In Vitro Studies to Define the Cell-Surface and Intracellular Targets of Polyarginine-Conjugated Sodium Borocaptate as a Potential Delivery Agent for Boron Neutron Capture Therapy



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Lentivirus Preparation
2.2. Sequence of shRNA and Oligonucleotides
2.3. Boron Compounds and Boron Content Measurement
2.4. Immunomagnetic Positive Selection of BSH-11R-Binding Cells
2.5. Immunofluorescent Analysis and Proximity Ligation Assay
2.6. Western Blotting
2.7. Immunoprecipitation
2.8. X-ray Irradiation and Sphere Formation Assay
2.9. Neutron Irradiation
2.10. Gene Expression Analyses
3. Results
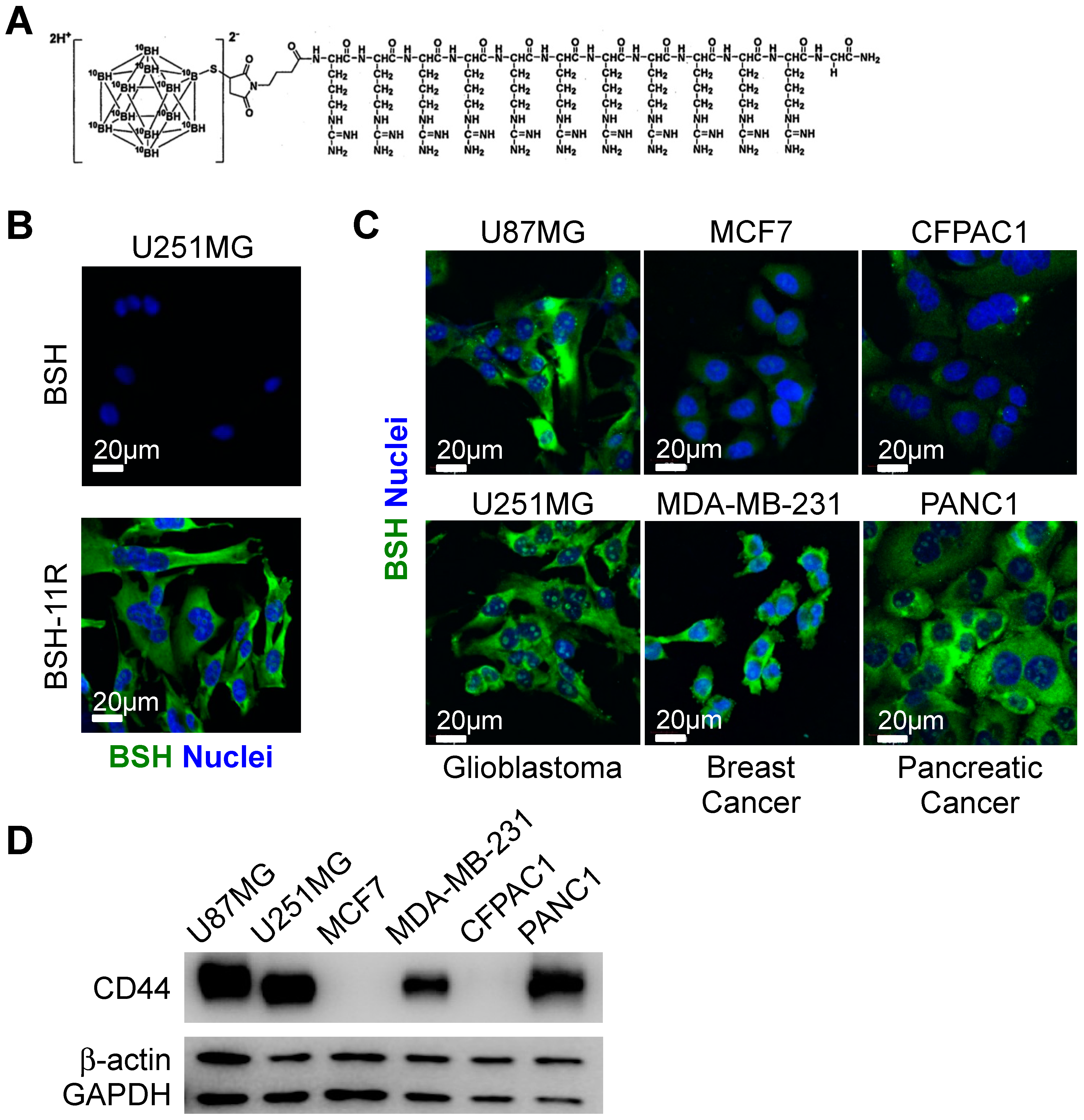
3.1. Cellular Uptake of BSH-polyR in Various Types of Cancers
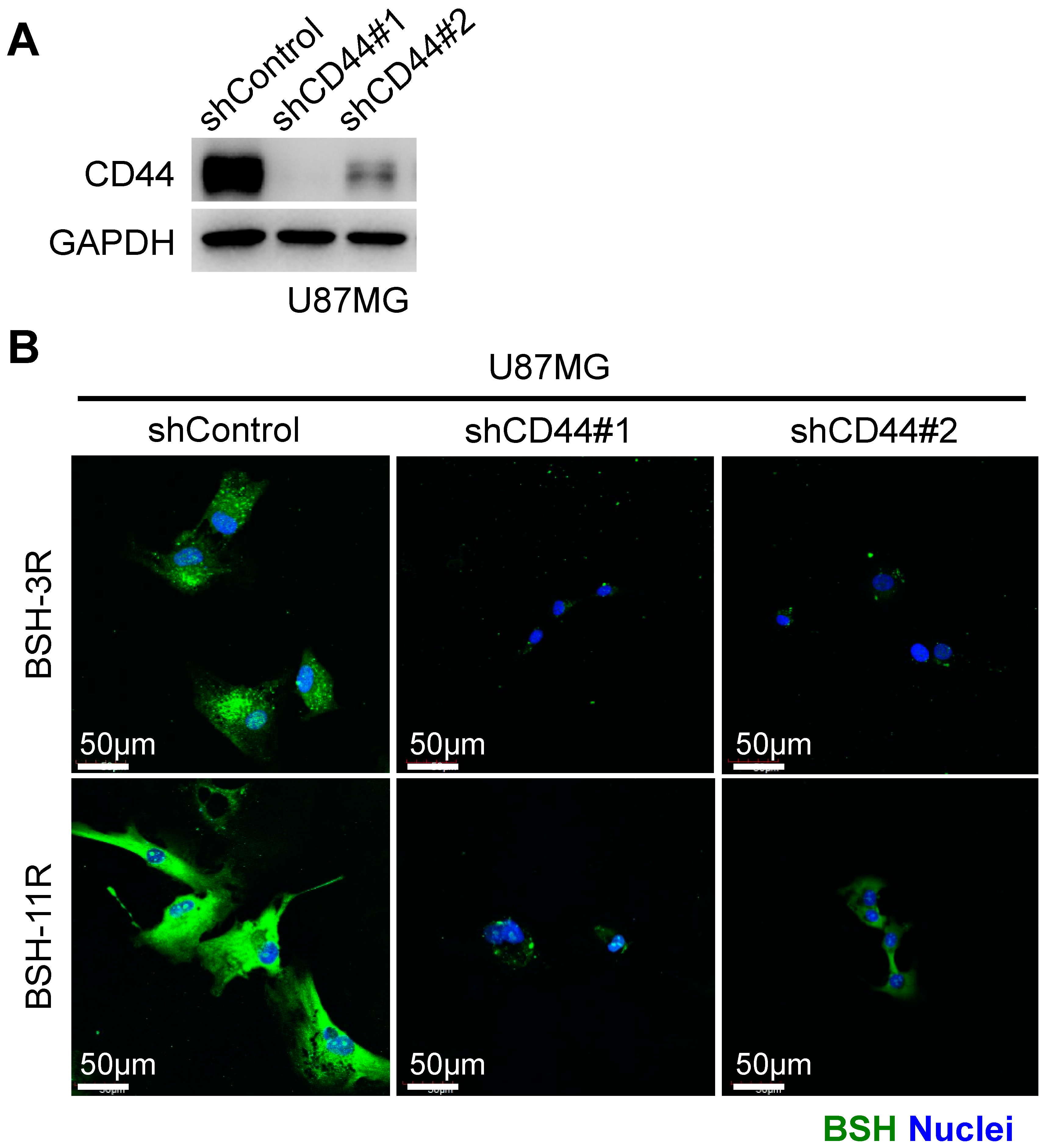
3.2. CD44 Is Required for BSH-PolyR Cellular Uptake
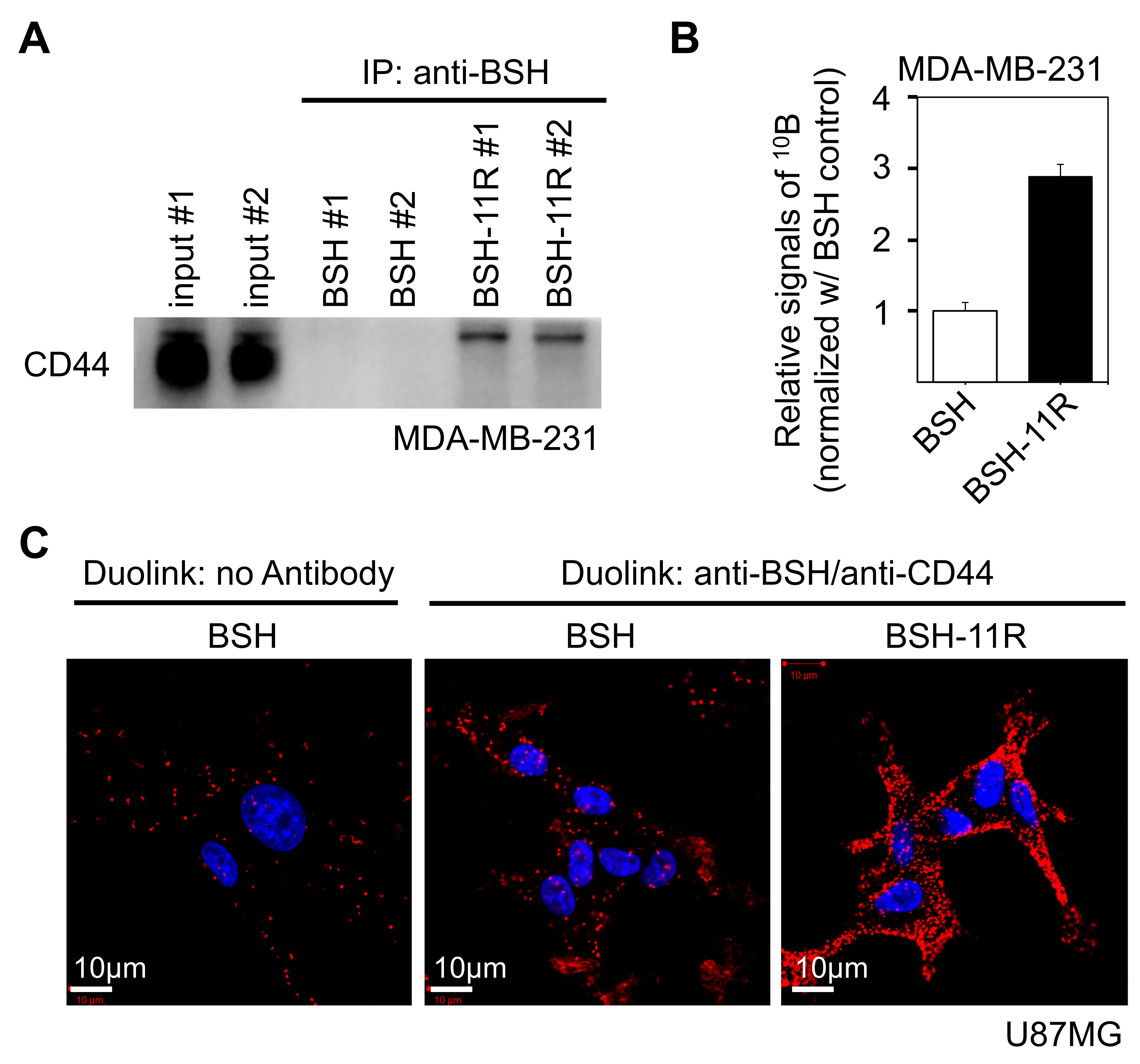
3.3. Direct Interaction between BSH-PolyR and CD44 Cell-Surface Protein
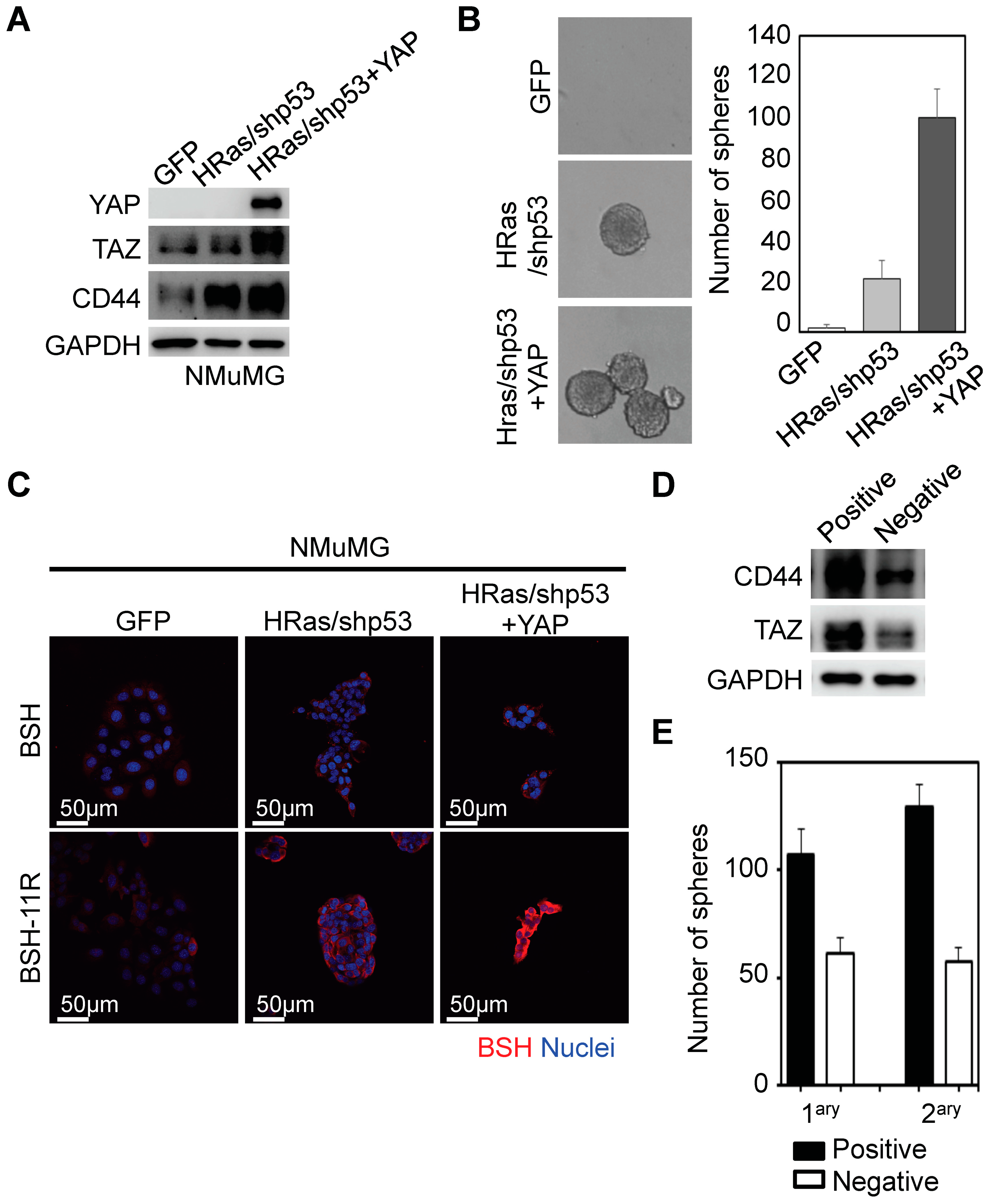
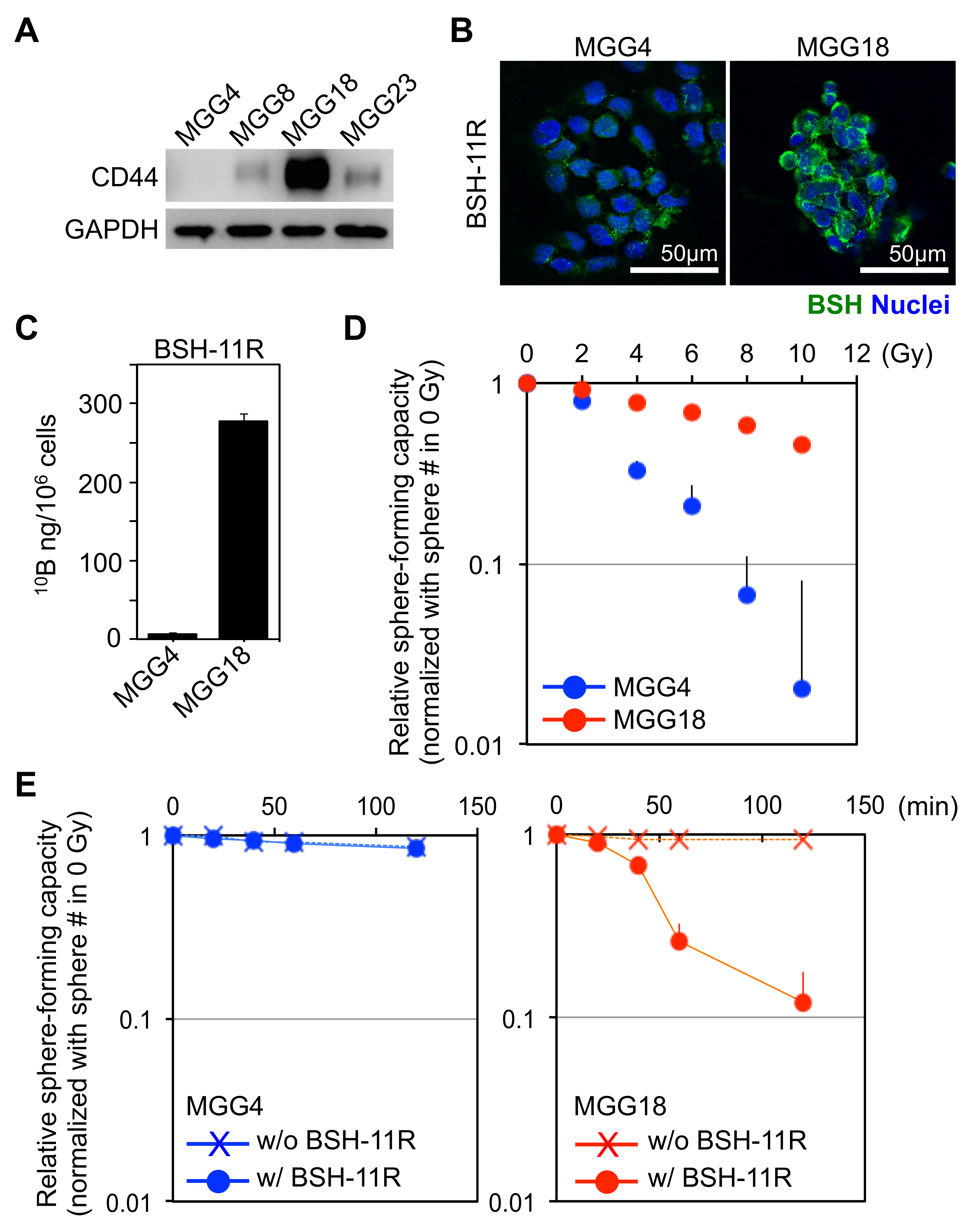
3.4. BSH-11R Is Efficiently Delivered to Cancer Stem-Like Cells
3.5. BSH-PolyR-Based BNCT Is Effective against CD44High Glioma Stem-Like Cells
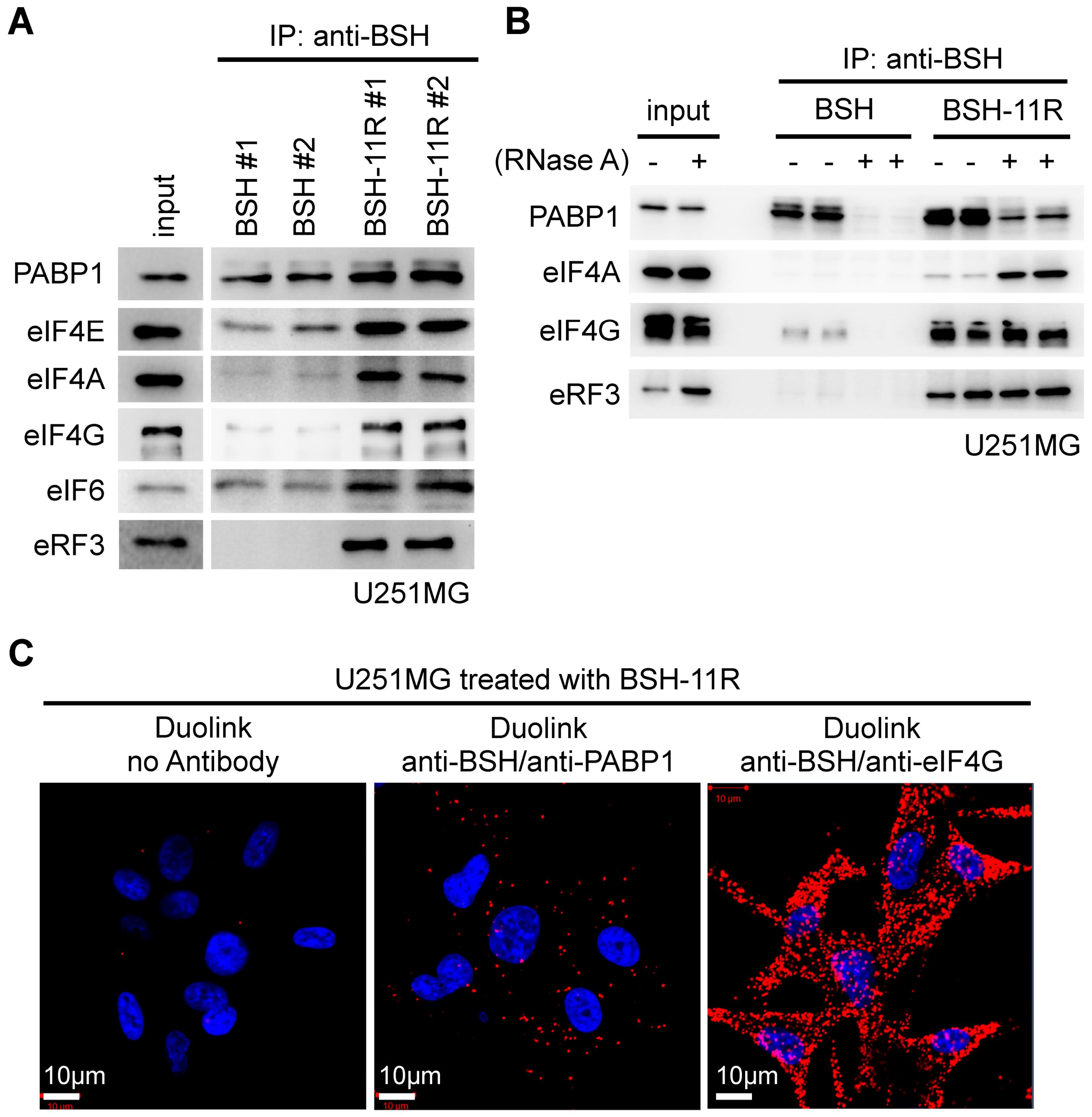
3.6. Intracellular Target of BSH-11R
3.7. Molecular Properties of BSH-PolyR Define the Intracellular Retention Time of BSH-PolyR
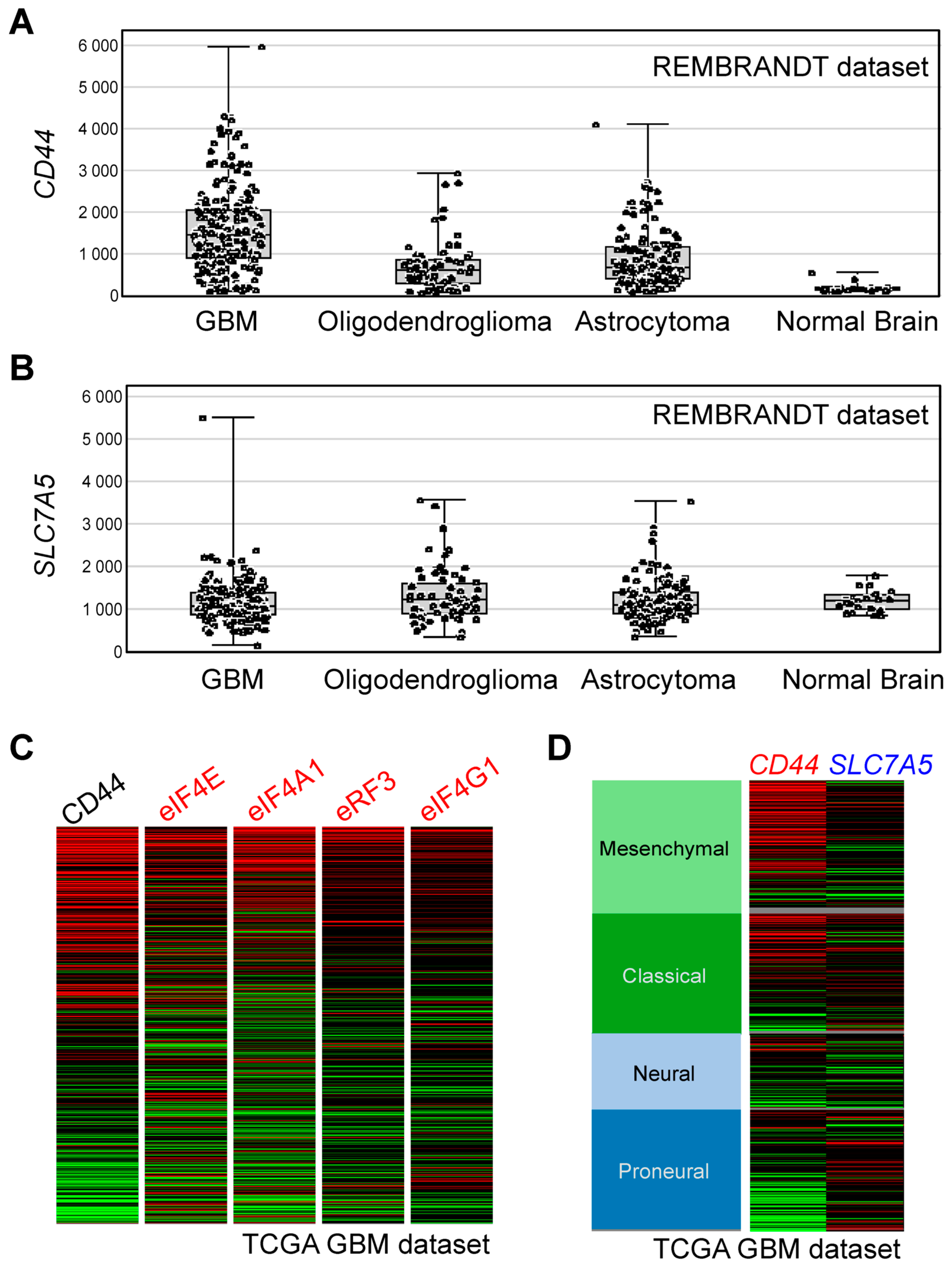
3.8. Bioinformatics Analyses
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sauerwein, W. Principles and Roots of Neutron Capture Therapy. Neutron Capture Ther. 2012, 169, 1–16. [Google Scholar] [CrossRef]
