HLA Class II Genotype Does Not Affect the Myelin Responsiveness of Multiple Sclerosis Patients

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Ethics
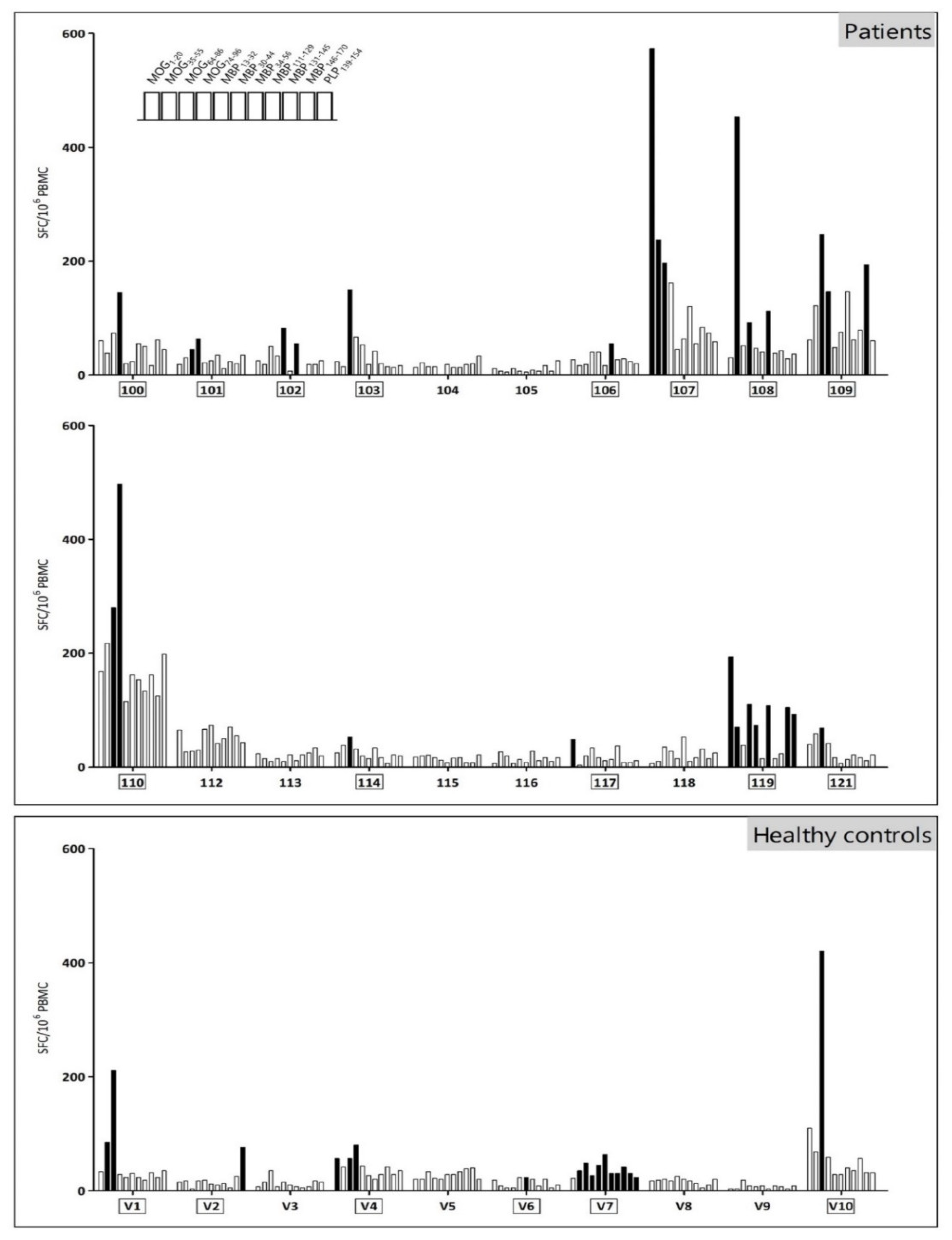
2.2. Myelin Reactivity Screening
2.3. HLA Class II Haplotyping
2.4. Data Analysis
3. Results
3.1. Study Subjects’ Characteristics
3.2. The Myelin Reactivity Profile of MS Patients and Healthy Controls Displays a High Inter- and Intramolecular Variability
3.3. The Majority of MS Patients Carries MS-Associated HLA Risk Alleles
3.4. The HLA Class II Genotype Does Not Affect Myelin-Specific Responsiveness
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hohlfeld, R.; Dornmair, K.; Meinl, E.; Wekerle, H. The search for the target antigens of multiple sclerosis, part 1: Autoreactive CD4+ T lymphocytes as pathogenic effectors and therapeutic targets. Lancet Neurol. 2015, 15, 198–209. [Google Scholar] [CrossRef]
- Sáez-Torres, I.; Brieva, L.; Espejo, C.; Barrau, M.A.; Montalban, X.; Martínez-Caceres, E.M. Specific proliferation towards myelin antigens in patients with multiple sclerosis during a relapse. Autoimmunity 2002, 35, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Greer, J.M. Autoimmune T-cell reactivity to myelin proteolipids and glycolipids in multiple sclerosis. Mult. Scler. Int. 2013, 2013, 151427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goebels, N.; Hofstetter, H.; Schmidt, S.; Brunner, C.; Wekerle, H.; Hohlfeld, R. Repertoire dynamics of autoreactive T cells in multiple sclerosis patients and healthy subjects: Epitope spreading versus clonal persistence. Brain 2000, 123, 508–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuohy, V.K.; Kinkel, R.P. Epitope spreading: A mechanism for progression of autoimmune disease. Arch. Immunol. Ther. Exp. 2000, 48, 347–351. [Google Scholar]
- Tuohy, V.K.; Yu, M.; Yin, L.; Kawczak, J.A.; Johnson, J.M.; Mathisen, P.M.; Weinstock-Guttman, B.; Kinkel, R.P. The epitope spreading cascade during progression of experimental autoimmune encephalomyelitis and multiple sclerosis. Immunol. Rev. 1998, 164, 93–100. [Google Scholar] [CrossRef]
- Davies, S.; Nicholson, T.; Laura, M.; Giovannoni, G.; Altmann, D.M. Spread of T lymphocyte immune responses to myelin epitopes with duration of multiple sclerosis. J. Neuropathol. Exp. Neurol. 2005, 64, 371–377. [Google Scholar] [CrossRef] [Green Version]
- Hilkens, C.M.; Isaacs, J.D.; Thomson, A.W. Development of dendritic cell-based immunotherapy for autoimmunity. Int. Rev. Immunol. 2010, 29, 156–183. [Google Scholar] [CrossRef]
- Yu, M.; Johnson, J.M.; Tuohy, V.K. A predictable sequential determinant spreading cascade invariably accompanies progression of experimental autoimmune encephalomyelitis: A basis for peptide-specific therapy after onset of clinical disease. J. Exp. Med. 1996, 183, 1777–1788. [Google Scholar] [CrossRef] [Green Version]
- Derdelinckx, J.; Mansilla, M.J.; De Laere, M.; Lee, W.P.; Navarro-Barriuso, J.; Wens, I.; Nkansah, I.; Daans, J.; De Reu, H.; Jolanta Keliris, A.; et al. Clinical and immunological control of experimental autoimmune encephalomyelitis by tolerogenic dendritic cells loaded with MOG-encoding mRNA. J. Neuroinflamm. 2019, 16, 167. [Google Scholar] [CrossRef] [Green Version]
