New Insights into the Liver–Visceral Adipose Axis During Hepatic Resection and Liver Transplantation

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
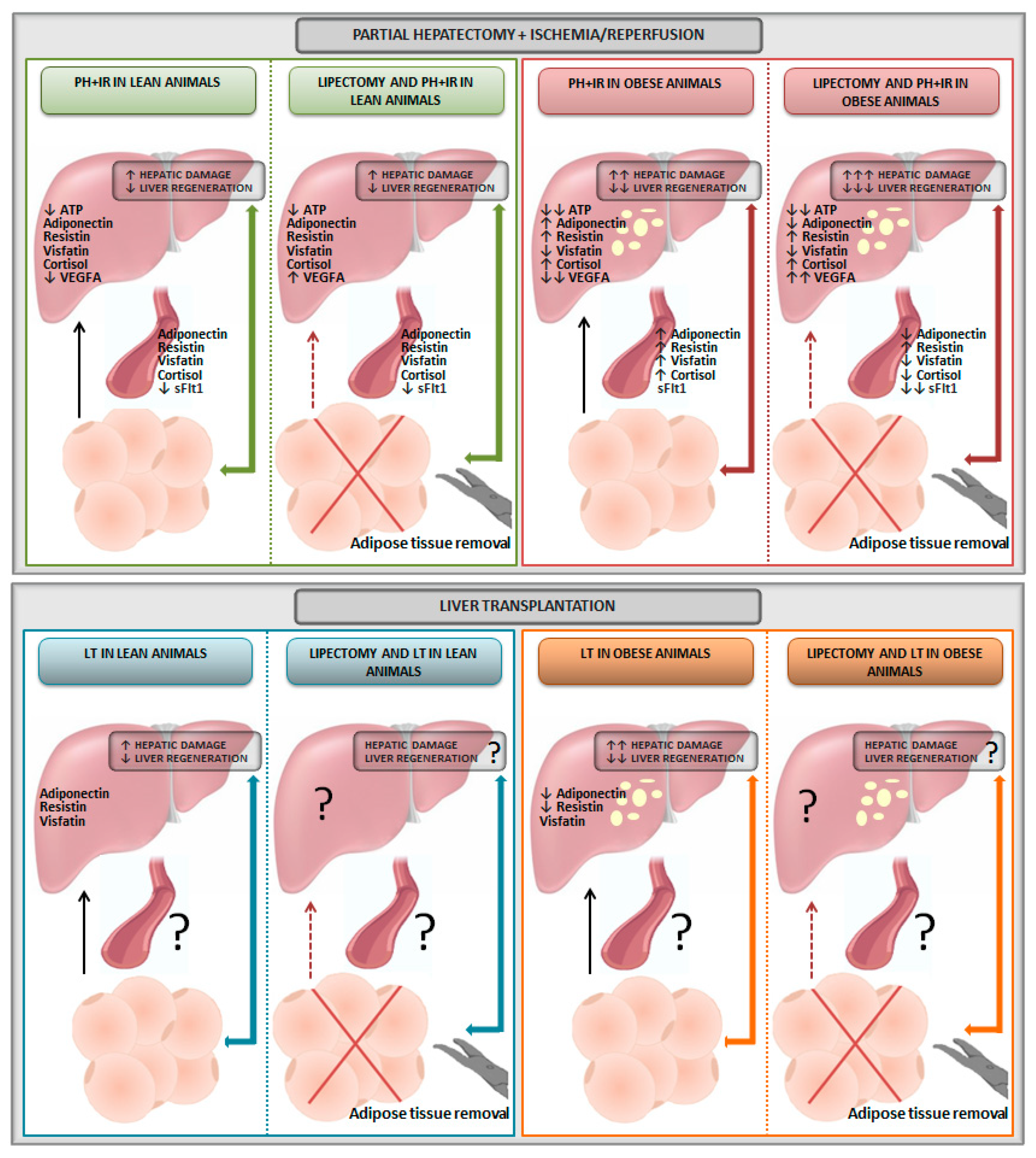
2. Relevance of Adipose Tissue in Experimental Models of Liver Resection
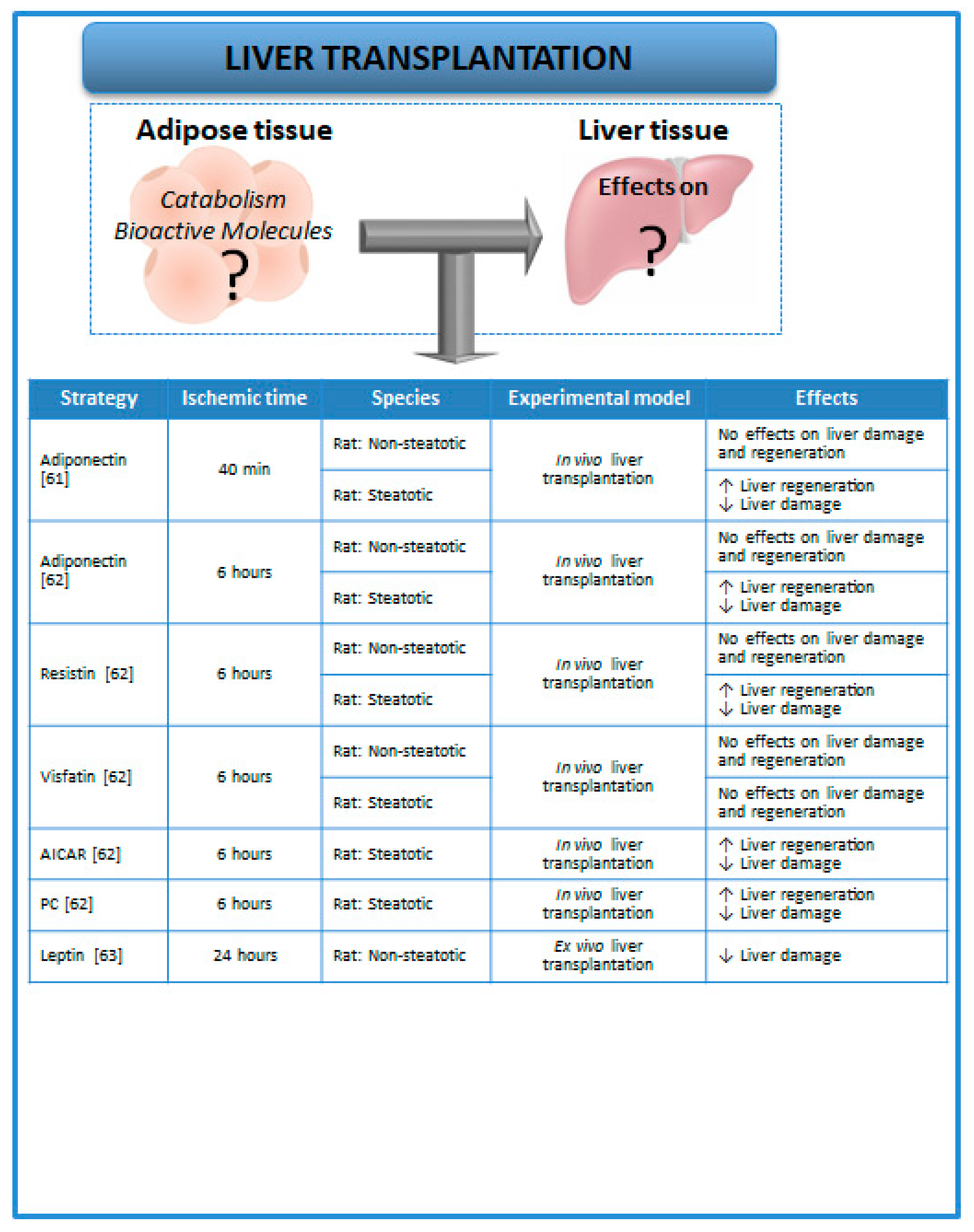
3. Relevance of Adipose Tissue in Experimental Models of Liver Transplantation
4. Relevance of Adipose Tissue in Patients Undergoing Liver Surgery
5. Experimental Strategies to Evaluate Adipose Tissue in Liver Surgery
5.1. Lipectomy
5.2. Transgenic Animal Models
6. Conclusions
Funding
Conflicts of Interest
Abbreviations
| AICAR | Cell-permeable adenosine analog that is a selective activator of AMPK |
| AMPK | Adenosine monophosphate-activated protein kinase |
| ATP | Adenosine triphosphate |
| BMI | Body mass index |
| BMP | Bone morphogenetic protein |
| CT | Computed tomography |
| Fas/CD95 | Apoptosis antigen 1 or cluster of differentiation 95 |
| FFA | Free fatty acids |
| HCC | Hepatocellular carcinoma |
| HGF | Hepatocyte growth factor |
| I/R | Ischemia–reperfusion |
| IL | Interleukin |
| JNK1 | c-Jun N-terminal kinase-1 |
| LDLT | Living donor liver transplantation |
| LT | Liver transplantation |
| M-CSF | Macrophage colony-stimulating factor |
| NO | Nitric oxide |
| PAI-1 | Plasminogen activator inhibitor-1 |
| PC | Preconditioning |
| PH | Partial hepatectomy |
| sFlt1 | Soluble form of the VEGF receptor 1 |
| SIP1 | Smad interacting protein 1 |
| TGF-β | Transforming growth factor-β |
| TNFα | Tumor necrosis factor α |
| UCP-1 | Mitochondrial uncoupling protein-1 |
| VEGFA | Vascular endothelial growth factor type A |
References
- Kershaw, E.E.; Flier, J.S. Adipose tissue as an endocrine organ. J. Clin. Endocrinol. Metab. 2004, 89, 2548–2556. [Google Scholar] [CrossRef] [PubMed]
- Frühbeck, G.; Gómez-Ambrosi, J.; Muruzábal, F.J.; Burrell, M.A. The adipocyte: A model for integration of endocrine and metabolic signaling in energy metabolism regulation. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E827–E847. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.G.; Dou, X.B.; Zhou, Z.X.; Song, Z.Y. Adipose tissue-liver axis in alcoholic liver disease. World J. Gastrointest. Pathophysiol. 2016, 7, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Parker, R. The role of adipose tissue in fatty liver diseases. Liver Res. 2018, 2, 35–42. [Google Scholar] [CrossRef]
- Large, V.; Peroni, O.; Letexier, D.; Ray, H.; Beylot, M. Metabolism of lipids in human white adipocyte. Diabetes Metab. 2004, 30, 294–309. [Google Scholar] [CrossRef]
- Nielsen, T.S.; Jessen, N.; Jørgensen, J.O.; Møller, N.; Lund, S. Dissecting adipose tissue lipolysis: Molecular regulation and implications for metabolic disease. J. Mol. Endocrinol. 2014, 52, R199–R222. [Google Scholar] [CrossRef] [PubMed]
- Blüher, M. Transgenic animal models for the study of adipose tissue biology. Best Pract. Res. Clin. Endocrinol. Metab. 2005, 19, 605–623. [Google Scholar] [CrossRef] [PubMed]
