
Vessel-on-a-Chip: A Powerful Tool for Investigating Endothelial COVID-19 Fingerprints
 ,
,  , , and
, , and 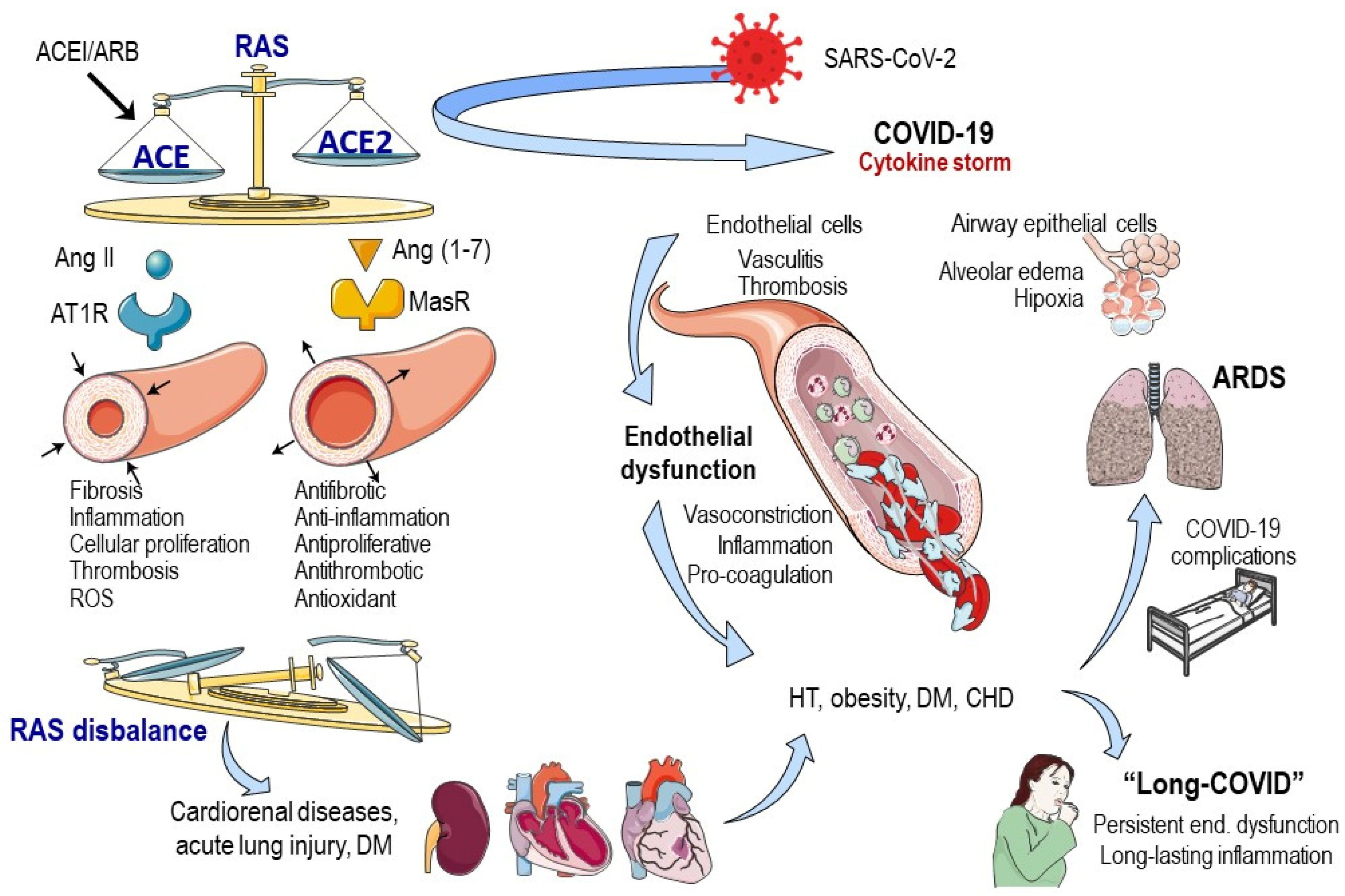{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Vascular Fingerprint of COVID-19
2.1. SARS-CoV-2 Gate
2.2. ACE2 and COVID-19 Severity and Medical Interventions
2.3. Endothelial Dysfunction and COVID-19
2.4. Long COVID-19
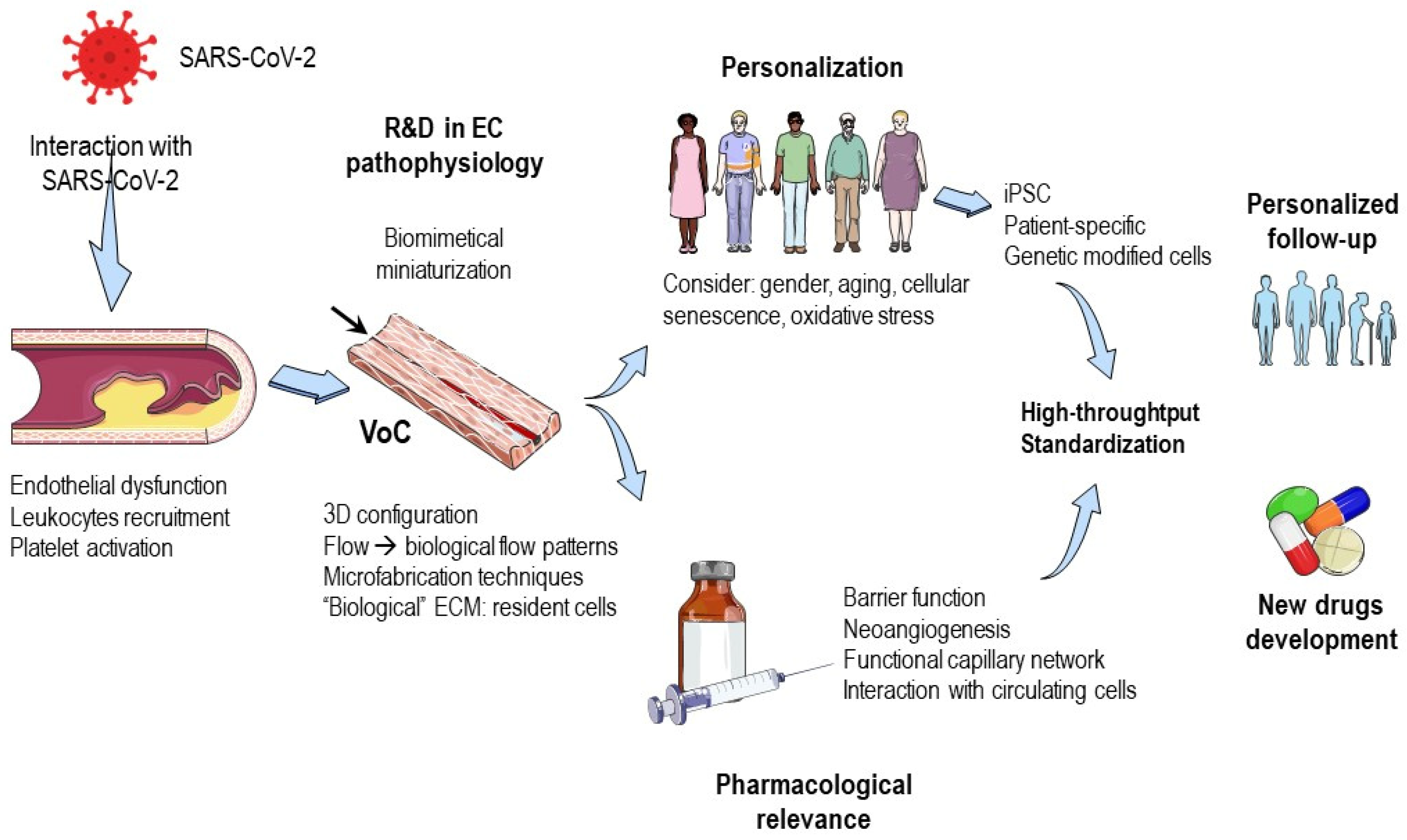
3. Contribution of Microfluidic Technology
- Determining the main effects of SARS-CoV-2 on endothelial function, including activation of endothelial cells, recruitment of leukocytes, and platelet activation. Factors such as cellular senescence, oxidative stress, and other aging-related features should also be taken into consideration, along with gender influence. Therefore, microfluidic chips should be personalized to address these concerns.