- Moss, R. Critical review, with an optimistic outlook, on Boron Neutron Capture Therapy (BNCT). Appl. Radiat. Isot. 2014, 88, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M. Boron neutron capture therapy (BNCT): A unique role in radiotherapy with a view to entering the accelerator-based BNCT era. Int. J. Clin. Oncol. 2019, 25, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Detta, A.; Cruickshank, G.S. l-Amino Acid Transporter-1 and Boronophenylalanine-Based Boron Neutron Capture Therapy of Human Brain Tumors. Cancer Res. 2009, 69, 2126–2132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wongthai, P.; Hagiwara, K.; Miyoshi, Y.; Wiriyasermkul, P.; Wei, L.; Ohgaki, R.; Kato, I.; Hamase, K.; Nagamori, S.; Kanai, Y. Boronophenylalanine, a boron delivery agent for boron neutron capture therapy, is transported by ATB0,+, LAT1 and LAT2. Cancer Sci. 2015, 106, 279–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, R.F.; Vicente, M.G.H.; Harling, O.K.; Kiger, W.; Riley, K.J.; Binns, P.J.; Wagner, F.M.; Suzuki, M.; Aihara, T.; Kato, I.; et al. Current status of boron neutron capture therapy of high grade gliomas and recurrent head and neck cancer. Radiat. Oncol. 2012, 7, 146. [Google Scholar] [CrossRef] [Green Version]
- Hatanaka, H. Experience of Boron-neutron Capture Therapy for Malignant Brain Tumours—With Special Reference to the Problems of Postoperative CT Follow-ups. In Proceedings of the 8th European Congress of Neurosurgery Barcelona, 6–11 September 1987; Springer: Vienna, Austria, 1988; Volume 42, pp. 187–192. [Google Scholar] [CrossRef]
- Mishima, Y.; Ichihashi, M.; Tsuji, M.; Hatta, S.; Ueda, M.; Honda, C.; Suzuki, T. Treatment of Malignant Melanoma by Selective Thermal Neutron Capture Therapy Using Melanoma-Seeking Compound. J. Investig. Dermatol. 1989, 92, 321S–325S. [Google Scholar] [CrossRef] [Green Version]