- Corthay, A. A three-cell model for activation of naive T helper cells. Scand. J. Immunol. 2006, 64, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, J.M.; Lalor, S.J.; Sweeney, C.M.; Tubridy, N.; Mills, K.H. T cells in multiple sclerosis and experimental autoimmune encephalomyelitis. Clin. Exp. Immunol. 2010, 162, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hollenbach, J.A.; Oksenberg, J.R. The immunogenetics of multiple sclerosis: A comprehensive review. J. Autoimmun. 2015, 64, 13–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawcer, S.; Hellenthal, G.; Pirinen, M.; Spencer, C.C.; Patsopoulos, N.A.; Moutsianas, L.; Dilthey, A.; Su, Z.; Freeman, C.; Hunt, S.E.; et al. Genetic risk and a primary role for cell-mediated immune mechanisms in multiple sclerosis. Nature 2011, 476, 214–219. [Google Scholar] [PubMed]
- Schmidt, H.; Williamson, D.; Ashley-Koch, A. HLA-DR15 haplotype and multiple sclerosis: A HuGE review. Am. J. Epidemiol. 2007, 165, 1097–1109. [Google Scholar] [CrossRef]
- Moutsianas, L.; Jostins, L.; Beecham, A.H.; Dilthey, A.T.; Xifara, D.K.; Ban, M.; Shah, T.S.; Patsopoulos, N.A.; Alfredsson, L.; Anderson, C.A.; et al. Class II HLA interactions modulate genetic risk for multiple sclerosis. Nat. Genet. 2015, 47, 1107–1113. [Google Scholar]
- Field, J.; Browning, S.R.; Johnson, L.J.; Danoy, P.; Varney, M.D.; Tait, B.D.; Gandhi, K.S.; Charlesworth, J.C.; Heard, R.N.; Australia and New Zealand Multiple Sclerosis Genetics Consortium; et al. A polymorphism in the HLA-DPB1 gene is associated with susceptibility to multiple sclerosis. PLoS ONE 2010, 5, e13454. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, D.; Arrhenius, T.; Sidney, J.; Del Guercio, M.F.; Albertson, M.; Wall, M.; Oseroff, C.; Southwood, S.; Colón, S.M.; Gaeta, F.C.; et al. On the interaction of promiscuous antigenic peptides with different DR alleles. Identification of common structural motifs. J. Immunol. 1991, 147, 2663–2669. [Google Scholar]
- Wucherpfennig, K.W.; Strominger, J.L. Selective binding of self peptides to disease-associated major histocompatibility complex (MHC) molecules: A mechanism for MHC-linked susceptibility to human autoimmune diseases. J. Exp. Med. 1995, 181, 1597–1601. [Google Scholar] [CrossRef]
- Busch, R.; Strang, G.; Howland, K.; Rothbard, J.B. Degenerate binding of immunogenic peptides to HLA-DR proteins on B cell surfaces. Int. Immunol. 1990, 2, 443–451. [Google Scholar] [CrossRef]
- Valli, A.; Sette, A.; Kappos, L.; Oseroff, C.; Sidney, J.; Miescher, G.; Hochberger, M.; Albert, E.D.; Adorini, L. Binding of myelin basic protein peptides to human histocompatibility leukocyte antigen class II molecules and their recognition by T cells from multiple sclerosis patients. J. Clin. Investig. 