- Ye, D.W.; Rong, X.L.; Xu, A.M.; Guo, J. Liver-adipose tissue crosstalk: A key player in the pathogenesis of glucolipid metabolic disease. Chin. J. Integr. Med. 2017, 23, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Schäffler, A.; Schölmerich, J.; Büchler, C. Mechanisms of disease: Adipocytokines and visceral adipose tissue-emerging role in nonalcoholic fatty liver disease. Nat. Clin. Pract. Gastroenterol. Hepatol. 2005, 2, 273–280. [Google Scholar] [CrossRef]
- Casillas-Ramírez, A.; Mosbah, I.B.; Ramalho, F.; Roselló-Catafau, J.; Peralta, C. Past and future approaches to ischemia-reperfusion lesion associated with liver transplantation. Life Sci. 2006, 79, 1881–1894. [Google Scholar] [CrossRef]
- Serafin, A.; Rosello-Catafau, J.; Prats, N.; Xaus, C.; Gelpi, E.; Peralta, C. Ischemic preconditioning increases the tolerance of fatty liver to hepatic ischemia-reperfusion injury in the rat. Am. J. Pathol. 2002, 161, 587–601. [Google Scholar] [CrossRef]
- Clavien, P.; Harvey, P.; Strasberg, S. Preservation and reperfusion injuries in liver allografts. An overview and synthesis of current studies. Transplantation 1992, 53, 957–978. [Google Scholar] [CrossRef] [PubMed]
- Huguet, C.; Gavelli, A.; Chieco, P.; Bona, S.; Harb, J.; Joseph, J.M.; Jobard, J.; Gramaglia, M.; Lasserre, M. Liver ischemia for hepatic resection: Where is the limit? Surgery 1992, 111, 251–259. [Google Scholar] [PubMed]
- Fu, P.; Li, W. Nitric oxide in liver ischemia-reperfusion injury. In Liver Pathophysiology; Muriel, P., Ed.; Elsevier Inc.: London, UK, 2017; Volume 8, pp. 125–127. [Google Scholar]
- Cornide-Petronio, M.E.; Jiménez-Castro, M.B.; Gracia-Sancho, J.; Peralta, C. Ischemic preconditioning directly or remotely applied on the liver to reduce ischemia-reperfusion injury in resections and transplantation. In Liver Disease and Surgery; Tsoulfas, G., Ed.; IntechOpen: London, UK, 2019; in press. [Google Scholar]
- Gracia-Sancho, J.; Casillas-Ramírez, A.; Peralta, C. Molecular pathways in protecting the liver from ischaemia/reperfusion injury: A 2015 update. Clin. Sci. (Lond.) 2015, 129, 345–362. [Google Scholar] [CrossRef] [PubMed]
- Pratap, A.; Panakanti, R.; Yang, N.; Lakshmi, R.; Modanlou, K.A.; Eason, J.D.; Mahato, R.I. Cyclopamine attenuates acute warm ischemia reperfusion injury in cholestatic rat liver: Hope for marginal livers. Mol. Pharm. 2011, 8, 958–968. [Google Scholar] [CrossRef] [PubMed]
- Nadig, S.N.; Periyasamy, B.; Shafizadeh, S.F.; Polito, C.; Fiorini, R.N.; Rodwell, D.; Evans, Z.; Cheng, G.; Dunkelberger, D.; Schmidt, M.; et al. Hepatocellular ultrastructure after ischemia/reperfusion injury in human orthotopic liver transplantation. J. Gastrointest. Surg. 2004, 8, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Verran, D.; Kusyk, T.; Painter, D.; Fisher, J.; Koorey, D.; Strasser, S.; Stewart, G.; McCaughan, G. Clinical experience gained from the use of 120 steatotic donor livers for orthotopic liver transplantation. Liver Transplant. 2003, 9, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Schaubel, D.E.; Sima, C.S.; Goodrich, N.P.; Feng, S.; Merion, R.M. The survival benefit of deceased donor liver transplantation as a function of candidate disease severity and donor quality. Am. J. Transplant. 2008, 8, 419–425. [Google Scholar] [CrossRef]