- Kawabata, S.; Miyatake, S.-I.; Nonoguchi, N.; Hiramatsu, R.; Iida, K.; Miyata, S.; Yokoyama, K.; Doi, A.; Kuroda, Y.; Kuroiwa, T.; et al. Survival benefit from boron neutron capture therapy for the newly diagnosed glioblastoma patients. Appl. Radiat. Isot. 2009, 67, S15–S18. [Google Scholar] [CrossRef]
- Takeuchi, K.; Hattori, Y.; Kawabata, S.; Futamura, G.; Hiramatsu, R.; Wanibuchi, M.; Tanaka, H.; Masunaga, S.-I.; Ono, K.; Miyatake, S.-I.; et al. Synthesis and Evaluation of Dodecaboranethiol Containing Kojic Acid (KA-BSH) as a Novel Agent for Boron Neutron Capture Therapy. Cells 2020, 9, 1551. [Google Scholar] [CrossRef]
- Tsurubuchi, T.; Shirakawa, M.; Kurosawa, W.; Matsumoto, K.; Ubagai, R.; Umishio, H.; Suga, Y.; Yamazaki, J.; Arakawa, A.; Maruyama, Y.; et al. Evaluation of a Novel Boron-Containing α-d-Mannopyranoside for BNCT. Cells 2020, 9, 1277. [Google Scholar] [CrossRef]
- Satapathy, R.; Dash, B.P.; Bode, B.P.; Byczynski, E.A.; Hosmane, S.N.; Bux, S.; Hosmane, N.S. New classes of carborane-appended 5-thio-d-glucopyranose derivatives. Dalton Trans. 2012, 41, 8982–8988. [Google Scholar] [CrossRef]
- Zhu, Y.; Lin, Y.; Hosmane, N.S. Synthesis and in vitro anti-tumor activity of carboranyl levodopa. Bioorg. Chem. 2019, 90, 103090. [Google Scholar] [CrossRef]
- Iguchi, Y.; Michiue, H.; Kitamatsu, M.; Hayashi, Y.; Takenaka, F.; Nishiki, T.-I.; Matsui, H. Tumor-specific delivery of BSH-3R for boron neutron capture therapy and positron emission tomography imaging in a mouse brain tumor model. Biomaterials 2015, 56, 10–17. [Google Scholar] [CrossRef]
- Borrelli, A.; Tornesello, A.L.; Tornesello, M.L.; Buonaguro, L. Cell Penetrating Peptides as Molecular Carriers for Anti-Cancer Agents. Molecules 2018, 23, 295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A. Macropinocytosis: Searching for an endocytic identity and role in the uptake of cell penetrating