
Rotating Magnetic Field Mitigates Ankylosing Spondylitis Targeting Osteocytes and Chondrocytes via Ameliorating Immune Dysfunctions
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice and Experimental Design
2.2. Scoring Severity of Bone Disease and Athletic Ability
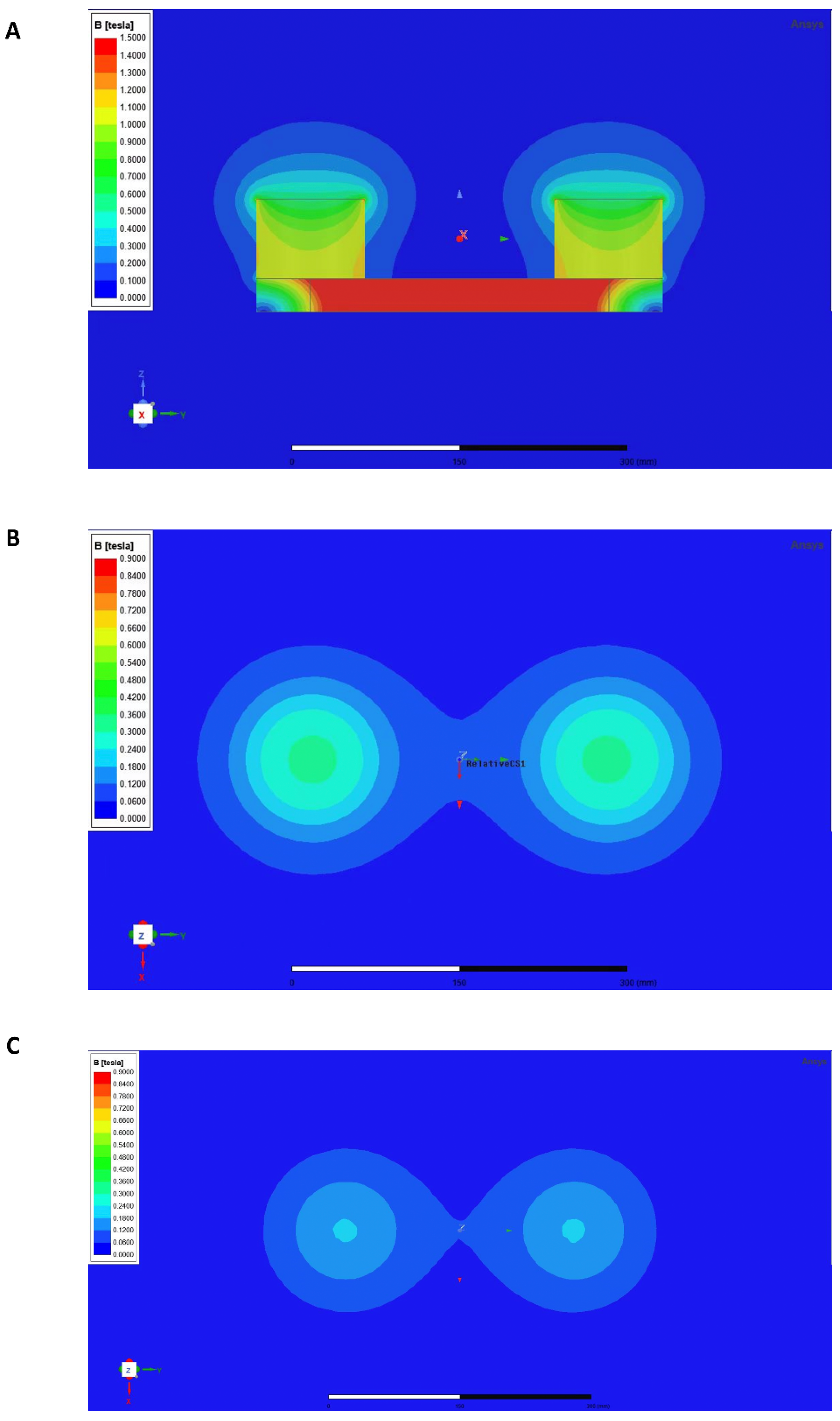
2.3. Rotating Magnetic Field
2.4. Locomotor Behavioral Test
2.5. Radiography Analysis
2.6. Skeletal Preparations
2.7. Histological Staining
2.8. Immunohistochemistry
2.9. Immunofluorescence Staining
2.10. Enzyme-Linked Immunosorbent Assay and Cytometric Bead Array