- Ploeg, R.J.; D’Alessandro, A.M.; Knechtle, S.J.; Stegall, M.D.; Pirsch, J.D.; Hoffmann, R.M.; Sasaki, T.; Sollinger, H.W.; Belzer, F.O.; Kalayoglu, M. Risk factors for primary dysfunction after liver transplantation-a multivariate analysis. Transplantation 1993, 55, 807–813. [Google Scholar] [CrossRef]
- Behrns, K.E.; Tsiotos, G.G.; DeSouza, N.F.; Krishna, M.K.; Ludwig, J.; Nagorney, D.M. Hepatic steatosis as a potential risk factor for major hepatic resection. J. Gastrointest. Surg. 1998, 2, 292–298. [Google Scholar] [CrossRef]
- Safwan, M.; Collins, K.M.; Abouljoud, M.S.; Salgia, R. Outcome of liver transplantation in patients with prior bariatric surgery. Liver Transplant. 2017, 23, 1415–1421. [Google Scholar] [CrossRef] [PubMed]
- Shook, B.; Rivera Gonzalez, G.; Ebmeier, S.; Grisotti, G.; Zwick, R.; Horsley, V. The role of adipocytes in tissue regeneration and stem cell niches. Annu. Rev. Cell Dev. Biol. 2016, 32, 609–631. [Google Scholar] [CrossRef] [PubMed]
- Odegaard, J.I.; Chawla, A. Alternative macrophage activation and metabolism. Annu. Rev. Pathol. 2011, 6, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.; Muise, E.S.; Iyengar, P.; Wang, Z.V.; Chandalia, M.; Abate, N.; Zhang, B.B.; Bonaldo, P.; Chua, S.; Scherer, P.E. Metabolic dys-regulation and adipose tissue fibrosis: Role of collagen VI. Mol. Cell. Biol. 2009, 29, 1575–1591. [Google Scholar] [CrossRef] [PubMed]
- Hosogai, N.; Fukuhara, A.; Oshima, K.; Miyata, Y.; Tanaka, S.; Segawa, K.; Furukawa, S.; Tochino, Y.; Komuro, R.; Matsuda, M.; et al. Adipose tissue hypoxia in obesity and its impact on adipocytokine dysregulation. Diabetes 2007, 56, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Elias-Miro, M.; Massip-Salcedo, M.; Jimenez-Castro, M.; Peralta, C. Does adiponectin benefit steatotic liver transplantation? Liver Transplant. 2011, 17, 993–1004. [Google Scholar] [CrossRef]
- Tilg, H.; Hotamisligil, G.S. Nonalcoholic fatty liver disease: Cytokine-adipokine interplay and regulation of insulin resistance. Gastroenterology 2006, 131, 934–945. [Google Scholar] [CrossRef]
- Farrell, G.C. Probing Prometheus: Fat fueling the fire? Hepatology 2004, 40, 1252–1255. [Google Scholar] [CrossRef]
- Gordon, S. Macrophage heterogeneity and tissue lipids. J. Clin. Investig. 2007, 117, 89–93. [Google Scholar] [CrossRef]
- Wellen, K.E.; Hotamisligil, G.S. Inflammation, stress, and diabetes. J. Clin. Investig. 2005, 115, 1111–1119. [Google Scholar] [CrossRef] [Green Version]