peptides. J. Cell. Mol. Med. 2007, 11, 670–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, G.; Nakase, I.; Fukuda, Y.; Masuda, R.; Oishi, S.; Shimura, K.; Kawaguchi, Y.; Takatani-Nakase, T.; Langel, Ü.; Gräslund, A.; et al. CXCR4 Stimulates Macropinocytosis: Implications for Cellular Uptake of Arginine-Rich Cell-Penetrating Peptides and HIV. Chem. Biol. 2012, 19, 1437–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimura, A.; Michiue, H.; Cheng, Y.; Uneda, A.; Tani, Y.; Nishiki, T.-I.; Ichikawa, T.; Wei, F.-Y.; Tomizawa, K.; Matsui, H. Cyclin G2 Promotes Hypoxia- Driven Local Invasion of Glioblastoma by Orchestrating Cytoskeletal Dynamics. Neoplasia 2013, 15, 1272–1281. [Google Scholar] [CrossRef] [Green Version]
- Wakimoto, H.; Mohapatra, G.; Kanai, R.; Curry, W.T.; Yip, S.; Nitta, M.; Patel, A.P.; Barnard, Z.R.; Stemmer-Rachamimov, A.O.; Louis, D.N.; et al. Maintenance of primary tumor phenotype and genotype in glioblastoma stem cells. J. Neurooncol. 2011, 14, 132–144. [Google Scholar] [CrossRef] [Green Version]
- Michiue, H.; Sakurai, Y.; Kondo, N.; Kitamatsu, M.; Bin, F.; Nakajima, K.; Hirota, Y.; Kawabata, S.; Nishiki, T.-I.; Ohmori, I.; et al. The acceleration of boron neutron capture therapy using multi-linked mercaptoundecahydrododecaborate (BSH) fused cell-penetrating peptide. Biomaterials 2014, 35, 3396–3405. [Google Scholar] [CrossRef]
- Feng, B.; Tomizawa, K.; Michiue, H.; Miyatake, S.-I.; Han, X.-J.; Fujimura, A.; Seno, M.; Kirihata, M.; Matsui, H. Delivery of sodium borocaptate to glioma cells using immunoliposome conjugated with anti-EGFR antibodies by ZZ-His. Biomaterials 2009, 30, 1746–1755. [Google Scholar] [CrossRef]