- Exploring the effects of common cardiovascular drugs on endothelial responses to SARS-CoV-2.
- Contributions of miniaturized biomimetic endothelium in vitro models.
- Pharmacological relevance of endothelial functions in microfluidic chips.
- Advantages of vessel-on-a-chip systems with respect to 2D classical in vitro models.
- “Personalization” of endothelial models.
- Tools for “point-of-care” solutions, diagnosis, or follow-up.
3.1. Contributions of Miniaturized Biomimetic Endothelium In Vitro Models
3.2. Pharmacological Relevance of Endothelial Functions in Microfluidic Chips
3.2.1. Barrier Function
3.2.2. Angiogenesis
3.2.3. Capillary Perfusable Network
3.2.4. Interaction with Circulating Cells
3.3. Advantages of Vessel-on-a-Chip Systems with Respect to 2D Classical In Vitro Models
3.4. “Personalization” of Endothelial Models
3.5. Tools for “Point-of-Care” Solutions, Diagnosis or Follow-Up
4. Concluding Remarks
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hyams, C.; Challen, R.; Marlow, R.; Nguyen, J.; Begier, E.; Southern, J.; King, J.; Morley, A.; Kinney, J.; Clout, M.; et al. Severity of Omicron (B.1.1.529) and Delta (B.1.617.2) SARS-CoV-2 Infection among Hospitalised Adults: A Prospective Cohort Study in Bristol, United Kingdom. Lancet Reg. Health Eur. 2023, 25, 100556. [Google Scholar] [CrossRef]
- Parums, D.V. Editorial: The XBB.1.5 (‘Kraken’) Subvariant of Omicron SARS-CoV-2 and Its Rapid Global Spread. Med. Sci. Monit. 2023, 29, e939580. [Google Scholar] [CrossRef] [PubMed]
- Dessie, Z.G.; Zewotir, T. Mortality-Related Risk Factors of COVID-19: A Systematic Review and Meta-Analysis of 42 Studies and 423,117 Patients. BMC Infect. Dis. 2021, 21, 855. [Google Scholar] [CrossRef] [PubMed]
- Davis, H.E.; McCorkell, L.; Vogel, J.M.; Topol, E.J. Long COVID: Major Findings, Mechanisms and Recommendations. Nat. Rev. Microbiol. 2023, 21, 133–146. [Google Scholar] [CrossRef]
- Mitrani, R.D.; Dabas, N.; Goldberger, J.J. COVID-19 Cardiac Injury: Implications for Long-Term Surveillance and Outcomes in Survivors. Heart Rhythm 2020, 17, 1984–1990. [Google Scholar] [CrossRef] [PubMed]
- Tran, V.T.; Porcher, R.; Pane, I.; Ravaud, P. Course of Post COVID-19 Disease Symptoms over Time in the ComPaRe Long COVID Prospective e-Cohort. Nat. Commun. 2022, 13, 1812. [Google Scholar] [CrossRef] [PubMed]
- Greenhalgh, T.; Knight, M.; A’Court, C.; Buxton, M.; Husain, L. Management of Post-Acute COVID-19 in Primary Care. BMJ 2020, 370, m3026. [Google Scholar] [CrossRef]
- Zhou, Y.; Chi, J.; Lv, W.; Wang, Y. Obesity and Diabetes as High-Risk Factors for Severe Coronavirus Disease 2019 (COVID-19). Diabetes Metab. Res. Rev. 2020, 37, e3377. [Google Scholar] [CrossRef]
- Evans, P.C.; Rainger, G.E.; Mason, J.C.; Guzik, T.J.; Osto, E.; Stamataki, Z.; Neil, D.; Hoefer, I.E.; Fragiadaki, M.; Waltenberger, J.; et al. Endothelial Dysfunction in COVID-19: A Position Paper of the ESC Working Group for Atherosclerosis and Vascular Biology, and the ESC Council of Basic Cardiovascular Science. Cardiovasc. Res. 2020, 116, 2177–2184. [Google Scholar] [CrossRef] [PubMed]
- Eassa, H.A.; Helal, N.A.; Amer, A.M.; Fouad, A.; Bedair, A.F.; Nagib, R.; Mansoor, I.; Hawash, M.; Abdul-Latif, M.; Mohammed, K.H.A.; et al. 3D-Printed Microfluidics Potential in Combating Future and Current Pandemics (COVID-19). Recent Adv. Drug Deliv. Formul. 2022, 16, 192–216. [Google Scholar] [CrossRef]
- Shahabipour, F.; Satta, S.; Mahmoodi, M.; Sun, A.; de Barros, N.R.; Li, S.; Hsiai, T.; Ashammakhi, N. Engineering Organ-on-a-Chip Systems to Model Viral Infections. Biofabrication 2023, 15, 022001. [Google Scholar] [CrossRef] [PubMed]
- Kirtipal, N.; Bharadwaj, S.; Kang, S.G. From SARS to SARS-CoV-2, Insights on Structure, Pathogenicity and Immunity Aspects of Pandemic Human Coronaviruses. Infect. Genet. Evol. 2020, 85, 104502. [Google Scholar] [CrossRef] [PubMed]
- Cascella, M.; Rajnik, M.; Aleem, A.; Dulebohn, S.C.; Di Napoli, R. Features, Evaluation, and Treatment of Coronavirus (COVID-19); [Updated 9 January 2023], In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK554776/ (accessed on 19 March 2023).
