. Lancet 2017, 390, 849–860. [Google Scholar] [CrossRef]
- Kushnarev, M.; Pirvulescu, I.P.; Candido, K.D.; Knezevic, N.N. Neuropathic pain: Preclinical and early clinical progress with voltage-gated sodium channel blockers. Expert Opin. Investig. Drugs 2020, 29, 259–271. [Google Scholar] [CrossRef]
- Urru, M.; Muzzi, M.; Coppi, E.; Ranieri, G.; Buonvicino, D.; Camaioni, E.; Coppini, R.; Pugliese, A.M.; Tanaka, B.; Estacion, M.; et al. Dexpramipexole blocks Nav1.8 sodium channels and provides analgesia in multiple nociceptive and neuropathic pain models. Pain 2020, 161, 831–841. [Google Scholar] [CrossRef]
- Cudkowicz, M.; Bozik, M.E.; Ingersoll, E.W.; Miller, R.; Mitsumoto, H.; Shefner, J.; Moore, D.H.; Schoenfeld, D.; Mather, J.L.; Archibald, D.; et al. The effects of dexpramipexole (KNS-760704) in individuals with amyotrophic lateral sclerosis. Nat. Med. 2011, 17, 1652–1656. [Google Scholar] [CrossRef]
- Cudkowicz, M.E.; van den Berg, L.H.; Shefner, J.M.; Mitsumoto, H.; Mora, J.S.; Ludolph, A.; Hardiman, O.; Bozik, M.E.; Ingersoll, E.W.; Archibald, D.; et al. Dexpramipexole versus placebo for patients with amyotrophic lateral sclerosis (EMPOWER): A randomised, double-blind, phase 3 trial. Lancet Neurol. 2013, 12, 1059–1067. [Google Scholar] [CrossRef]
- Dworetzky, S.I.; Hebrank, G.T.; Archibald, D.G.; Reynolds, I.J.; Farwell, W.; Bozik, M.E. The targeted eosinophil-lowering effects of dexpramipexole in clinical studies. Blood Cells Mol. Dis. 2017, 63, 62–65. [Google Scholar] [CrossRef]
- Ji, G.; Zhou, S.; Carlton, S.M. Intact Adelta-fibers up-regulate transient receptor potential A1 and contribute to cold hypersensitivity in neuropathic rats. Neuroscience 2008, 154, 1054–1066. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Zhang, S.J.; Yang, L.; Fang, X.L.; Hu, H.F.; Zhao, M.Y.; Li, L.; Guo, Y.Y.; Shao, J.P. Voltage-gated sodium channel 1.7 expression decreases in dorsal root ganglia in a spinal nerve ligation neuropathic pain model. Kaohsiung J. Med. Sci. 2019, 35, 493–500. [Google Scholar] [CrossRef]
- Ma, R.S.Y.; Kayani, K.; Whyte-Oshodi, D.; Whyte-Oshodi, A.; Nachiappan, N.; Gnanarajah, S.; Mohammed, R. Voltage gated sodium channels as therapeutic targets for chronic pain. J. Pain Res. 2019, 12, 2709–2722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Leng, C.; Ru, Q.; Xiong, Q.; Zhou, M.; Wu, Y. Retrograde Labeling of Different Distribution Features of DRG P2X2 and P2X3 Receptors in a Neuropathic Pain Rat Model. Biomed. Res. Int. 2020, 2020, 9861459. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, G.W.; Wang, C.; Gu, Y.; Huang, L.Y. Mechanisms underlying enhanced P2X receptor-mediated responses in the neuropathic pain state. Pain 2005, 119, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Frederick, J.; Buck, M.E.; Matson, D.J.; Cortright, D.N. Increased TRPA1, TRPM8, and TRPV2 expression in dorsal root ganglia by nerve injury. Biochem. Biophys. Res. Commun. 2007, 358, 1058–1064. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ling, D.; Wu, C.; Han, J.; Zhao, Y. Baicalin prevents the up-regulation of TRPV1 in dorsal root ganglion and attenuates chronic neuropathic pain. Vet. Med. Sci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Staaf, S.; Oerther, S.; Lucas, G.; Mattsson, J.P.; Ernfors, P. Differential regulation of TRP channels in a rat model of neuropathic pain. Pain 2009, 144, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Casals-Diaz, L.; Casas, C.; Navarro, X. Changes of voltage-gated sodium channels in sensory nerve regeneration and neuropathic pain models. Restor. Neurol. Neurosci. 2015, 33, 321–334. [Google Scholar] [CrossRef]
- Laedermann, C.J.; Pertin, M.; Suter, M.R.; Decosterd, I. Voltage-gated sodium channel expression in mouse DRG after SNI leads to re-evaluation of projections of injured fibers. Mol. Pain 2014, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Kan, H.W.; Chang, C.H.; Lin, C.L.; Lee, Y.C.; Hsieh, S.T.; Hsieh, Y.L. Downregulation of adenosine and adenosine A1 receptor contributes to neuropathic pain in resiniferatoxin neuropathy. Pain 2018, 159, 1580–1591. [Google Scholar] [CrossRef] [PubMed]
- Ta, L.E.; Bieber, A.J.; Carlton, S.M.; Loprinzi, C.L.; Low, P.A.; Windebank, A.J. Transient Receptor Potential Vanilloid 1 is essential for cisplatin-induced heat hyperalgesia in mice. Mol. Pain 2010, 6, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leo, M.; Schmitt, L.I.; Jastrow, H.; Thomale, J.; Kleinschnitz, C.; Hagenacker, T. Cisplatin alters the function and expression of N-type voltage-gated calcium channels in the absence of morphological damage of sensory neurons. Mol. Pain 2017, 13. [Google Scholar] [CrossRef] [PubMed]

| Advantage | Disadvantage | Application | |
|---|---|---|---|
| SNL [21] | Specific injury in L5 and L6 DRG | Time consuming | Spinal nerve injection |
| CCI [22] | Combine injury and uninjured fibers | Variation in each operation | Decompression model |
| SNI [23] | Combine injury and uninjured fibers Robustness of the response | Low local inflammation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeh, T.-Y.; Luo, I.-W.; Hsieh, Y.-L.; Tseng, T.-J.; Chiang, H.; Hsieh, S.-T. Peripheral Neuropathic Pain: From Experimental Models to Potential Therapeutic Targets in Dorsal Root Ganglion Neurons. Cells 2020, 9, 2725. https://doi.org/10.3390/cells9122725
Yeh T-Y, Luo I-W, Hsieh Y-L, Tseng T-J, Chiang H, Hsieh S-T. Peripheral Neuropathic Pain: From Experimental Models to Potential Therapeutic Targets in Dorsal Root Ganglion Neurons. Cells. 2020; 9(12):2725. https://doi.org/10.3390/cells9122725
Chicago/Turabian StyleYeh, Ti-Yen, I-Wei Luo, Yu-Lin Hsieh, To-Jung Tseng, Hao Chiang, and Sung-Tsang Hsieh. 2020. "Peripheral Neuropathic Pain: From Experimental Models to Potential Therapeutic Targets in Dorsal Root Ganglion Neurons" Cells 9, no. 12: 2725. https://doi.org/10.3390/cells9122725