- Yamamoto, T.; Fujimura, A.; Wei, F.-Y.; Shinojima, N.; Kuroda, J.-I.; Mukasa, A.; Tomizawa, K. 2-Methylthio Conversion of N6-Isopentenyladenosine in Mitochondrial tRNAs by CDK5RAP1 Promotes the Maintenance of Glioma-Initiating Cells. iScience 2019, 21, 42–56. [Google Scholar] [CrossRef] [Green Version]
- Fujimura, A.; Michiue, H.; Nishiki, T.-I.; Ohmori, I.; Wei, F.-Y.; Matsui, H.; Tomizawa, K. Expression of a Constitutively Active Calcineurin Encoded by an Intron-Retaining mRNA in Follicular Keratinocytes. PLoS ONE 2011, 6, e17685. [Google Scholar] [CrossRef]
- Uritani, A.; Menjo, Y.; Watanabe, K.; Yamazaki, A.; Kiyanagi, Y.; Tsuchida, K. Design of Beam Shaping Assembly for an Accelerator-driven BNCT System in Nagoya University. JPS Conf. Proc. 2018, 22, 011002–1–011002–7. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Uritani, A.; Watanabe, K.; Yoshihashi, S.; Yamazaki, A.; Kiyanagi, Y.; Tsuchida, K. Improved Design of the Exit of a Beam Shaping Assembly for an Accelerator-driven BNCT System in Nagoya. JPS Conf. Proc. 2018, 22, 011003–1–011003–7. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, M.; Tomizawa, K.; Moriwaki, A.; Li, S.-T.; Terada, H.; Matsui, H. A High-Efficiency Protein Transduction System Demonstrating the Role of PKA in Long-Lasting Long-Term Potentiation. J. Neurosci. 2001, 21, 6000–6007. [Google Scholar] [CrossRef] [Green Version]
- Wadia, J.S.; Stan, R.V.; Dowdy, S.F. Transducible TAT-HA fusogenic peptide enhances escape of TAT-fusion proteins after lipid raft macropinocytosis. Nat. Med. 2004, 10, 310–315. [Google Scholar] [CrossRef]
- Kaplan, I.M.; Wadia, J.S.; Dowdy, S.F. Cationic TAT peptide transduction domain enters cells by macropinocytosis. J. Control. Release 2005, 102, 247–253. [Google Scholar] [CrossRef]
- Patra, S.K. Dissecting lipid raft facilitated cell signaling pathways in cancer. Biochim. Biophys. Acta (BBA) Bioenerg. 2008, 1785, 182–206. [Google Scholar] [CrossRef]