1993, 91, 616–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fridkis-Hareli, M.; Teitelbaum, D.; Gurevich, E.; Pecht, I.; Brautbar, C.; Kwon, O.J.; Brenner, T.; Arnon, R.; Sela, M. Direct binding of myelin basic protein and synthetic copolymer 1 to class II major histocompatibility complex molecules on living antigen-presenting cells—Specificity and promiscuity. Proc. Natl. Acad. Sci. USA 1994, 91, 4872–4876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissert, R.; Kuhle, J.; de Graaf, K.L.; Wienhold, W.; Herrmann, M.M.; Müller, C.; Forsthuber, T.G.; Wiesmüller, K.H.; Melms, A. High immunogenicity of intracellular myelin oligodendrocyte glycoprotein epitopes. J. Immunol. 2002, 169, 548–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polman, C.H.; Reingold, S.C.; Banwell, B.; Clanet, M.; Cohen, J.A.; Filippi, M.; Fujihara, K.; Havrdova, E.; Hutchinson, M.; Kappos, L.; et al. Diagnostic criteria for multiple sclerosis: 2010 revisions to the McDonald criteria. Ann. Neurol. 2011, 69, 292–302. [Google Scholar] [CrossRef] [Green Version]
- Vergelli, M.; Kalbus, M.; Rojo, S.C.; Hemmer, B.; Kalbacher, H.; Tranquill, L.; Beck, H.; McFarland, H.F.; De Mars, R.; Long, E.O.; et al. T cell response to myelin basic protein in the context of the multiple sclerosis-associated HLA-DR15 haplotype: Peptide binding, immunodominance and effector functions of T cells. J. Neuroimmunol. 1997, 77, 195–203. [Google Scholar] [CrossRef]
- Wallström, E.; Khademi, M.; Andersson, M.; Weissert, R.; Linington, C.; Olsson, T. Increased reactivity to myelin oligodendrocyte glycoprotein peptides and epitope mapping in HLA DR2(15)+ multiple sclerosis. Eur. J. Immunol. 1998, 28, 3329–3335. [Google Scholar] [CrossRef]
- Pette, M.; Fujita, K.; Wilkinson, D.; Altmann, D.M.; Trowsdale, J.; Giegerich, G.; Hinkkanen, A.; Epplen, J.T.; Kappos, L.; Wekerle, H. Myelin autoreactivity in multiple sclerosis: Recognition of myelin basic protein in the context of HLA-DR2 products by T lymphocytes of multiple-sclerosis patients and healthy donors. Proc. Natl. Acad. Sci. USA 1990, 87, 7968–7972. [Google Scholar] [CrossRef] [Green Version]
- Jingwu, Z.; Medaer, R.; Hashim, G.A.; Chin, Y.; van den Berg-Loonen, E.; Raus, J.C. Myelin basic protein-specific T lymphocytes in multiple sclerosis and controls: Precursor frequency, fine specificity, and cytotoxicity. Ann. Neurol. 1992, 32, 330–338. [Google Scholar] [CrossRef]
- Bronge, M.; Ruhrmann, S.; Carvalho-Queiroz, C.; Nilsson, O.B.; Kaiser, A.; Holmgren, E.; Macrini, C.; Winklmeier, S.; Meinl, E.; Brundin, L.; et al. Myelin oligodendrocyte glycoprotein revisited-sensitive detection of MOG-specific T-cells in multiple sclerosis. J. Autoimmun. 2019, 102, 38–49. [Google Scholar] [CrossRef]
- Olsson, T.; Barcellos, L.F.; Alfredsson, L. Interactions between genetic, lifestyle and environmental risk factors for multiple sclerosis. Nat. Rev. Neurol. 2017, 13, 25–36. [Google Scholar] [CrossRef]
- Waubant, E.; Lucas, R.; Mowry, E.; Graves, J.; Olsson, T.; Alfredsson, L.; Langer-Gould, A. Environmental and genetic risk factors for MS: An integrated review. Ann. Clin. Transl. Neurol. 2019, 6, 1905–1922. [Google Scholar] [CrossRef] [PubMed]
- Lysandropoulos, A.P.; Racapé, J.; Holovska, V.; Toungouz, M. Human leucocyte antigen (HLA) class I and II typing in Belgian multiple sclerosis patients. Acta Neurol. Belg. 2017, 117, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Luomala, M.; Elovaara, I.; Ukkonen, M.; Koivula, T.; Lehtimäki, T. The combination of HLA-DR1 and HLA-DR53 protects against MS. Neurology 2001, 56, 383–385. [Google Scholar] [CrossRef] [PubMed]