- Wellen, K.E.; Hotamisligil, G.S. Obesity-induced inflammatory changes in adipose tissue. J. Clin. Investig. 2003, 112, 1785–1788. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; Mccann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Duwaerts, C.C.; Maher, J.J. Macronutrients and the Adipose-Liver Axis in Obesity and Fatty Liver. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 749–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harman-Boehm, I.; Bluher, M.; Redel, H.; Sion-Vardy, N.; Ovadia, S.; Avinoach, E.; Shai, I.; Klöting, N.; Stumvoll, M.; Bashan, N.; et al. Macrophage infiltration into omental versus subcutaneous fat across different populations: Effect of regional adiposity and the comorbidities of obesity. J. Clin. Endocrinol. Metab. 2007, 92, 2240–2247. [Google Scholar] [CrossRef] [PubMed]
- Nov, O.; Shapiro, H.; Ovadia, H.; Tarnovscki, T.; Dvir, I.; Shemesh, E.; Kovsan, J.; Shelef, I.; Carmi, Y.; Voronov, E.; et al. Interleukin-1β regulates fat-liver crosstalk in obesity by auto-paracrine modulation of adipose tissue inflammation and expandability. PLoS ONE 2013, 8, e53626. [Google Scholar] [CrossRef] [PubMed]
- Bergman, R.N.; Kim, S.P.; Hsu, I.R.; Catalano, K.J.; Chiu, J.D.; Kabir, M.; Richey, J.M.; Ader, M. Abdominal obesity: Role in the pathophysiology of metabolic disease and cardiovascular risk. Am. J. Med. 2007, 120, S3–S8; discussion S29–S32. [Google Scholar] [CrossRef] [PubMed]
- Kabir, M.; Catalano, K.J.; Ananthnarayan, S.; Kim, S.P.; Van Citters, G.W.; Dea, M.K.; Bergman, R.N. Molecular evidence supporting the portal theory: A causative link between visceral adiposity and hepatic insulin resistance. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E454–E461. [Google Scholar] [CrossRef]
- Sabio, G.; Das, M.; Mora, A.; Zhang, Z.; Jun, J.Y.; Ko, H.J.; Barrett, T.; Kim, J.K.; Davis, R.J. A stress signalling pathway in adipose tissue regulates hepatic insulin resistance. Science 2008, 322, 1539–1543. [Google Scholar] [CrossRef] [PubMed]
- Wueest, S.; Rapold, R.A.; Schumann, D.M.; Rytka, J.M.; Schildknecht, A.; Nov, O.; Chervonsky, A.V.; Rudich, A.; Schoenle, E.J.; Donath, M.Y.; et al. Deletion of Fas in adipocytes relieves adipose tissue inflammation and hepatic manifestations of obesity in mice. J. Clin. Investig. 2010, 120, 191–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caruana, J.A.; Whalen, D.A., Jr.; Anthony, W.P.; Sunby, C.R.; Ciechoski, M.P. Paradoxical effects of glucose feeding on liver regeneration and survival after partial hepatectomy. Endocr. Res. 1986, 12, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, T.; Ozawa, K.; Asano, M.; Ukikusa, M.; Kamiyama, Y.; Tobe, T. Differences in predominant energy substrate in relation to the resected hepatic mass in the phase immediately after hepatectomy. J. Lab. Clin. Med. 1981, 97, 887–898. [Google Scholar] [PubMed]
- Nakatani, T.; Yasuda, K.; Ozawa, K.; Kawashima, S.; Tobe, T. Effects of (+)-octanoylcarnitine on deoxyribonucleic acid synthesis in regenerating rabbit liver. Clin. Sci. (Lond.) 1982, 62, 295–297. [Google Scholar] [CrossRef] [PubMed]