- Johannes, L.; Billet, A. Glycosylation and raft endocytosis in cancer. Cancer Metastasis Rev. 2020, 39, 375–396. [Google Scholar] [CrossRef]
- Mollinedo, F.; Gajate, C. Lipid rafts as signaling hubs in cancer cell survival/death and invasion: Implications in tumor progression and therapy. J. Lipid Res. 2020, 61, 611–635. [Google Scholar] [CrossRef] [Green Version]
- Cordenonsi, M.; Zanconato, F.; Azzolin, L.; Forcato, M.; Rosato, A.; Frasson, C.; Inui, M.; Montagner, M.; Parenti, A.R.; Poletti, A.; et al. The Hippo Transducer TAZ Confers Cancer Stem Cell-Related Traits on Breast Cancer Cells. Cell 2011, 147, 759–772. [Google Scholar] [CrossRef]
- Kai, K.; Arima, Y.; Kamiya, T.; Saya, H. Breast cancer stem cells. Breast Cancer 2009, 17, 80–85. [Google Scholar] [CrossRef]
- Rambaruth, N.D.; Dwek, M.V. Cell surface glycan–lectin interactions in tumor metastasis. Acta Histochem. 2011, 113, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Senbanjo, L.T.; Chellaiah, M.A. CD44: A Multifunctional Cell Surface Adhesion Receptor Is a Regulator of Progression and Metastasis of Cancer Cells. Front. Cell Dev. Biol. 2017, 5, 18. [Google Scholar] [CrossRef] [Green Version]
- Skandalis, S.S.; Karalis, T.T.; Chatzopoulos, A.; Karamanos, N.K. Hyaluronan-CD44 axis orchestrates cancer stem cell functions. Cell. Signal. 2019, 63, 109377. [Google Scholar] [CrossRef]
- Donatello, S.; Babina, I.S.; Hazelwood, L.D.; Hill, A.D.; Nabi, I.R.; Hopkins, A.M. Lipid Raft Association Restricts CD44-Ezrin Interaction and Promotion of Breast Cancer Cell Migration. Am. J. Pathol. 2012, 181, 2172–2187. [Google Scholar] [CrossRef]