2.11. Murine Lymphocyte Isolation
2.12. Flow Cytometry Analysis and Antibodies
2.13. Immunoblotting
2.14. Total RNA Extraction and RNA Sequencing
2.15. Quantitative Polymerase Chain Reaction
2.16. Metabolomics Detection
2.17. Statistical Analysis
3. Results
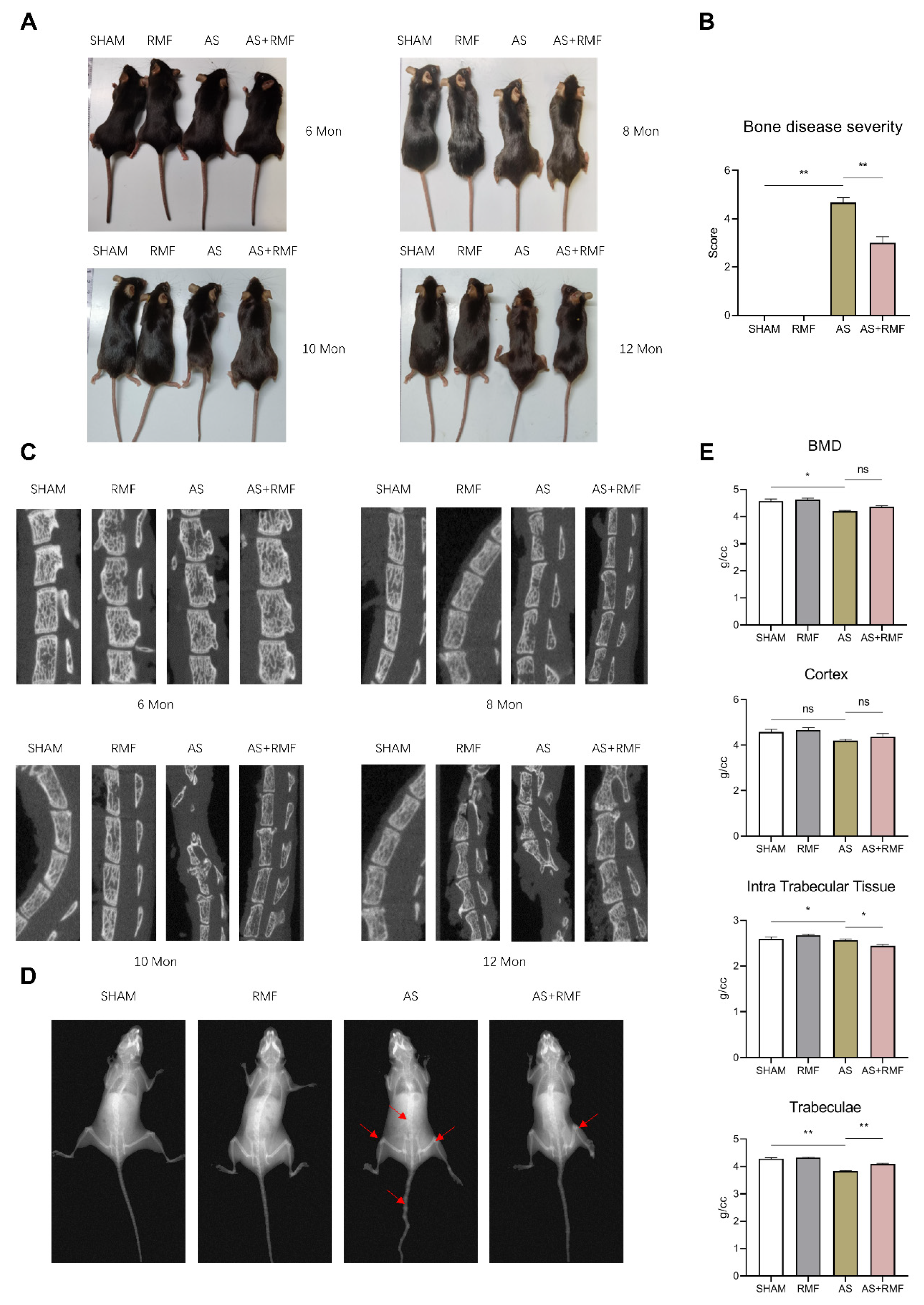