- Abdelrahman, Z.; Li, M.; Wang, X. Comparative Review of SARS-CoV-2, SARS-CoV, MERS-CoV, and Influenza A Respiratory Viruses. Front. Immunol. 2020, 11, 552909. [Google Scholar] [CrossRef] [PubMed]
- Hasoksuz, M.; Kilic, S.; Sarac, F. Coronaviruses and SARS-CoV-2. Turk. J. Med. Sci. 2020, 50, 549–556. [Google Scholar] [CrossRef]
- Sharma, A.; Ahmad Farouk, I.; Lal, S.K. COVID-19: A Review on the Novel Coronavirus Disease Evolution, Transmission, Detection, Control and Prevention. Viruses 2021, 13, 202. [Google Scholar] [CrossRef]
- Li, J.; Gao, J.; Xu, Y.P.; Zhou, T.L.; Jin, Y.Y.; Lou, J.N. [Expression of severe acute respiratory syndrome coronavirus receptors, ACE2 and CD209L in different organ derived microvascular endothelial cells]. Zhonghua Yi Xue Za Zhi 2007, 87, 833–837. [Google Scholar]
- Hamming, I.; Timens, W.; Bulthuis, M.L.; Lely, A.T.; Navis, G.; van Goor, H. Tissue Distribution of ACE2 Protein, the Functional Receptor for SARS Coronavirus. A First Step in Understanding SARS Pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Wang, M.; Hao, H.; Leeper, N.J.; Zhu, L. Thrombotic Regulation From the Endothelial Cell Perspectives. Arterioscler. Thromb. Vasc. Biol. 2018, 38, e90–e95. [Google Scholar] [CrossRef]
- Sarzi-Puttini, P.; Giorgi, V.; Sirotti, S.; Marotto, D.; Ardizzone, S.; Rizzardini, G.; Antinori, S.; Galli, M. COVID-19, Cytokines and Immunosuppression: What Can We Learn from Severe Acute Respiratory Syndrome? Clin. Exp. Rheumatol. 2020, 38, 337–342. [Google Scholar] [CrossRef]
- Amraei, R.; Rahimi, N. COVID-19, Renin-Angiotensin System and Endothelial Dysfunction. Cells 2020, 9, 1652. [Google Scholar] [CrossRef]
- Wong, W.T.; Tian, X.Y.; Huang, Y. Endothelial Dysfunction in Diabetes and Hypertension: Cross Talk in RAS, BMP4, and ROS-Dependent COX-2-Derived Prostanoids. J. Cardiovasc. Pharmacol. 2013, 61, 204–214. [Google Scholar] [CrossRef]
- Patel, V.B.; Zhong, J.C.; Grant, M.B.; Oudit, G.Y. Role of the ACE2/Angiotensin 1-7 Axis of the Renin-Angiotensin System in Heart Failure. Circ. Res. 2016, 118, 1313–1326. [Google Scholar] [CrossRef] [PubMed]
- South, A.M.; Shaltout, H.A.; Washburn, L.K.; Hendricks, A.S.; Diz, D.I.; Chappell, M.C. Fetal Programming and the Angiotensin-(1-7) Axis: A Review of the Experimental and Clinical Data. Clin. Sci. 2019, 133, 55–74. [Google Scholar] [CrossRef]
- Long, B.; Brady, W.J.; Koyfman, A.; Gottlieb, M. Cardiovascular Complications in COVID-19. Am. J. Emerg. Med. 2020, 38, 1504–1507. [Google Scholar] [CrossRef]
- Guzik, T.J.; Mohiddin, S.A.; Dimarco, A.; Patel, V.; Savvatis, K.; Marelli-Berg, F.M.; Madhur, M.S.; Tomaszewski, M.; Maffia, P.; D’Acquisto, F.; et al. COVID-19 and the Cardiovascular System: Implications for Risk Assessment, Diagnosis, and Treatment Options. Cardiovasc. Res. 2020, 116, 1666–1687. [Google Scholar] [CrossRef] [PubMed]
- Kuba, K.; Imai, Y.; Rao, S.; Gao, H.; Guo, F.; Guan, B.; Huan, Y.; Yang, P.; Zhang, Y.; Deng, W.; et al. A Crucial Role of Angiotensin Converting Enzyme 2 (ACE2) in SARS Coronavirus-Induced Lung Injury. Nat. Med. 2005, 11, 875–879. [Google Scholar] [CrossRef]
- Del Turco, S.; Vianello, A.; Ragusa, R.; Caselli, C.; Basta, G. COVID-19 and Cardiovascular Consequences: Is the Endothelial Dysfunction the Hardest Challenge? Thromb. Res. 2020, 196, 143–151. [Google Scholar] [CrossRef]
- Flammer, A.J.; Anderson, T.; Celermajer, D.S.; Creager, M.A.; Deanfield, J.; Ganz, P.; Hamburg, N.M.; Luscher, T.F.; Shechter, M.; Taddei, S.; et al. The Assessment of Endothelial Function: From Research into Clinical Practice. Circulation 2012, 126, 753–767. [Google Scholar] [CrossRef]
- Forstermann, U.; Sessa, W.C. Nitric Oxide Synthases: Regulation and Function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Colantuoni, A.; Martini, R.; Caprari, P.; Ballestri, M.; Capecchi, P.L.; Gnasso, A.; Lo Presti, R.; Marcoccia, A.; Rossi, M.; Caimi, G. COVID-19 Sepsis and Microcirculation Dysfunction. Front. Physiol. 2020, 11, 747. [Google Scholar] [CrossRef]
- Chen, F.; Chen, Y.; Ke, Q.; Wang, Y.; Gong, Z.; Chen, X.; Cai, Y.; Li, S.; Sun, Y.; Peng, X.; et al. ApoE4 Associated with Severe COVID-19 Outcomes via Downregulation of ACE2 and Imbalanced RAS Pathway. J. Transl. Med. 2023, 21, 103. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 Entry into Cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Martel, J.; Ko, Y.F.; Young, J.D.; Ojcius, D.M. Could Nasal Nitric Oxide Help to Mitigate the Severity of COVID-19? Microbes Infect. 2020, 22, 168–171. [Google Scholar] [CrossRef] [PubMed]
- Wadman, M. Why Obesity Worsens COVID-19. Science 2020, 369, 1280–1281. [Google Scholar] [CrossRef] [PubMed]
- Connors, J.M.; Levy, J.H. COVID-19 and Its Implications for Thrombosis and Anticoagulation. Blood 2020, 135, 2033–2040. [Google Scholar] [CrossRef] [PubMed]
- Green, S.J. COVID-19 Accelerates Endothelial Dysfunction and Nitric Oxide Deficiency. Microbes Infect. 2020, 22, 149–150. [Google Scholar] [CrossRef]
- Iba, T.; Levy, J.H.; Levi, M.; Thachil, J. Coagulopathy in COVID-19. J. Thromb. Haemost. 2020, 18, 2103–2109. [Google Scholar] [CrossRef]
- Rouaud, F.; Mean, I.; Citi, S. The ACE2 Receptor for Coronavirus Entry Is Localized at Apical Cell-Cell Junctions of Epithelial Cells. Cells 2022, 11, 627. [Google Scholar] [CrossRef]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and Immunological Features of Severe and Moderate Coronavirus Disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef]
- Rojas-Garcia, M.; Vazquez, B.; Torres-Poveda, K.; Madrid-Marina, V. Lethality Risk Markers by Sex and Age-Group for COVID-19 in Mexico: A Cross-Sectional Study Based on Machine Learning Approach. BMC Infect. Dis. 2023, 23, 18. [Google Scholar] [CrossRef]
- La Vignera, S.; Cannarella, R.; Condorelli, R.A.; Torre, F.; Aversa, A.; Calogero, A.E. Sex-Specific SARS-CoV-2 Mortality: Among Hormone-Modulated ACE2 Expression, Risk of Venous Thromboembolism and Hypovitaminosis D. Int. J. Mol. Sci. 2020, 21, 2948. [Google Scholar] [CrossRef]
- Hayden, M.R. Endothelial Activation and Dysfunction in Metabolic Syndrome, Type 2 Diabetes and Coronavirus Disease 2019. J. Int. Med. Res. 2020, 48, 300060520939746. [Google Scholar] [CrossRef] [PubMed]