- Babina, I.S.; A McSherry, E.; Donatello, S.; Hill, A.; Hopkins, A.M. A novel mechanism of regulating breast cancer cell migration via palmitoylation-dependent alterations in the lipid raft affiliation of CD44. Breast Cancer Res. 2014, 16, R19. [Google Scholar] [CrossRef] [Green Version]
- Naor, D.; Sionov, R.V.; Ish-Shalom, D. CD44: Structure, Function and Association with the Malignant Process. Adv. Cancer Res. 1997, 71, 241–319. [Google Scholar] [CrossRef]
- Bourguignon, L.Y. CD44 isoform-cytoskeleton interaction in oncogenic signaling and tumor progression. Front. Biosci. 1998, 3, d637–d649. [Google Scholar] [CrossRef] [Green Version]
- Söderberg, O.; Gullberg, M.; Jarvius, M.; Ridderstråle, K.; Leuchowius, K.-J.; Jarvius, J.; Wester, K.; Hydbring, P.; Bahram, F.; Larsson, L.-G.; et al. Direct observation of individual endogenous protein complexes in situ by proximity ligation. Nat. Methods 2006, 3, 995–1000. [Google Scholar] [CrossRef]
- Gzil, A.; Zarębska, I.; Bursiewicz, W.; Antosik, P.; Grzanka, D.; Szylberg, Ł. Markers of pancreatic cancer stem cells and their clinical and therapeutic implications. Mol. Biol. Rep. 2019, 46, 6629–6645. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-C.; Yeh, C.-T.; Lin, K.-H. Cancer Stem Cell Functions in Hepatocellular Carcinoma and Comprehensive Therapeutic Strategies. Cells 2020, 9, 1331. [Google Scholar] [CrossRef] [PubMed]
- Zanconato, F.; Cordenonsi, M.; Piccolo, S. YAP/TAZ at the Roots of Cancer. Cancer Cell 2016, 29, 783–803. [Google Scholar] [CrossRef] [PubMed]
- Kovar, H.; Bierbaumer, L.; Radic-Sarikas, B. The YAP/TAZ Pathway in Osteogenesis and Bone Sarcoma Pathogenesis. Cells 2020, 9, 972. [Google Scholar] [CrossRef] [Green Version]
- Brogna, S.; Sato, T.-A.; Rosbash, M. Ribosome Components Are Associated with Sites of Transcription. Mol. Cell 2002, 10, 93–104. [Google Scholar] [CrossRef]