- Gazit, V.; Weymann, A.; Hartman, E.; Finck, B.N.; Hruz, P.W.; Tzekov, A.; Rudnick, D.A. Liver regeneration is impaired in lipodystrophic fatty liver dystrophy mice. Hepatology 2010, 52, 2109–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tijburg, L.B.; Nyathi, C.B.; Meijer, G.W.; Geelen, M.J. Biosynthesis and secretion of triacylglycerol in rat liver after partial hepatectomy. Biochem. J. 1991, 277, 723–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brasaemle, D.L. Cell biology. A metabolic push to proliferate. Science 2006, 313, 1581–1582. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.; Garfinkel, A.S.; Schotz, M.C. Hepatic triglyceride secretion in relation to lipogenesis and free fatty acid mobilization in fasted and glucose-refed rats. J. Lipid Res. 1968, 9, 1–7. [Google Scholar] [PubMed]
- Michalopoulos, G.K.; DeFrances, M.C. Liver regeneration. Science 1997, 276, 60–66. [Google Scholar] [CrossRef]
- Palmero, E.; Ricart, D.; Llobera, M.; Peinado-Onsurbe, J. Partial hepatectomy and/or surgical stress provoke changes in the expression of lipoprotein lipase and actin in liver and extrahepatic tissues. Biochim. Biophys. Acta 1999, 1441, 61–68. [Google Scholar] [CrossRef]
- Sabugal, R.; Robert, M.Q.; Julve, J.; Auwerx, J.; Llobera, M.; Peinado-Onsurbe, J. Hepatic regeneration induces changes in lipoprotein lipase activity in several tissues and its re-expression in the liver. Biochem. J. 1996, 318, 597–602. [Google Scholar] [CrossRef] [Green Version]
- Shteyer, E.; Liao, Y.; Muglia, L.J.; Hruz, P.W.; Rudnick, D.A. Disruption of hepaticadipogenesis is associated with impaired liver regeneration in mice. Hepatology 2004, 40, 1322–1332. [Google Scholar] [CrossRef]
- Kohjima, M.; Tsai, T.H.; Tackett, B.C.; Thevananther, S.; Li, L.; Chang, B.H.; Chan, L. Delayed liver regeneration after partial hepatectomy in adipose differentiation related protein-null mice. J. Hepatol. 2013, 59, 1246–1254. [Google Scholar] [CrossRef] [Green Version]
- El-Badry, A.M.; Moritz, W.; Contaldo, C.; Tian, Y.; Graf, R.; Clavien, P.A. Prevention of reperfusion injury and microcirculatory failure in macrosteatotic mouse liver by omega-3 fatty acids. Hepatology 2007, 45, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Marsman, H.A.; de Graaf, W.; Heger, M.; van Golen, R.F.; Ten Kate, F.J.; Bennink, R.; van Gulik, T.M. Hepatic regeneration and functional recovery following partial liver resection in an experimental model of hepatic steatosis treated with omega-3 fatty acids. Br. J. Surg. 2013, 100, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, Y.; Shi, Y.; Bu, H.; Ye, F. Deletion of SIP1 promotes liver regeneration and lipid accumulation. Pathol. Res. Pract. 2016, 212, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Mendes-Braz, M.; Elias-Miró, M.; Kleuser, B.; Fayyaz, S.; Jiménez-Castro, M.B.; Massip-Salcedo, M.; Gracia-Sancho, J.; Ramalho, F.S.; Rodes, J.; Peralta, C. The effects of glucose and lipids in steatotic and nonsteatotic livers in conditions of partial hepatectomy under ischaemia-reperfusion. Liver Int. 2014, 34, e271–e289. [Google Scholar] [CrossRef] [PubMed]