- Weiss, P.; Murdoch, D.R. Clinical Course and Mortality Risk of Severe COVID-19. Lancet 2020, 395, 1014–1015. [Google Scholar] [CrossRef] [PubMed]
- Chavda, V.; Chaurasia, B.; Fiorindi, A.; Umana, G.E.; Lu, B.; Montemurro, N. Ischemic Stroke and SARS-CoV-2 Infection: The Bidirectional Pathology and Risk Morbidities. Neurol. Int. 2022, 14, 391–405. [Google Scholar] [CrossRef]
- Kumar, N.; Verma, R.; Lohana, P.; Lohana, A.; Ramphul, K. Acute Myocardial Infarction in COVID-19 Patients. A Review of Cases in the Literature. Arch. Med. Sci. Atheroscler. Dis. 2021, 6, e169–e175. [Google Scholar] [CrossRef] [PubMed]
- Tousoulis, D.; Kampoli, A.M.; Papageorgiou, N.; Androulakis, E.; Antoniades, C.; Toutouzas, K.; Stefanadis, C. Pathophysiology of Atherosclerosis: The Role of Inflammation. Curr. Pharm. Des. 2011, 17, 4089–4110. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.; Bae, J.H.; Kwon, H.S.; Nauck, M.A. COVID-19 and Diabetes Mellitus: From Pathophysiology to Clinical Management. Nat. Rev. Endocrinol. 2021, 17, 11–30. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.J.; Wu, Z.Y.; Nie, X.W.; Bian, J.S. Role of Endothelial Dysfunction in Cardiovascular Diseases: The Link Between Inflammation and Hydrogen Sulfide. Front. Pharmacol. 2020, 10, 1568. [Google Scholar] [CrossRef]
- Mengozzi, A.; Masi, S.; Virdis, A. Obesity-Related Endothelial Dysfunction: Moving from Classical to Emerging Mechanisms. Endocr. Metab. Sci. 2020, 1, 100063. [Google Scholar] [CrossRef]
- Varga, Z.; Flammer, A.J.; Steiger, P.; Haberecker, M.; Andermatt, R.; Zinkernagel, A.S.; Mehra, M.R.; Schuepbach, R.A.; Ruschitzka, F.; Moch, H. Endothelial Cell Infection and Endotheliitis in COVID-19. Lancet 2020, 395, 1417–1418. [Google Scholar] [CrossRef]
- Wadowski, P.P.; Panzer, B.; Jozkowicz, A.; Kopp, C.W.; Gremmel, T.; Panzer, S.; Koppensteiner, R. Microvascular Thrombosis as a Critical Factor in Severe COVID-19. Int. J. Mol. Sci. 2023, 24, 2492. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zheng, Y.; Gou, X.; Pu, K.; Chen, Z.; Guo, Q.; Ji, R.; Wang, H.; Wang, Y.; Zhou, Y. Prevalence of Comorbidities and Its Effects in Patients Infected with SARS-CoV-2: A Systematic Review and Meta-Analysis. Int. J. Infect. Dis. 2020, 94, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Chen, X.; Cai, Y.; Xia, J.; Zhou, X.; Xu, S.; Huang, H.; Zhang, L.; Du, C.; Zhang, Y.; et al. Risk Factors Associated with Acute Respiratory Distress Syndrome and Death in Patients with Coronavirus Disease 2019 Pneumonia in Wuhan, China. JAMA Intern. Med. 2020, 180, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Kai, H.; Kai, M. Interactions of Coronaviruses with ACE2, Angiotensin II, and RAS Inhibitors-Lessons from Available Evidence and Insights into COVID-19. Hypertens. Res. 2020, 43, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, Y.N.; Yan, R.; Wang, G.; Zhang, Y.; Zhang, Z.R.; Li, Y.; Ou, J.; Chu, W.; Liang, Z.; et al. ACE2-Targeting Monoclonal Antibody as Potent and Broad-Spectrum Coronavirus Blocker. Signal Transduct. Target. Ther. 2021, 6, 315. [Google Scholar] [CrossRef]
- Bastolla, U.; Chambers, P.; Abia, D.; García-Bermejo, M.L.; Fresno, M. Is COVID-19 Severity Associated with ACE2 Degradation? Front. Drug Discov. 2022, 1, 789710. [Google Scholar] [CrossRef]
- Lee, I.T.; Nakayama, T.; Wu, C.T.; Goltsev, Y.; Jiang, S.; Gall, P.A.; Liao, C.K.; Shih, L.C.; Schurch, C.M.; McIlwain, D.R.; et al. ACE2 Localizes to the Respiratory Cilia and Is Not Increased by ACE Inhibitors or ARBs. Nat. Commun. 2020, 11, 5453. [Google Scholar] [CrossRef]
- Beyerstedt, S.; Casaro, E.B.; Rangel, E.B. COVID-19: Angiotensin-Converting Enzyme 2 (ACE2) Expression and Tissue Susceptibility to SARS-CoV-2 Infection. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 905–919. [Google Scholar] [CrossRef]
- Kornilov, S.A.; Lucas, I.; Jade, K.; Dai, C.L.; Lovejoy, J.C.; Magis, A.T. Plasma Levels of Soluble ACE2are Associated with Sex, Metabolic Syndrome, and Its Biomarkers in a Large Cohort, Pointing to a Possible Mechanism for Increased Severity in COVID-19. Crit. Care 2020, 24, 452. [Google Scholar] [CrossRef]
- Libby, P.; Luscher, T. COVID-19 Is, in the End, an Endothelial Disease. Eur. Heart J. 2020, 41, 3038–3044. [Google Scholar] [CrossRef]
- Tetzlaff, F.; Fischer, A. Control of Blood Vessel Formation by Notch Signaling. Adv. Exp. Med. Biol. 2018, 1066, 319–338. [Google Scholar] [CrossRef]
- Jourde-Chiche, N.; Fakhouri, F.; Dou, L.; Bellien, J.; Burtey, S.; Frimat, M.; Jarrot, P.A.; Kaplanski, G.; Le Quintrec, M.; Pernin, V.; et al. Endothelium Structure and Function in Kidney Health and Disease. Nat. Rev. Nephrol. 2019, 15, 87–108. [Google Scholar] [CrossRef]
- Iwakiri, Y. Nitric Oxide in Liver Fibrosis: The Role of Inducible Nitric Oxide Synthase. Clin. Mol. Hepatol. 2015, 21, 319–325. [Google Scholar] [CrossRef]
- Sanghavi, D.K.; Titus, A.; Caulfield, T.R.; David Freeman, W. Endotheliitis, Endothelin, and Endothelin Receptor Blockers in COVID-19. Med. Hypotheses 2021, 150, 110564. [Google Scholar] [CrossRef]
- Mosleh, W.; Chen, K.; Pfau, S.E.; Vashist, A. Endotheliitis and Endothelial Dysfunction in Patients with COVID-19: Its Role in Thrombosis and Adverse Outcomes. J. Clin. Med. 2020, 9, 1862. [Google Scholar] [CrossRef]
- Ritter, A.; Kreis, N.N.; Louwen, F.; Yuan, J. Obesity and COVID-19: Molecular Mechanisms Linking Both Pandemics. Int. J. Mol. Sci. 2020, 21, 5793. [Google Scholar] [CrossRef] [PubMed]
- South, A.M.; Tomlinson, L.; Edmonston, D.; Hiremath, S.; Sparks, M.A. Controversies of Renin-Angiotensin System Inhibition during the COVID-19 Pandemic. Nat. Rev. Nephrol. 2020, 16, 305–307. [Google Scholar] [CrossRef] [PubMed]
- Verdoni, L.; Mazza, A.; Gervasoni, A.; Martelli, L.; Ruggeri, M.; Ciuffreda, M.; Bonanomi, E.; D’Antiga, L. An Outbreak of Severe Kawasaki-like Disease at the Italian Epicentre of the SARS-CoV-2 Epidemic: An Observational Cohort Study. Lancet 2020, 395, 1771–1778. [Google Scholar] [CrossRef]
- Coperchini, F.; Chiovato, L.; Croce, L.; Magri, F.; Rotondi, M. The Cytokine Storm in COVID-19: An Overview of the Involvement of the Chemokine/Chemokine-Receptor System. Cytokine Growth Factor Rev. 2020, 53, 25–32. [Google Scholar] [CrossRef] [PubMed]