- Creary, L.E.; Mallempati, K.C.; Gangavarapu, S.; Caillier, S.J.; Oksenberg, J.R.; Fernández-Viňa, M.A. Deconstruction of HLA-DRB1*04:01:01 and HLA-DRB1*15:01:01 class II haplotypes using next-generation sequencing in European-Americans with multiple sclerosis. Mult. Scler. 2018, 25, 772–782. [Google Scholar] [CrossRef] [PubMed]
- Caillier, S.J.; Briggs, F.; Cree, B.A.; Baranzini, S.E.; Fernandez-Viña, M.; Ramsay, P.P.; Khan, O.; Royal, W., 3rd; Hauser, S.L.; Barcellos, L.F.; et al. Uncoupling the roles of HLA-DRB1 and HLA-DRB5 genes in multiple sclerosis. J. Immunol. 2008, 181, 5473–5480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brum, D.G.; Barreira, A.A.; dos Santos, A.C.; Kaimen-Maciel, D.R.; Matiello, M.; Costa, R.M.; Deghaide, N.H.; Costa, L.S.; Louzada-Junior, P.; Diniz, P.R.; et al. HLA-DRB association in neuromyelitis optica is different from that observed in multiple sclerosis. Mult. Scler. 2010, 16, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Grau-López, L.; Raïch, D.; Ramo-Tello, C.; Naranjo-Gómez, M.; Dávalos, A.; Pujol-Borrell, R.; Borràs, F.E.; Martínez-Cáceres, E. Specific T-cell proliferation to myelin peptides in relapsing-remitting multiple sclerosis. Eur. J. Neurol. 2011, 18, 1101–1104. [Google Scholar] [CrossRef]
- Streeter, H.B.; Rigden, R.; Martin, K.F.; Scolding, N.J.; Wraith, D.C. Preclinical development and first-in-human study of ATX-MS-1467 for immunotherapy of MS. Neurol. Neuroimmunol. NeuroInflamm. 2015, 2, e93. [Google Scholar] [CrossRef] [Green Version]
- Anderton, S.M.; Viner, N.J.; Matharu, P.; Lowrey, P.A.; Wraith, D.C. Influence of a dominant cryptic epitope on autoimmune T cell tolerance. Nat. Immunol. 2002, 3, 175–181. [Google Scholar] [CrossRef]
- Hellings, N.; Barée, M.; Verhoeven, C.; D’hooghe, M.B.; Medaer, R.; Bernard, C.C.; Raus, J.; Stinissen, P. T-cell reactivity to multiple myelin antigens in multiple sclerosis patients and healthy controls. J. Neurosci. Res. 2001, 63, 290–302. [Google Scholar] [CrossRef]
- Van der Aa, A.; Hellings, N.; Medaer, R.; Gelin, G.; Palmers, Y.; Raus, J.; Stinissen, P. T cell vaccination in multiple sclerosis patients with autologous CSF-derived activated T cells: Results from a pilot study. Clin. Exp. Immunol. 2003, 131, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Link, H.; Olsson, T.; Xiao, B.G.; Andersson, G.; Ekre, H.P.; Linington, C.; Diener, P. T and B cell responses to myelin-oligodendrocyte glycoprotein in multiple sclerosis. J. Immunol. 1991, 146, 1490–1495. [Google Scholar] [PubMed]
- Sun, J.B.; Rask, C.; Olsson, T.; Holmgren, J.; Czerkinsky, C. Treatment of experimental autoimmune encephalomyelitis by feeding myelin basic protein conjugated to cholera toxin B subunit. Proc. Natl. Acad. Sci. USA 1996, 93, 7196–7201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raddassi, K.; Kent, S.C.; Yang, J.; Bourcier, K.; Bradshaw, E.M.; Seyfert-Margolis, V.; Nepom, G.T.; Kwok, W.W.; Hafler, D.A. Increased frequencies of myelin oligodendrocyte glycoprotein/MHC class II-binding CD4 cells in patients with multiple sclerosis. J. Immunol. 2011, 187, 1039–1046. [Google Scholar] [CrossRef] [Green Version]