- Elias-Miró, M.; Mendes-Braz, M.; Cereijo, R.; Villarroya, F.; Jiménez-Castro, M.B.; Gracia-Sancho, J.; Guixé-Muntet, S.; Massip-Salcedo, M.; Domingo, J.C.; Bermudo, R.; et al. Resistin and visfatin in steatotic and nonsteatotic livers in the setting of partial hepatectomy under ischemia-reperfusion. J. Hepatol. 2014, 60, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Cornide-Petronio, M.E.; Bujaldon, E.; Mendes-Braz, M.; Avalos de León, C.G.; Jiménez-Castro, M.B.; Álvarez-Mercado, A.I.; Gracia-Sancho, J.; Rodés, J.; Peralta, C. The impact of cortisol in steatotic and nonsteatotic liver surgery. J. Cell. Mol. Med. 2017, 21, 2344–2358. [Google Scholar] [CrossRef] [PubMed]
- Bujaldon, E.; Cornide-Petronio, M.E.; Gulfo, J.; Rotondo, F.; Ávalos de León, C.; Negrete-Sánchez, E.; Gracia-Sancho, J.; Novials, A.; Jiménez-Castro, M.B.; Peralta Uroz, C. Relevance of VEGFA in rat livers subjected to partial hepatectomy under ischemia-reperfusion. J. Mol. Med. (Berl.) 2019, 97, 1299–1314. [Google Scholar] [CrossRef] [Green Version]
- Man, K.; Zhao, Y.; Xu, A.; Lo, C.M.; Lam, K.S.; Ng, K.T.; Ho, J.W.; Sun, C.K.; Lee, T.K.; Li, X.L.; et al. Fat-derived hormone adiponectin combined with FTY720 significantly improves small-for-size fatty liver graft survival. Am. J. Transplant. 2006, 6, 467–476. [Google Scholar] [CrossRef]
- Jiménez-Castro, M.B.; Casillas-Ramírez, A.; Mendes-Braz, M.; Massip-Salcedo, M.; Gracia-Sancho, J.; Elias-Miró, M.; Rodés, J.; Peralta, C. Adiponectin and resistin protect steatotic livers undergoing transplantation. J. Hepatol. 2013, 59, 1208–1214. [Google Scholar] [CrossRef]
- Carbone, M.; Campagnolo, L.; Angelico, M.; Tisone, G.; Almerighi, C.; Telesca, C.; Lenci, I.; Moscatelli, I.; Massoud, R.; Baiocchi, L. Leptin attenuates ischemia-reperfusion injury in the rat liver. Transpl. Int. 2012, 25, 1282–1288. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Castro, M.B.; Meroño, N.; Mendes-Braz, M.; Gracia-Sancho, J.; Martínez-Carreres, L.; Cornide-Petronio, M.E.; Casillas-Ramirez, A.; Rodés, J.; Peralta, C. The effect of brain death in rat steatotic and nonsteatotic liver transplantation with previous ischemic preconditioning. J. Hepatol. 2015, 62, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Castro, M.B.; Gracia-Sancho, J.; Peralta, C. Brain death and marginal grafts in liver transplantation. Cell Death Dis. 2015, 6, e1777. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wang, Z.Y.; Zhu, L.L.; Wu, F.; Chen, D.Q.; Sun, L.F.; Lu, Z.Q. Resistin expression in adipose tissues and its effect on glucose metabolism in rats with brain injury. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Marra, F.; Bertolani, C. Adipokines in liver diseases. Hepatology 2009, 50, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Mullen, J.T.; Moorman, D.W.; Davenport, D.L. The obesity paradox: Body mass index and outcomes in patients undergoing nonbariatric general surgery. Ann. Surg. 2009, 250, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Dindo, D.; Muller, M.K.; Weber, M.; Clavien, P.A. Obesity in general elective surgery. Lancet 2003, 361, 2032–2035. [Google Scholar] [CrossRef]