- McGonagle, D.; Sharif, K.; O’Regan, A.; Bridgewood, C. The Role of Cytokines Including Interleukin-6 in COVID-19 Induced Pneumonia and Macrophage Activation Syndrome-Like Disease. Autoimmun. Rev. 2020, 19, 102537. [Google Scholar] [CrossRef]
- Levy, T. The Importance of HPV Testing for Cervical Cancer Screening in Israel. Isr. Med. Assoc. J. 2017, 19, 465–644. [Google Scholar]
- Alam, W. Hypercoagulability in COVID-19: A Review of the Potential Mechanisms Underlying Clotting Disorders. SAGE Open Med 2021, 9, 20503121211002996. [Google Scholar] [CrossRef] [PubMed]
- Davidson, M.; Menon, S.; Chaimani, A.; Evrenoglou, T.; Ghosn, L.; Grana, C.; Henschke, N.; Cogo, E.; Villanueva, G.; Ferrand, G.; et al. Interleukin-1 Blocking Agents for Treating COVID-19. Cochrane Database Syst. Rev. 2022, 1, CD015308. [Google Scholar] [CrossRef] [PubMed]
- Ghosn, L.; Chaimani, A.; Evrenoglou, T.; Davidson, M.; Grana, C.; Schmucker, C.; Bollig, C.; Henschke, N.; Sguassero, Y.; Nejstgaard, C.H.; et al. Interleukin-6 Blocking Agents for Treating COVID-19: A Living Systematic Review. Cochrane Database Syst. Rev. 2021, 3, CD013881. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, A.; Lin, M.I.; Murata, T.; Landskroner-Eiger, S.; Schleicher, M.; Kothiya, M.; Iwakiri, Y.; Yu, J.; Huang, P.L.; Sessa, W.C. ENOS-Derived Nitric Oxide Regulates Endothelial Barrier Function through VE-Cadherin and Rho GTPases. J. Cell Sci. 2013, 126, 5541–5552. [Google Scholar] [CrossRef] [PubMed]
- GBD 2017 Disease and Injury Incidence and Prevalence Collaborators. Global, Regional, and National Incidence, Prevalence, and Years Lived with Disability for 354 Diseases and Injuries for 195 Countries and Territories, 1990–2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet 2019, 392, 1789–1858. [Google Scholar] [CrossRef]
- van Hinsbergh, V.W. Endothelium--Role in Regulation of Coagulation and Inflammation. Semin. Immunopathol. 2012, 34, 93–106. [Google Scholar] [CrossRef]
- Drozdz, D.; Latka, M.; Drozdz, T.; Sztefko, K.; Kwinta, P. Thrombomodulin as a New Marker of Endothelial Dysfunction in Chronic Kidney Disease in Children. Oxid. Med. Cell Longev. 2018, 2018, 1619293. [Google Scholar] [CrossRef] [PubMed]
- Mucka, S.; Miodonska, M.; Jakubiak, G.K.; Starzak, M.; Cieslar, G.; Stanek, A. Endothelial Function Assessment by Flow-Mediated Dilation Method: A Valuable Tool in the Evaluation of the Cardiovascular System. Int. J. Environ. Res. Public Health 2022, 19, 11242. [Google Scholar] [CrossRef]
- Miyauchi, T.; Sakai, S. Endothelin and the Heart in Health and Diseases. Peptides 2019, 111, 77–88. [Google Scholar] [CrossRef]
- Soriano, J.B.; Murthy, S.; Marshall, J.C.; Relan, P.; Diaz, J.V. A Clinical Case Definition of Post-COVID-19 Condition by a Delphi Consensus. Lancet Infect. Dis. 2022, 22, e102–e107. [Google Scholar] [CrossRef] [PubMed]
- Sudre, C.H.; Murray, B.; Varsavsky, T.; Graham, M.S.; Penfold, R.S.; Bowyer, R.C.; Pujol, J.C.; Klaser, K.; Antonelli, M.; Canas, L.S.; et al. Attributes and Predictors of Long COVID. Nat. Med. 2021, 27, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Ballering, A.V.; van Zon, S.K.R.; Olde Hartman, T.C.; Rosmalen, J.G.M. Persistence of Somatic Symptoms after COVID-19 in the Netherlands: An Observational Cohort Study. Lancet 2022, 400, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Ceban, F.; Ling, S.; Lui, L.M.W.; Lee, Y.; Gill, H.; Teopiz, K.M.; Rodrigues, N.B.; Subramaniapillai, M.; Di Vincenzo, J.D.; Cao, B.; et al. Fatigue and Cognitive Impairment in Post-COVID-19 Syndrome: A Systematic Review and Meta-Analysis. Brain Behav. Immun. 2022, 101, 93–135. [Google Scholar] [CrossRef] [PubMed]
- Silva Andrade, B.; Siqueira, S.; de Assis Soares, W.R.; de Souza Rangel, F.; Santos, N.O.; Dos Santos Freitas, A.; Ribeiro da Silveira, P.; Tiwari, S.; Alzahrani, K.J.; Goes-Neto, A.; et al. Long-COVID and Post-COVID Health Complications: An Up-to-Date Review on Clinical Conditions and Their Possible Molecular Mechanisms. Viruses 2021, 13, 700. [Google Scholar] [CrossRef] [PubMed]
- Asadi-Pooya, A.A.; Akbari, A.; Emami, A.; Lotfi, M.; Rostamihosseinkhani, M.; Nemati, H.; Barzegar, Z.; Kabiri, M.; Zeraatpisheh, Z.; Farjoud-Kouhanjani, M.; et al. Long COVID Syndrome-Associated Brain Fog. J. Med. Virol. 2022, 94, 979–984. [Google Scholar] [CrossRef]
- Notarte, K.I.; Catahay, J.A.; Velasco, J.V.; Pastrana, A.; Ver, A.T.; Pangilinan, F.C.; Peligro, P.J.; Casimiro, M.; Guerrero, J.J.; Gellaco, M.M.L.; et al. Impact of COVID-19 Vaccination on the Risk of Developing Long-COVID and on Existing Long-COVID Symptoms: A Systematic Review. EClinicalMedicine 2022, 53, 101624. [Google Scholar] [CrossRef] [PubMed]
- Cyranoski, D. Why Children Avoid the Worst Coronavirus Complications Might Lie in Their Arteries. Nature 2020, 582, 324–325. [Google Scholar] [CrossRef]
- Selskyy, B.; Kostiv, S.; Ivankiv, Y.; Pak, A.; Palii, S.; Shevchuk, O.; Korda, M.; Vari, S. Diagnosis of Endothelial Dysfunction with Endothelium-Dependent Vasodilation in Post-COVID-19 Patients. In Proceedings of the 17th RECOOP Bridges in Life Sciences, Prague, Czech Republic, 21 April 2022; p. 72. [Google Scholar]
- Zheng, Y.Y.; Ma, Y.T.; Zhang, J.Y.; Xie, X. COVID-19 and the Cardiovascular System. Nat. Rev. Cardiol. 2020, 17, 259–260. [Google Scholar] [CrossRef]
- D’Marco, L.; Puchades, M.J.; Romero-Parra, M.; Gorriz, J.L. Diabetic Kidney Disease and COVID-19: The Crash of Two Pandemics. Front. Med. 2020, 7, 199. [Google Scholar] [CrossRef]
- Castanares-Zapatero, D.; Chalon, P.; Kohn, L.; Dauvrin, M.; Detollenaere, J.; Maertens de Noordhout, C.; Primus-de Jong, C.; Cleemput, I.; Van den Heede, K. Pathophysiology and Mechanism of Long COVID: A Comprehensive Review. Ann. Med. 2022, 54, 1473–1487. [Google Scholar] [CrossRef] [PubMed]
- Mangalmurti, N.; Hunter, C.A. Cytokine Storms: Understanding COVID-19. Immunity 2020, 53, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, H.; Chung, M.; Jeon, N.L. Engineering of Functional, Perfusable 3D Microvascular Networks on a Chip. Lab Chip 2013, 13, 1489–1500. [Google Scholar] [CrossRef] [PubMed]