- Tejada-Simon, M.V.; Hong, J.; Rivera, V.M.; Zhang, J.Z. Reactivity pattern and cytokine profile of T cells primed by myelin peptides in multiple sclerosis and healthy individuals. Eur. J. Immunol. 2001, 31, 907–917. [Google Scholar] [CrossRef]
- Bielekova, B.; Sung, M.H.; Kadom, N.; Simon, R.; McFarland, H.; Martin, R. Expansion and functional relevance of high-avidity myelin-specific CD4+ T cells in multiple sclerosis. J. Immunol. 2004, 172, 3893–3904. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Goods, B.A.; Raddassi, K.; Nepom, G.T.; Kwok, W.W.; Love, J.C.; Hafler, D.A. Functional inflammatory profiles distinguish myelin-reactive T cells from patients with multiple sclerosis. Sci. Transl. Med. 2015, 7, 287ra74. [Google Scholar] [CrossRef] [Green Version]
- Lees, J.R.; Cross, A.H. A little stress is good: IFN-gamma, demyelination, and multiple sclerosis. J. Clin. Investig. 2007, 117, 297–299. [Google Scholar] [CrossRef] [Green Version]
- Berthelot, L.; Laplaud, D.A.; Pettré, S.; Ballet, C.; Michel, L.; Hillion, S.; Braudeau, C.; Connan, F.; Lefrère, F.; Wiertlewski, S.; et al. Blood CD8+ T cell responses against myelin determinants in multiple sclerosis and healthy individuals. Eur. J. Immunol. 2008, 38, 1889–1899. [Google Scholar] [CrossRef]
- Martin, R.; Jaraquemada, D.; Flerlage, M.; Richert, J.; Whitaker, J.; Long, E.O.; McFarlin, D.E.; McFarland, H.F. Fine specificity and HLA restriction of myelin basic protein-specific cytotoxic T cell lines from multiple sclerosis patients and healthy individuals. J. Immunol. 1990, 145, 540–548. [Google Scholar]


{kind=link}
| MS Patients | Healthy Controls | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Code | Gender | Age | Disease Duration (Years) | EDSS Score | Current Treatment (Treatment Duration in Years) | Active MS | Code | Gender | Age |
| 100 | male | 62 | 25 | 6.5 | glatiramer acetate (16) | no | V1 | male | 23 |
| 101 | female | 69 | 32 | 6.0 | - | no | V2 | male | 46 |
| 102 | female | 39 | 12 | 1.5 | dimethyl fumarate (0.5) | no | V3 | female | 39 |
| 103 | female | 49 | 14 | 3.0 | - | yes | V4 | male | 27 |
| 104 | female | 52 | 4 | 2.5 | glatiramer acetate (0.5) | no | V5 | female | 50 |
| 105 | male | 31 | 10 | 1.0 | interferon-β (9) | no | V6 | female | 56 |
| 106 | female | 66 | 23 | 2.0 | glatiramer acetate (24) | no | V7 | female | 22 |
| 107 | female | 27 | 3 | 2.0 | - | yes | V8 | female | 34 |
| 108 | female | 24 | 5 | 0 | dimethyl fumarate (5) | no | V9 | female | 64 |
| 109 | male | 62 | 19 | 2.5 | interferon-β (19) | no | V10 | male | 64 |
| 110 | male | 53 | 16 | 3.0 | interferon-β (8) | no | |||
| 112 | male | 24 | 2 | 2.5 | dimethyl fumarate (2) | no | |||
| 113 | female | 27 | 11 | 2.0 | dimethyl fumarate (3) | no | |||
| 114 | male | 46 | 1 | 2.0 | - | no | |||
| 115 | male | 50 | 1 | 1.0 | dimethyl fumarate (1) | no | |||
| 116 | female | 30 | 5 | 1.0 | dimethyl fumarate (3) | no | |||
| 117 | male | 53 | 16 | 3.5 | - | no | |||
| 118 | female | 28 | 1 | 0 | - | no | |||