- McLeod, T.; Abdullahi, A.; Li, M.; Brogna, S. Recent studies implicate the nucleolus as the major site of nuclear translation. Biochem. Soc. Trans. 2014, 42, 1224–1228. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Makala, L.; Wu, D.; Cai, Y. Targeting translation: EIF4E as an emerging anticancer drug target. Expert Rev. Mol. Med. 2016, 18, 2. [Google Scholar] [CrossRef]
- Ramamurthy, V.P.; Ramalingam, S.; Kwegyir-Afful, A.K.; Hussain, A.; Njar, V.C. Targeting of protein translation as a new treatment paradigm for prostate cancer. Curr. Opin. Oncol. 2017, 29, 210–220. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujimura, A.; Yasui, S.; Igawa, K.; Ueda, A.; Watanabe, K.; Hanafusa, T.; Ichikawa, Y.; Yoshihashi, S.; Tsuchida, K.; Kamiya, A.; et al. In Vitro Studies to Define the Cell-Surface and Intracellular Targets of Polyarginine-Conjugated Sodium Borocaptate as a Potential Delivery Agent for Boron Neutron Capture Therapy. Cells 2020, 9, 2149. https://doi.org/10.3390/cells9102149
Fujimura A, Yasui S, Igawa K, Ueda A, Watanabe K, Hanafusa T, Ichikawa Y, Yoshihashi S, Tsuchida K, Kamiya A, et al. In Vitro Studies to Define the Cell-Surface and Intracellular Targets of Polyarginine-Conjugated Sodium Borocaptate as a Potential Delivery Agent for Boron Neutron Capture Therapy. Cells. 2020; 9(10):2149. https://doi.org/10.3390/cells9102149
Chicago/Turabian StyleFujimura, Atsushi, Seiji Yasui, Kazuyo Igawa, Ai Ueda, Kaori Watanabe, Tadashi Hanafusa, Yasuaki Ichikawa, Sachiko Yoshihashi, Kazuki Tsuchida, Atsunori Kamiya, and et al. 2020. "In Vitro Studies to Define the Cell-Surface and Intracellular Targets of Polyarginine-Conjugated Sodium Borocaptate as a Potential Delivery Agent for Boron Neutron Capture Therapy" Cells 9, no. 10: 2149. https://doi.org/10.3390/cells9102149