- Aymerich, M.; Alvarez, E.; Bao-Varela, C.; Moscoso, I.; Gonzalez-Juanatey, J.R.; Flores-Arias, M.T. Laser Technique for the Fabrication of Blood Vessels-like Models for Preclinical Studies of Pathologies under Flow Conditions. Biofabrication 2017, 9, 25033. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.B.; Wang, X.; Faley, S.; Baer, B.; Balikov, D.A.; Sung, H.J.; Bellan, L.M. Development of 3D Microvascular Networks within Gelatin Hydrogels Using Thermoresponsive Sacrificial Microfibers. Adv. Healthc. Mater. 2016, 5, 781–785. [Google Scholar] [CrossRef]
- Zhang, Y.S.; Arneri, A.; Bersini, S.; Shin, S.R.; Zhu, K.; Goli-Malekabadi, Z.; Aleman, J.; Colosi, C.; Busignani, F.; Dell’Erba, V.; et al. Bioprinting 3D Microfibrous Scaffolds for Engineering Endothelialized Myocardium and Heart-on-a-Chip. Biomaterials 2016, 110, 45–59. [Google Scholar] [CrossRef]
- Dobos, A.; Gantner, F.; Markovic, M.; Van Hoorick, J.; Tytgat, L.; Van Vlierberghe, S.; Ovsianikov, A. On-Chip High-Definition Bioprinting of Microvascular Structures. Biofabrication 2021, 13, 15016. [Google Scholar] [CrossRef]
- Hu, C.; Chen, Y.; Tan, M.J.A.; Ren, K.; Wu, H. Microfluidic Technologies for Vasculature Biomimicry. Analyst 2019, 144, 4461–4471. [Google Scholar] [CrossRef]
- Li, L.M.; Wang, X.Y.; Hu, L.S.; Chen, R.S.; Huang, Y.; Chen, S.J.; Huang, W.H.; Huo, K.F.; Chu, P.K. Vascular Lumen Simulation and Highly-Sensitive Nitric Oxide Detection Using Three-Dimensional Gelatin Chip Coupled to TiC/C Nanowire Arrays Microelectrode. Lab Chip 2012, 12, 4249–4256. [Google Scholar] [CrossRef]
- Baker, B.M.; Trappmann, B.; Stapleton, S.C.; Toro, E.; Chen, C.S. Microfluidics Embedded within Extracellular Matrix to Define Vascular Architectures and Pattern Diffusive Gradients. Lab Chip 2013, 13, 3246–3252. [Google Scholar] [CrossRef]
- Molteni, R.; Bianchi, E.; Patete, P.; Fabbri, M.; Baroni, G.; Dubini, G.; Pardi, R. A Novel Device to Concurrently Assess Leukocyte Extravasation and Interstitial Migration within a Defined 3D Environment. Lab Chip 2015, 15, 195–207. [Google Scholar] [CrossRef]
- Bai, J.; Khajavi, M.; Sui, L.; Fu, H.; Tarakkad Krishnaji, S.; Birsner, A.E.; Bazinet, L.; Kamm, R.D.; D’Amato, R.J. Angiogenic Responses in a 3D Micro-Engineered Environment of Primary Endothelial Cells and Pericytes. Angiogenesis 2021, 24, 111–127. [Google Scholar] [CrossRef] [PubMed]
- van Dijk, C.G.M.; Brandt, M.M.; Poulis, N.; Anten, J.; van der Moolen, M.; Kramer, L.; Homburg, E.; Louzao-Martinez, L.; Pei, J.; Krebber, M.M.; et al. A New Microfluidic Model That Allows Monitoring of Complex Vascular Structures and Cell Interactions in a 3D Biological Matrix. Lab Chip 2020, 20, 1827–1844. [Google Scholar] [CrossRef] [PubMed]
- Takehara, H.; Sakaguchi, K.; Sekine, H.; Okano, T.; Shimizu, T. Microfluidic Vascular-Bed Devices for Vascularized 3D Tissue Engineering: Tissue Engineering on a Chip. Biomed. Microdevices 2019, 22, 9. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Shridhar, S.V.; Gerwitz, M.; Soman, P. An In Vitro Vascular Chip Using 3D Printing-Enabled Hydrogel Casting. Biofabrication 2016, 8, 35015. [Google Scholar] [CrossRef]
- Abudupataer, M.; Chen, N.; Yan, S.; Alam, F.; Shi, Y.; Wang, L.; Lai, H.; Li, J.; Zhu, K.; Wang, C. Bioprinting a 3D Vascular Construct for Engineering a Vessel-on-a-Chip. Biomed. Microdevices 2019, 22, 10. [Google Scholar] [CrossRef]
- Kinoshita, K.; Iwase, M.; Yamada, M.; Yajima, Y.; Seki, M. Fabrication of Multilayered Vascular Tissues Using Microfluidic Agarose Hydrogel Platforms. Biotechnol. J. 2016, 11, 1415–1423. [Google Scholar] [CrossRef] [PubMed]
- Nie, J.; Gao, Q.; Wang, Y.; Zeng, J.; Zhao, H.; Sun, Y.; Shen, J.; Ramezani, H.; Fu, Z.; Liu, Z.; et al. Vessel-on-a-Chip with Hydrogel-Based Microfluidics. Small 2018, 14, e1802368. [Google Scholar] [CrossRef]
- Das, P.; van der Meer, A.D.; Vivas, A.; Arik, Y.B.; Remigy, J.C.; Lahitte, J.F.; Lammertink, R.G.H.; Bacchin, P. Tunable Microstructured Membranes in Organs-on-Chips to Monitor Transendothelial Hydraulic Resistance. Tissue Eng. Part A 2019, 25, 1635–1645. [Google Scholar] [CrossRef]
- Bulboaca, A.E.; Boarescu, P.M.; Melincovici, C.S.; Mihu, C.M. Microfluidic Endothelium-on-a-Chip Development, from in Vivo to In Vitro Experimental Models. Rom. J. Morphol. Embryol. 2020, 61, 15–23. [Google Scholar] [CrossRef]
- Song, J.W.; Daubriac, J.; Tse, J.M.; Bazou, D.; Munn, L.L. RhoA Mediates Flow-Induced Endothelial Sprouting in a 3-D Tissue Analogue of Angiogenesis. Lab Chip 2012, 12, 5000–5006. [Google Scholar] [CrossRef] [PubMed]
- Tourovskaia, A.; Fauver, M.; Kramer, G.; Simonson, S.; Neumann, T. Tissue-Engineered Microenvironment Systems for Modeling Human Vasculature. Exp. Biol. Med. 2014, 239, 1264–1271. [Google Scholar] [CrossRef] [PubMed]
- van Engeland, N.C.A.; Pollet, A.; den Toonder, J.M.J.; Bouten, C.V.C.; Stassen, O.; Sahlgren, C.M. A Biomimetic Microfluidic Model to Study Signalling between Endothelial and Vascular Smooth Muscle Cells under Hemodynamic Conditions. Lab Chip 2018, 18, 1607–1620. [Google Scholar] [CrossRef] [PubMed]
- Sano, E.; Mori, C.; Nashimoto, Y.; Yokokawa, R.; Kotera, H.; Torisawa, Y.S. Engineering of Vascularized 3D Cell Constructs to Model Cellular Interactions through a Vascular Network. Biomicrofluidics 2018, 12, 42204. [Google Scholar] [CrossRef]
- Mao, M.; Bei, H.P.; Lam, C.H.; Chen, P.; Wang, S.; Chen, Y.; He, J.; Zhao, X. Human-on-Leaf-Chip: A Biomimetic Vascular System Integrated with Chamber-Specific Organs. Small 2020, 16, e2000546. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.; Park, J.K. Fabrication of a Perfusable 3D In Vitro Artery-Mimicking Multichannel System for Artery Disease Models. ACS Biomater. Sci. Eng. 2020, 6, 5326–5336. [Google Scholar] [CrossRef] [PubMed]
- Ellis, B.W.; Acun, A.; Can, U.I.; Zorlutuna, P. Human IPSC-Derived Myocardium-on-Chip with Capillary-like Flow for Personalized Medicine. Biomicrofluidics 2017, 11, 24105. [Google Scholar] [CrossRef]
- Shemesh, J.; Jalilian, I.; Shi, A.; Heng Yeoh, G.; Knothe Tate, M.L.; Ebrahimi Warkiani, M. Flow-Induced Stress on Adherent Cells in Microfluidic Devices. Lab Chip 2015, 15, 4114–4127. [Google Scholar] [CrossRef]
- Schimek, K.; Busek, M.; Brincker, S.; Groth, B.; Hoffmann, S.; Lauster, R.; Lindner, G.; Lorenz, A.; Menzel, U.; Sonntag, F.; et al. Integrating Biological Vasculature into a Multi-Organ-Chip Microsystem. Lab Chip 2013, 13, 3588–3598. [Google Scholar] [CrossRef]