| 119 | female | 41 | 1 | 2.0 | dimethyl fumarate (1) | no | |||
| 121 | female | 42 | 9 | 2.0 | glatiramer acetate (9) | no | |||
| Median (range) | 44 (24–69) | 9.5 (1–32) | 2.0 (0–6.5) | 42.5 (22–64) | |||||
| Subject Code | MOG1–20 | MOG35–55 | MOG64–86 | MOG74–96 | MBP13–32 | MBP30–44 | MBP34–56 | MBP111–129 | MBP131–145 | MBP146–170 | PLP139–154 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MS patients | 100 | - | - | - | x | - | - | - | - | - | - | - |
| 101 | - | - | x | x | - | - | - | - | - | - | - | |
| 102 | - | - | - | - | x | - | x | - | - | - | - | |
| 103 | - | - | x | - | - | - | - | - | - | - | - | |
| 104 | - | - | - | - | - | - | - | - | - | - | - | |
| 105 | - | - | - | - | - | - | - | - | - | - | - | |
| 106 | - | - | - | - | - | - | x | - | - | - | - | |
| 107 | x | x | x | - | - | - | - | - | - | - | - | |
| 108 | - | x | - | x | - | - | x | - | - | - | - | |
| 109 | - | - | x | x | - | - | - | - | - | x | - | |
| 110 | - | - | x | x | - | - | - | - | - | - | - | |
| 112 | - | - | - | - | - | - | - | - | - | - | - | |
| 113 | - | - | - | - | - | - | - | - | - | - | - | |
| 114 | - | - | x | - | - | - | - | - | - | - | - | |
| 115 | - | - | - | - | - | - | - | - | - | - | - | |
| 116 | - | - | - | - | - | - | - | - | - | - | - | |
| 117 | x | - | - | - | - | - | - | - | - | - | - | |
| 118 | - | - | - | - | - | - | - | - | - | - | - | |
| 119 | x | x | - | x | x | - | x | - | - | x | x | |
| 121 | - | - | x | - | - | - | - | - | - | - | - | |
| Total n° of responses | 3 | 3 | 7 | 6 | 2 | 0 | 4 | 0 | 0 | 2 | 1 | |
| Healthy controls | V1 | - | x | x | - | - | - | - | - | - | - | - |
| V2 | - | - | - | - | - | - | - | - | - | - | x | |
| V3 | - | - | - | - | - | - | - | - | - | - | - | |
| V4 | x | - | x | x | - | - | - | - | - | - | - | |
| V5 | - | - | - | - | - | - | - | - | - | - | - | |
| V6 | - | - | - | - | - | x | - | - | - | - | - | |
| V7 | - | x | x | x | x | x | x | x | x | x | x | |
| V8 | - | - | - | - | - | - | - | - | - | - | - | |
| V9 | - | - | - | - | - | - | - | - | - | - | - | |
| V10 | - | - | x | - | - | - | - | - | - | - | - | |
| Total n° of responses | 1 | 2 | 4 | 2 | 1 | 2 | 1 | 1 | 1 | 1 | 2 |
| Code | DRB1 | DQA1 | DQB1 | DRB3 | DRB4 | DRB5 | DPA1 | DPB1 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MS patients | 100 | *03:01 | *01:01 | *05:01 | *01:01 | *02:01 | *05:01 | *02:02 | - | - | - | *01:03 | *01:03 | *04:01 | *02:01 |
| 101 | *13:02 | *08:03 | *01:02 | *06:01 | *06:09 | *03:01 | *03:01 | - | - | - | *01:03 | - | *02:01 | *16:01 | |
| 102 | *03:01 | *01:02 | *05:01 | *01:01 | *02:01 | *05:01 | *01:01 | - | - | - | *02:01 | *01:03 | *17:01 | *02:01 | |
| 103 | *15:01 | *03:01 | *01:02 | *05:01 | *06:03 | *02:01 | *01:01 | - | - | *01:01 | *02:01 | *01:03 | *09:01 | *04:01 | |
| 104 | *11:01 | *15:01 | *05:09 | *01:02 | *03:01 | *06:02 | *02:02 | - | - | *01:01 | *01:03 | *02:06 | *03:01 | *05:01 | |
| 105 | *15:01 | *01:01 | *01:02 | *01:01 | *06:02 | *05:01 | - | - | - | *01:01 | *01:03 | *02:01 | *04:02 | *14:01 | |
| 106 | *15:01 | - | *01:02 | - | *06:02 | - | - | - | - | *01:01 | *01:03 | - | *04:01 | - | |