- Barone, M.; Viggiani, M.T.; Avolio, A.W.; Iannone, A.; Rendina, M.; Di Leo, A. Obesity as predictor of postoperative outcomes in liver transplant candidates: Review of the literature and future perspectives. Dig. Liver Dis. 2017, 49, 957–966. [Google Scholar] [CrossRef]
- Hamaguchi, Y.; Kaido, T.; Okumura, S.; Kobayashi, A.; Shirai, H.; Yao, S.; Yagi, S.; Kamo, N.; Seo, S.; Taura, K.; et al. Preoperative Visceral Adiposity and Muscularity Predict Poor Outcomes after Hepatectomy for Hepatocellular Carcinoma. Liver Cancer 2019, 8, 92–109. [Google Scholar] [CrossRef]
- Calle, E.E.; Rodriguez, C.; Walker-Thurmond, K.; Thun, M.J. Overweight, obesity, and mortality from cancer in a prospectively studied cohort of US adults. N. Engl. J. Med. 2003, 348, 1625–1638. [Google Scholar] [CrossRef]
- Renehan, A.G.; Tyson, M.; Egger, M.; Heller, R.F.; Zwahlen, M. Body-mass index and incidence of cancer: A systematic review and meta-analysis of prospective observational studies. Lancet 2008, 371, 569–578. [Google Scholar] [CrossRef]
- Arnold, M.; Pandeya, N.; Byrnes, G.; Renehan, A.G.; Stevens, G.A.; Ezzati, M.; Ferlay, J.; Miranda, J.J.; Romieu, I.; Dikshit, R.; et al. Global burden of cancer attributable to high body-mass index in 2012: A population-based study. Lancet Oncol. 2015, 16, 36–46. [Google Scholar] [CrossRef]
- Kaibori, M.; Ishizaki, M.; Iida, H.; Matsui, K.; Sakaguchi, T.; Inoue, K.; Mizuta, T.; Ide, Y.; Iwasaka, J.; Kimura, Y.; et al. Effect of Intramuscular Adipose Tissue Content on Prognosis in Patients Undergoing Hepatocellular Carcinoma Resection. J. Gastrointest. Surg. 2015, 19, 1315–1323. [Google Scholar] [CrossRef] [PubMed]
- Kamo, N.; Kaido, T.; Hamaguchi, Y.; Okumura, S.; Kobayashi, A.; Shirai, H.; Yao, S.; Yagi, S.; Uemoto, S. Impact of sarcopenic obesity on outcomes in patients undergoing living donor liver transplantation. Clin. Nutr. 2018. [Google Scholar] [CrossRef] [PubMed]
- Morris, K.; Tuorto, S.; Gönen, M.; Schwartz, L.; DeMatteo, R.; D'Angelica, M.; Jarnagin, W.R.; Fong, Y. Simple measurement of intra-abdominal fat for abdominal surgery outcome prediction. Arch. Surg. 2010, 145, 1069–1073. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.; Kaido, T.; Hamaguchi, Y.; Okumura, S.; Shirai, H.; Kamo, N.; Yagi, S.; Taura, K.; Okajima, H.; Uemoto, S. Impact of Visceral Adiposity as Well as Sarcopenic Factors on Outcomes in Patients Undergoing Liver Resection for Colorectal Liver Metastases. World J. Surg. 2018, 42, 1180–1191. [Google Scholar] [CrossRef] [PubMed]
- Hakimi, A.A.; Furberg, H.; Zabor, E.C.; Jacobsen, A.; Schultz, N.; Ciriello, G.; Mikklineni, N.; Fiegoli, B.; Kim, P.H.; Voss, M.H.; et al. An epidemiologic and genomic investigation into the obesity paradox in renal cell carcinoma. J. Natl. Cancer Inst. 2013, 105, 1862–1870. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; Birdsell, L.; Macdonald, N.; Reiman, T.; Clandinin, M.T.; McCargar, L.J.; Murphy, R.; Ghosh, S.; Sawyer, M.B.; Baracos, V.E. Cancer cachexia in the age of obesity: Skeletal muscle depletion is a powerful prognostic factor, independent of body mass index. J. Clin. Oncol. 