- Salipante, P.F.; Hudson, S.D.; Alimperti, S. Blood Vessel-on-a-Chip Examines the Biomechanics of Microvasculature. Soft Matter 2021, 18, 117–125. [Google Scholar] [CrossRef]
- Salman, M.M.; Marsh, G.; Kusters, I.; Delince, M.; Di Caprio, G.; Upadhyayula, S.; de Nola, G.; Hunt, R.; Ohashi, K.G.; Gray, T.; et al. Design and Validation of a Human Brain Endothelial Microvessel-on-a-Chip Open Microfluidic Model Enabling Advanced Optical Imaging. Front. Bioeng. Biotechnol. 2020, 8, 573775. [Google Scholar] [CrossRef]
- Berse, B.; Brown, L.F.; Van de Water, L.; Dvorak, H.F.; Senger, D.R. Vascular Permeability Factor (Vascular Endothelial Growth Factor) Gene Is Expressed Differentially in Normal Tissues, Macrophages, and Tumors. Mol. Biol. Cell 1992, 3, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Papadimitrou, J.M.; Woods, A.E. Structural and Functional Characteristics of the Microcirculation in Neoplasms. J. Pathol. 1975, 116, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.M.; Mavrogiannis, N.; Gagnon, Z.; Gerecht, S. Microfluidic Platform for the Real Time Measurement and Observation of Endothelial Barrier Function under Shear Stress. Biomicrofluidics 2018, 12, 42202. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.F.; Mohan, M.D.; Young, E.W.K.; Simmons, C.A. Integrated Electrochemical Measurement of Endothelial Permeability in a 3D Hydrogel-Based Microfluidic Vascular Model. Biosens. Bioelectron. 2020, 147, 111757. [Google Scholar] [CrossRef]
- Lee, H.; Kim, S.; Chung, M.; Kim, J.H.; Jeon, N.L. A Bioengineered Array of 3D Microvessels for Vascular Permeability Assay. Microvasc. Res. 2014, 91, 90–98. [Google Scholar] [CrossRef]
- Varone, A.; Nguyen, J.K.; Leng, L.; Barrile, R.; Sliz, J.; Lucchesi, C.; Wen, N.; Gravanis, A.; Hamilton, G.A.; Karalis, K.; et al. A Novel Organ-Chip System Emulates Three-Dimensional Architecture of the Human Epithelia and the Mechanical Forces Acting on It. Biomaterials 2021, 275, 120957. [Google Scholar] [CrossRef]
- Tronolone, J.J.; Jain, A. Engineering New Microvascular Networks On-Chip: Ingredients, Assembly, and Best Practices. Adv. Funct. Mater. 2021, 31, 2007199. [Google Scholar] [CrossRef]
- Haase, K.; Piatti, F.; Marcano, M.; Shin, Y.; Visone, R.; Redaelli, A.; Rasponi, M.; Kamm, R.D. Physiologic Flow-Conditioning Limits Vascular Dysfunction in Engineered Human Capillaries. Biomaterials 2022, 280, 121248. [Google Scholar] [CrossRef]
- Kim, S.; Ko, J.; Lee, S.R.; Park, D.; Park, S.; Jeon, N.L. Anchor-IMPACT: A Standardized Microfluidic Platform for High-Throughput Antiangiogenic Drug Screening. Biotechnol. Bioeng. 2021, 118, 2524–2535. [Google Scholar] [CrossRef]
- Ko, J.; Lee, Y.; Lee, S.; Lee, S.R.; Jeon, N.L. Human Ocular Angiogenesis-Inspired Vascular Models on an Injection-Molded Microfluidic Chip. Adv. Healthc. Mater. 2019, 8, e1900328. [Google Scholar] [CrossRef] [PubMed]
- Ryu, H.; Oh, S.; Lee, H.J.; Lee, J.Y.; Lee, H.K.; Jeon, N.L. Engineering a Blood Vessel Network Module for Body-on-a-Chip Applications. J. Lab. Autom. 2015, 20, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Hasenberg, T.; Muhleder, S.; Dotzler, A.; Bauer, S.; Labuda, K.; Holnthoner, W.; Redl, H.; Lauster, R.; Marx, U. Emulating Human Microcapillaries in a Multi-Organ-Chip Platform. J. Biotechnol. 2015, 216, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Phan, D.T.; Sobrino, A.; George, S.C.; Hughes, C.C.; Lee, A.P. Engineering Anastomosis between Living Capillary Networks and Endothelial Cell-Lined Microfluidic Channels. Lab Chip 2016, 16, 282–290. [Google Scholar] [CrossRef]
- Bogorad, M.I.; DeStefano, J.; Wong, A.D.; Searson, P.C. Tissue-Engineered 3D Microvessel and Capillary Network Models for the Study of Vascular Phenomena. Microcirculation 2017, 24, e12360. [Google Scholar] [CrossRef]
- Chrobak, K.M.; Potter, D.R.; Tien, J. Formation of Perfused, Functional Microvascular Tubes In Vitro. Microvasc. Res. 2006, 71, 185–196. [Google Scholar] [CrossRef]
- de Haan, L.; Suijker, J.; van Roey, R.; Berges, N.; Petrova, E.; Queiroz, K.; Strijker, W.; Olivier, T.; Poeschke, O.; Garg, S.; et al. A Microfluidic 3D Endothelium-on-a-Chip Model to Study Transendothelial Migration of T Cells in Health and Disease. Int. J. Mol. Sci. 2021, 22, 8234. [Google Scholar] [CrossRef]
- Poussin, C.; Kramer, B.; Lanz, H.L.; Van den Heuvel, A.; Laurent, A.; Olivier, T.; Vermeer, M.; Peric, D.; Baumer, K.; Dulize, R.; et al. 3D Human Microvessel-on-a-Chip Model for Studying Monocyte-to-Endothelium Adhesion under Flow—Application in Systems Toxicology. ALTEX 2020, 37, 47–63. [Google Scholar] [CrossRef]
- Genes, L.I.; Tolan, V.N.; Hulvey, M.K.; Martin, R.S.; Spence, D.M. Addressing a Vascular Endothelium Array with Blood Components Using Underlying Microfluidic Channels. Lab Chip 2007, 7, 1256–1259. [Google Scholar] [CrossRef]
- Bersini, S.; Moretti, M. 3D Functional and Perfusable Microvascular Networks for Organotypic Microfluidic Models. J. Mater. Sci. Mater. Med. 2015, 26, 180. [Google Scholar] [CrossRef]
- Aird, W.C. Phenotypic Heterogeneity of the Endothelium: II. Representative Vascular Beds. Circ. Res. 2007, 100, 174–190. [Google Scholar] [CrossRef] [PubMed]
- Aird, W.C. Phenotypic Heterogeneity of the Endothelium: I. Structure, Function, and Mechanisms. Circ. Res. 2007, 100, 158–173. [Google Scholar] [CrossRef] [PubMed]
- DeStefano, J.G.; Xu, Z.S.; Williams, A.J.; Yimam, N.; Searson, P.C. Effect of Shear Stress on IPSC-Derived Human Brain Microvascular Endothelial Cells (DhBMECs). Fluids Barriers CNS 2017, 14, 20. [Google Scholar] [CrossRef] [PubMed]
- Reinitz, A.; DeStefano, J.; Ye, M.; Wong, A.D.; Searson, P.C. Human Brain Microvascular Endothelial Cells Resist Elongation Due to Shear Stress. Microvasc. Res. 2015, 99, 8–18. [Google Scholar] [CrossRef]
- Kim, S.; Chung, M.; Ahn, J.; Lee, S.; Jeon, N.L. Interstitial Flow Regulates the Angiogenic Response and Phenotype of Endothelial Cells in a 3D Culture Model. Lab Chip 2016, 16, 4189–4199. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, L.; Inglebert, M.; Scrimieri, R.; Sinha, P.K.; Zuccotti, G.V.; Milani, P.; Bureau, L.; Misbah, C.; Maier, J.A.M. Human Endothelial Cells in High Glucose: New Clues from Culture in 3D Microfluidic Chips. FASEB J. 2022, 36, e22137. [Google Scholar] [CrossRef]