| 107 | *15:01 | *01:02 | *01:02 | *01:01 | *05:01 | *06:02 | - | - | - | *01:01 | *01:03 | *01:03 | *04:02 | *02:01 | |
| 108 | *15:01 | *08:XX | *01:02 | *04:01 | *06:02 | *04:02 | - | - | - | *01:01 | *01:03 | - | *04:01 | - | |
| 109 | *11:01 | *01:01 | *05:05 | *01:01 | *03:01 | *05:01 | *02:02 | - | - | - | *01:03 | *01:03 | *04:02 | *04:01 | |
| 110 | *12:01 | *01:01 | *05:05 | *01:01 | *03:01 | *05:01 | *02:02 | - | - | - | *01:03 | *01:03 | *02:01 | *03:01 | |
| 112 | *11:02 | *10:01 | *05:05 | *01:05 | *03:19 | *05:01 | *02:02 | - | - | - | *01:03 | *02:01 | *04:01 | *30:01 | |
| 113 | *07:01 | *15:01 | *02:01 | *01:02 | *02:02 | *06:02 | - | - | *01:01 | *01:01 | *01:03 | *02:01 | *04:01 | *11:01 | |
| 114 | *13:02 | *03:01 | *01:02 | *05:01 | *06:04 | *02:01 | *03:01 | *01:01 | - | - | *02:01 | *01:03 | *01:01 | *03:01 | |
| 115 | *03:01 | *15:01 | *05:01 | *01:02 | *02:01 | *06:02 | *01:01 | - | - | *01:01 | *02:01 | *01:03 | *01:01 | *04:01 | |
| 116 | *15:01 | *08:01 | *01:02 | *04:01 | *06:02 | *04:02 | - | - | - | *01:01 | *01:03 | *01:03 | *04:02 | - | |
| 117 | *04:01 | *03:01 | *03:01 | *05:01 | *03:02 | *02:01 | *01:01 | - | *01:03 | - | *01:03 | *01:03 | *04:01 | - | |
| 118 | *11:04 | *10:01 | *05:05 | *01:05 | *03:01 | *05:01 | *02:02 | - | - | - | *01:03 | *01:03 | *02:01 | *04:02 | |
| 119 | *07:01 | *01:01 | *02:01 | *01:02 | *03:03 | *05:04 | - | - | *01:03:01:02N | - | *01:03 | *01:03 | *02:01 | *04:01 | |
| 121 | *03:01 | *13:02 | *05:01 | *01:02 | *02:01 | *06:04 | *01:01 | *03:01 | - | - | *01:03 | *02:02 | *04:01 | *01:01 | |
| Healthy controls | V1 | *07:01 | *03:01 | *02:01 | *05:01 | *02:02 | *02:01 | *01:01 | - | *01:03 | - | *02:01 | *01:03 | - | - |
| V2 | *07:01 | *12:01 | *02:01 | *05:05 | *03:03 | *03:01 | *02:02 | - | *01:03:01:02N | - | *01:03 | *01:03 | - | - | |
| V3 | *14:54 | *07:01 | *01:04 | *02:01 | *05:03 | *02:02 | *02:02 | - | *01:01 | - | *01:03 | *01:03 | - | - | |
| V4 | *03:01 | *01:01 | *05:01 | *01:01 | *02:01 | *05:01 | *02:02 | - | - | - | *01:03 | *01:03 | - | - | |
| V5 | *07:01 | *03:01 | *02:01 | *05:01 | *02:02 | *02:01 | *01:01 | - | *01:03:01:02N | - | *02:01 | *01:03 | *11:01 | *04:02 | |
| V6 | *16:01 | *04:01 | *01:02 | *03:03 | *05:02 | *03:02 | - | - | *01:03 | *02:02 | *02:01 | *01:03 | *01:01 | *04:01 | |
| V7 | *13:01 | - | *01:03 | - | *06:03 | - | *02:02 | *01:01 | - | - | *01:03 | *02:02 | *04:02 | *05:01 | |
| V8 | *07:01 | *03:01 | *02:01 | *05:01 | *02:02 | *02:01 | *01:01 | - | *01:03 | - | *01:03 | - | *04:01 | - | |
| V9 | *03:01 | *15:01 | *05:01 | *01:02 | *02:01 | *06:02 | *01:01 | - | - | *01:01 | *02:01 | *01:03 | - | - | |
| V10 | *13:05 | *14:54 | *05:05 | *01:04 | *03:01 | *05:03 | *02:02 | *02:02 | - | - | *01:03 | *02:01 | - | - | |
| HLA-DR15 | HLA-DRB1*15:01 | HLA-DQA1*01:02 | HLA-DQB1*06:02 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| + (9/30) | - (21/30) | + (10/30) | - (20/30) | + (15/30) | - (15/30) | + (9/30) | - (21/30) | ||
| MOG | |||||||||
| All MOG | Response | 2 | 13 | 3 | 12 | 6 | 9 | 2 | 13 |
| MOG1–20 | Response | 1 | 3 | 1 | 3 | 2 | 2 | 1 | 3 |