2013, 31, 1539–1547. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, S.; Siegert, S.; Koch, M.; Walter, J.; Heits, N.; Hinz, S.; Jacobs, G.; Hampe, J.; Schafmayer, C.; Nöthlings, U. Postdiagnosis body mass index and risk of mortality in colorectal cancer survivors: A prospective study and metaanalysis. Cancer Causes Control 2014, 25, 1407–1418. [Google Scholar] [CrossRef] [PubMed]
- Amptoulach, S.; Gross, G.; Kalaitzakis, E. Differential impact of obesity and diabetes mellitus on survival after liver resection for colorectal cancer metastases. J. Surg. Res. 2015, 199, 378–385. [Google Scholar] [CrossRef]
- Matsumoto, K.; Miyake, Y.; Umeda, Y.; Matsushita, H.; Matsuda, H.; Takaki, A.; Sadamori, H.; Nouso, K.; Yagi, T.; Fujiwara, T.; et al. Serial changes of serum growth factor levels and liver regeneration after partial hepatectomy in healthy humans. Int. J. Mol. Sci. 2013, 14, 20877–20889. [Google Scholar] [CrossRef] [PubMed]
- Veldt, B.J.; Poterucha, J.J.; Watt, K.D.; Wiesner, R.H.; Hay, J.E.; Rosen, C.B.; Heimbach, J.K.; Janssen, H.L.; Charlton, M.R. Insulin resistance, serum adipokines and risk of fibrosis progression in patients transplanted for hepatitis C. Am. J. Transplant. 2009, 9, 1406–1413. [Google Scholar] [CrossRef] [PubMed]
- Short, M.K.; Clouthier, D.E.; Schaefer, I.M.; Hammer, R.E.; Magnuson, M.A.; Beale, E.G. Tissue-specific, developmental, hormonal, and dietary regulation of rat phosphoenolpyruvate carboxykinase-human growth hormone fusion genes in transgenic mice. Mol. Cell. Biol. 1992, 12, 1007–1020. [Google Scholar] [CrossRef] [PubMed]
- Cassard-Doulcier, A.M.; Gelly, C.; Fox, N.; Schrementi, J.; Raimbault, S.; Klaus, S.; Forest, C.; Bouillaud, F.; Ricquier, D. Tissue-specific and beta-adrenergic regulation of the mitochondrial uncoupling protein gene: Control by cis-acting elements in the 50-flanking region. Mol. Endocrinol. 1993, 7, 497–506. [Google Scholar] [PubMed]
- Valet, P.; Tavernier, G.; Castan-Laurell, I.; Saulnier-Blache, J.S.; Langin, D. Understanding adipose tissue development from transgenic animal models. J. Lipid Res. 2002, 43, 835–860. [Google Scholar] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cornide-Petronio, M.E.; Jiménez-Castro, M.B.; Gracia-Sancho, J.; Peralta, C. New Insights into the Liver–Visceral Adipose Axis During Hepatic Resection and Liver Transplantation. Cells 2019, 8, 1100. https://doi.org/10.3390/cells8091100
Cornide-Petronio ME, Jiménez-Castro MB, Gracia-Sancho J, Peralta C. New Insights into the Liver–Visceral Adipose Axis During Hepatic Resection and Liver Transplantation. Cells. 2019; 8(9):1100. https://doi.org/10.3390/cells8091100
Chicago/Turabian StyleCornide-Petronio, María Eugenia, Mónica B. Jiménez-Castro, Jordi Gracia-Sancho, and Carmen Peralta. 2019. "New Insights into the Liver–Visceral Adipose Axis During Hepatic Resection and Liver Transplantation" Cells 8, no. 9: 1100. https://doi.org/10.3390/cells8091100