- Moses, S.R.; Adorno, J.J.; Palmer, A.F.; Song, J.W. Vessel-on-a-Chip Models for Studying Microvascular Physiology, Transport, and Function In Vitro. Am. J Physiol. Cell Physiol. 2021, 320, C92–C105. [Google Scholar] [CrossRef]
- Sart, S.; Tomasi, R.F.; Amselem, G.; Baroud, C.N. Multiscale Cytometry and Regulation of 3D Cell Cultures on a Chip. Nat. Commun. 2017, 8, 469. [Google Scholar] [CrossRef]
- Cochrane, A.; Albers, H.J.; Passier, R.; Mummery, C.L.; van den Berg, A.; Orlova, V.V.; van der Meer, A.D. Advanced In Vitro Models of Vascular Biology: Human Induced Pluripotent Stem Cells and Organ-on-Chip Technology. Adv. Drug Deliv. Rev. 2019, 140, 68–77. [Google Scholar] [CrossRef]
- Paloschi, V.; Sabater-Lleal, M.; Middelkamp, H.; Vivas, A.; Johansson, S.; van der Meer, A.; Tenje, M.; Maegdefessel, L. Organ-on-a-Chip Technology: A Novel Approach to Investigate Cardiovascular Diseases. Cardiovasc. Res. 2021, 117, 2742–2754. [Google Scholar] [CrossRef]
- Mastrangeli, M.; Millet, S.; Orchid Partners, T.; Van den Eijnden-van Raaij, J. Organ-on-Chip in Development: Towards a Roadmap for Organs-on-Chip. ALTEX 2019, 36, 650–668. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, Y.K.; Yin, R.T.; Shang, M.R.; Shirure, V.S.; Moya, M.L.; George, S.C. Human Induced Pluripotent Stem Cell-Derived Endothelial Cells for Three-Dimensional Microphysiological Systems. Tissue Eng. Part C Methods 2017, 23, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Pars, S.; Achberger, K.; Kleger, A.; Liebau, S.; Pashkovskaia, N. Generation of Functional Vascular Endothelial Cells and Pericytes from Keratinocyte Derived Human Induced Pluripotent Stem Cells. Cells 2021, 10, 74. [Google Scholar] [CrossRef]
- Jeon, J.S.; Bersini, S.; Gilardi, M.; Dubini, G.; Charest, J.L.; Moretti, M.; Kamm, R.D. Human 3D Vascularized Organotypic Microfluidic Assays to Study Breast Cancer Cell Extravasation. Proc. Natl. Acad. Sci. USA 2015, 112, 214–219. [Google Scholar] [CrossRef]
- de Graaf, M.N.S.; Cochrane, A.; van den Hil, F.E.; Buijsman, W.; van der Meer, A.D.; van den Berg, A.; Mummery, C.L.; Orlova, V.V. Scalable Microphysiological System to Model Three-Dimensional Blood Vessels. APL Bioeng. 2019, 3, 26105. [Google Scholar] [CrossRef] [PubMed]
- Vila Cuenca, M.; Cochrane, A.; van den Hil, F.E.; de Vries, A.A.F.; Lesnik Oberstein, S.A.J.; Mummery, C.L.; Orlova, V.V. Engineered 3D Vessel-on-Chip Using HiPSC-Derived Endothelial- and Vascular Smooth Muscle Cells. Stem Cell Rep. 2021, 16, 2159–2168. [Google Scholar] [CrossRef]
- Weng, K.C.; Kurokawa, Y.K.; Hajek, B.S.; Paladin, J.A.; Shirure, V.S.; George, S.C. Human Induced Pluripotent Stem-Cardiac-Endothelial-Tumor-on-a-Chip to Assess Anticancer Efficacy and Cardiotoxicity. Tissue Eng. Part C Methods 2020, 26, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Doherty, E.L.; Aw, W.Y.; Hickey, A.J.; Polacheck, W.J. Microfluidic and Organ-on-a-Chip Approaches to Investigate Cellular and Microenvironmental Contributions to Cardiovascular Function and Pathology. Front. Bioeng. Biotechnol. 2021, 9, 624435. [Google Scholar] [CrossRef] [PubMed]
- Venugopal Menon, N.; Tay, H.M.; Pang, K.T.; Dalan, R.; Wong, S.C.; Wang, X.; Li, K.H.H.; Hou, H.W. A Tunable Microfluidic 3D Stenosis Model to Study Leukocyte-Endothelial Interactions in Atherosclerosis. APL Bioeng. 2018, 2, 16103. [Google Scholar] [CrossRef]
- Rauti, R.; Ess, A.; Roi, B.L.; Kreinin, Y.; Epshtein, M.; Korin, N.; Maoz, B.M. Transforming a Well into a Chip: A Modular 3D-Printed Microfluidic Chip. APL Bioeng. 2021, 5, 26103. [Google Scholar] [CrossRef]
- Li, Y.; Hu, C.; Wang, P.; Liu, Y.; Wang, L.; Pi, Q.; Gong, Z.; Yang, X.; Mak, M.; Wu, Y. Indoor Nanoscale Particulate Matter-Induced Coagulation Abnormality Based on a Human 3D Microvascular Model on a Microfluidic Chip. J. Nanobiotechnol. 2019, 17, 20. [Google Scholar] [CrossRef]
- Li, Y.; Wu, Y.; Liu, Y.; Deng, Q.H.; Mak, M.; Yang, X. Atmospheric Nanoparticles Affect Vascular Function Using a 3D Human Vascularized Organotypic Chip. Nanoscale 2019, 11, 15537–15549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Liu, W.; Zhou, S.; Jiang, L.; Wang, K.; Wei, Y.; Liu, A.; Wei, W.; Liu, S. Investigation of Environmental Pollutant-Induced Lung Inflammation and Injury in a 3D Coculture-Based Microfluidic Pulmonary Alveolus System. Anal. Chem. 2020, 92, 7200–7208. [Google Scholar] [CrossRef] [PubMed]
- Jung, O.; Tung, Y.T.; Sim, E.; Chen, Y.C.; Lee, E.; Ferrer, M.; Song, M.J. Development of Human-Derived, Three-Dimensional Respiratory Epithelial Tissue Constructs with Perfusable Microvasculature on a High-Throughput Microfluidics Screening Platform. Biofabrication 2022, 14, 025012. [Google Scholar] [CrossRef]
- Lu, R.X.Z.; Lai, B.F.L.; Rafatian, N.; Gustafson, D.; Campbell, S.B.; Banerjee, A.; Kozak, R.; Mossman, K.; Mubareka, S.; Howe, K.L.; et al. Vasculature-on-a-Chip Platform with Innate Immunity Enables Identification of Angiopoietin-1 Derived Peptide as a Therapeutic for SARS-CoV-2 Induced Inflammation. Lab Chip 2022, 22, 1171–1186. [Google Scholar] [CrossRef] [PubMed]
- Buzhdygan, T.P.; DeOre, B.J.; Baldwin-Leclair, A.; Bullock, T.A.; McGary, H.M.; Khan, J.A.; Razmpour, R.; Hale, J.F.; Galie, P.A.; Potula, R.; et al. The SARS-CoV-2 Spike Protein Alters Barrier Function in 2D Static and 3D Microfluidic in-Vitro Models of the Human Blood-Brain Barrier. Neurobiol. Dis. 2020, 146, 105131. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shevchuk, O.; Palii, S.; Pak, A.; Chantada, N.; Seoane, N.; Korda, M.; Campos-Toimil, M.; Álvarez, E. Vessel-on-a-Chip: A Powerful Tool for Investigating Endothelial COVID-19 Fingerprints. Cells 2023, 12, 1297. https://doi.org/10.3390/cells12091297
Shevchuk O, Palii S, Pak A, Chantada N, Seoane N, Korda M, Campos-Toimil M, Álvarez E. Vessel-on-a-Chip: A Powerful Tool for Investigating Endothelial COVID-19 Fingerprints. Cells. 2023; 12(9):1297. https://doi.org/10.3390/cells12091297
Chicago/Turabian StyleShevchuk, Oksana, Svitlana Palii, Anastasiia Pak, Nuria Chantada, Nuria Seoane, Mykhaylo Korda, Manuel Campos-Toimil, and Ezequiel Álvarez. 2023. "Vessel-on-a-Chip: A Powerful Tool for Investigating Endothelial COVID-19 Fingerprints" Cells 12, no. 9: 1297. https://doi.org/10.3390/cells12091297