| Frequency | 78.5 ± 185.8 | 49.2 ± 50.4 | 73.0 ± 176.0 | 50.5 ± 51.3 | 68.3 ± 147.0 | 47.7 ± 43.7 | 78.5 ± 185.8 | 49.2 ± 50.4 | |
| MOG35–55 | Response | 2 | 2 | 2 | 3 | 3 | 2 | 2 | 2 |
| Frequency | 88.9 ± 155.1 | 45.5 ± 49.0 | 81.5 ± 148.1 | 47.0 ± 49.8 | 68.0 ± 121.1 | 49.0 ± 56.4 | 88.9 ± 155.1 | 45.5 ± 49.0 | |
| MOG64–86 | Response | 1 | 10 | 2 | 9 | 5 | 6 | 1 | 10 |
| Frequency | 39.5 ± 60.4 | 91.5 ± 108.8 | 50.6 ± 66.8 | 68.2 ± 93.3 | 47.7 ± 55.2 | 104.1 ± 124.3 | 39.5 ± 60.4 | 91.5 ± 108.8 | |
| MOG74–96 | Response | 1 | 7 | 1 | 7 | 3 | 5 | 1 | 7 |
| Frequency | 40.7 ± 52.7 | 45.8 ± 34.7 | 43.3 ± 50.3 | 49.3 ± 41.5 | 45.7 ± 45.6 | 42.9 ± 35.0 | 40.7 ± 52.7 | 45.8 ± 34.7 | |
| MBP | |||||||||
| All MBP | Response | 2 | 5 | 2 | 5 | 4 | 3 | 2 | 5 |
| MBP13–32 | Response | 0 | 3 | 0 | 3 | 1 | 2 | 0 | 3 |
| Frequency | 20.1 ± 18.4 | 39.8 ± 32.1 | 23.4 ± 20.3 | 30.8 ± 20.4 | 25.9 ± 21.0 | 41.8 ± 35.6 | 20.1 ± 18.4 | 39.8 ± 32.1 | |
| MBP30–44 | Response | 0 | 2 | 0 | 2 | 1 | 1 | 0 | 2 |
| Frequency | 21.1 ± 19.1 | 27.2 ± 19.3 | 20.8 ± 18.1 | 26.0 ±18.0 | 19.5 ± 15.1 | 31.2 ± 21.4 | 21.1 ± 19.1 | 27.2 ± 19.3 | |
| MBP34–56 | Response | 2 | 3 | 2 | 3 | 3 | 2 | 2 | 3 |
| Frequency | 40.8 ± 45.2 | 30.6 ± 22.7 | 40.9 ± 42.6 | 32.5 ± 23.5 | 41.3 ± 39.8 | 26.1 ± 15.5 | 40.8 ± 45.2 | 30.6 ± 22.7 | |
| MBP111–129 | Response | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 |
| Frequency | 22.0 ± 15.9 | 24.4 ± 15.3 | 21.8 ±15.0 | 23.3 ± 15.2 | 19.4 ± 12.8 | 28.0 ± 16.6 | 22.0 ± 15.9 | 24.4 ± 15.3 | |
| MBP131–145 | Response | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 |
| Frequency | 27.4 ± 23.8 | 25.0 ± 17.2 | 26.1 ± 22.7 | 25.1 ±17.6 | 23.4 ± 19.0 | 28.0 ± 19.4 | 27.4 ± 23.8 | 25.0 ± 17.2 | |
| MBP146–170 | Response | 0 | 3 | 0 | 3 | 1 | 2 | 0 | 3 |
| Frequency | 22.9 ± 21.6 | 28.7 ± 23.4 | 22.0 ± 20.6 | 30.4 ± 24.1 | 25.5 ± 28.0 | 28.3 ± 16.7 | 22.9 ± 21.6 | 28.7 ± 23.4 | |
| PLP | |||||||||
| PLP139–154 | Response | 0 | 3 | 0 | 3 | 1 | 2 | 0 | 3 |
| Frequency | 26.6 ± 14.6 | 30.6 ± 20.6 | 25.6 ± 14.1 | 33.3 ± 20.7 | 29.1 ± 21.7 | 29.8 ± 16.3 | 26.6 ± 14.6 | 30.6 ± 20.6 | |
| All peptides | |||||||||
| Response to at least one peptide | 3 | 16 | 4 | 15 | 9 | 10 | 3 | 16 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Derdelinckx, J.; Nkansah, I.; Ooms, N.; Van Bruggen, L.; Emonds, M.-P.; Daniëls, L.; Reynders, T.; Willekens, B.; Cras, P.; Berneman, Z.N.; et al. HLA Class II Genotype Does Not Affect the Myelin Responsiveness of Multiple Sclerosis Patients. Cells 2020, 9, 2703. https://doi.org/10.3390/cells9122703
Derdelinckx J, Nkansah I, Ooms N, Van Bruggen L, Emonds M-P, Daniëls L, Reynders T, Willekens B, Cras P, Berneman ZN, et al. HLA Class II Genotype Does Not Affect the Myelin Responsiveness of Multiple Sclerosis Patients. Cells. 2020; 9(12):2703. https://doi.org/10.3390/cells9122703
Chicago/Turabian StyleDerdelinckx, Judith, Irene Nkansah, Naomi Ooms, Laura Van Bruggen, Marie-Paule Emonds, Liesbeth Daniëls, Tatjana Reynders, Barbara Willekens, Patrick Cras, Zwi N. Berneman, and et al. 2020. "HLA Class II Genotype Does Not Affect the Myelin Responsiveness of Multiple Sclerosis Patients" Cells 9, no. 12: 2703. https://doi.org/10.3390/cells9122703