3.1. RMF Alleviates Age-Related AS-like Bone Disease Caused by Ptpn11 Deletion in CD4-Expressing Cells
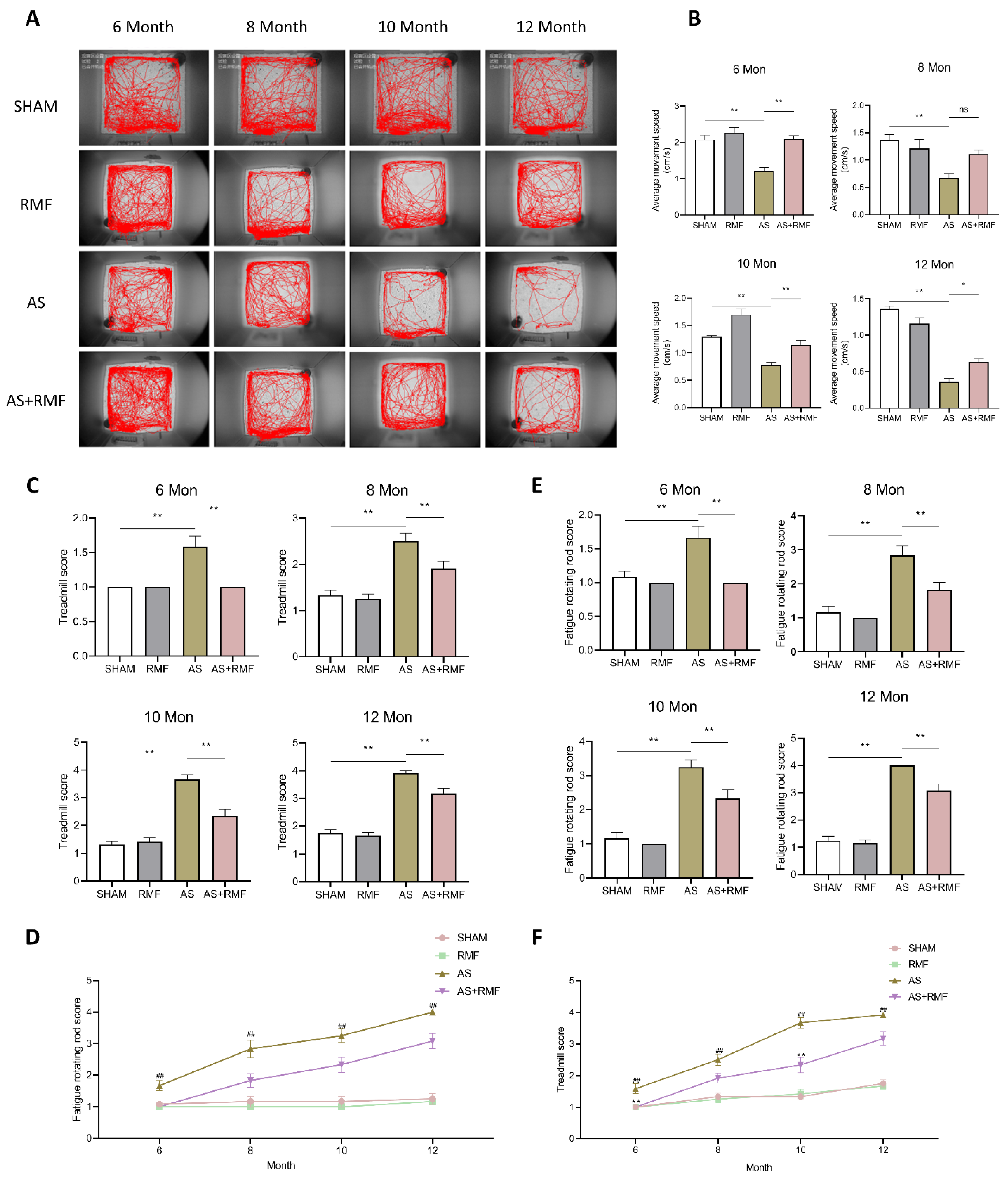
3.2. RMF Mitigates Athletic Injury and Age-Related Articular Deterioration in AS Mice
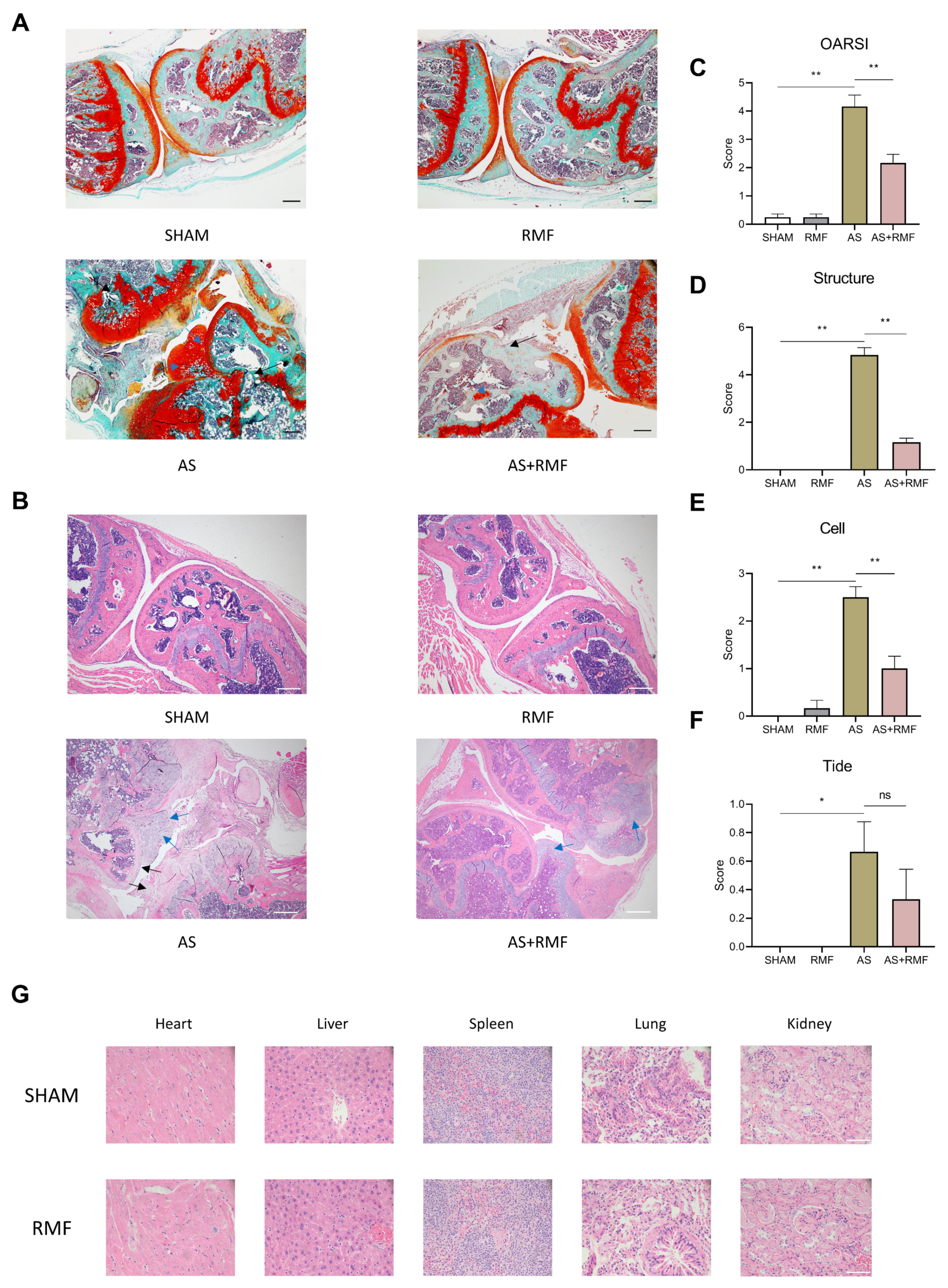
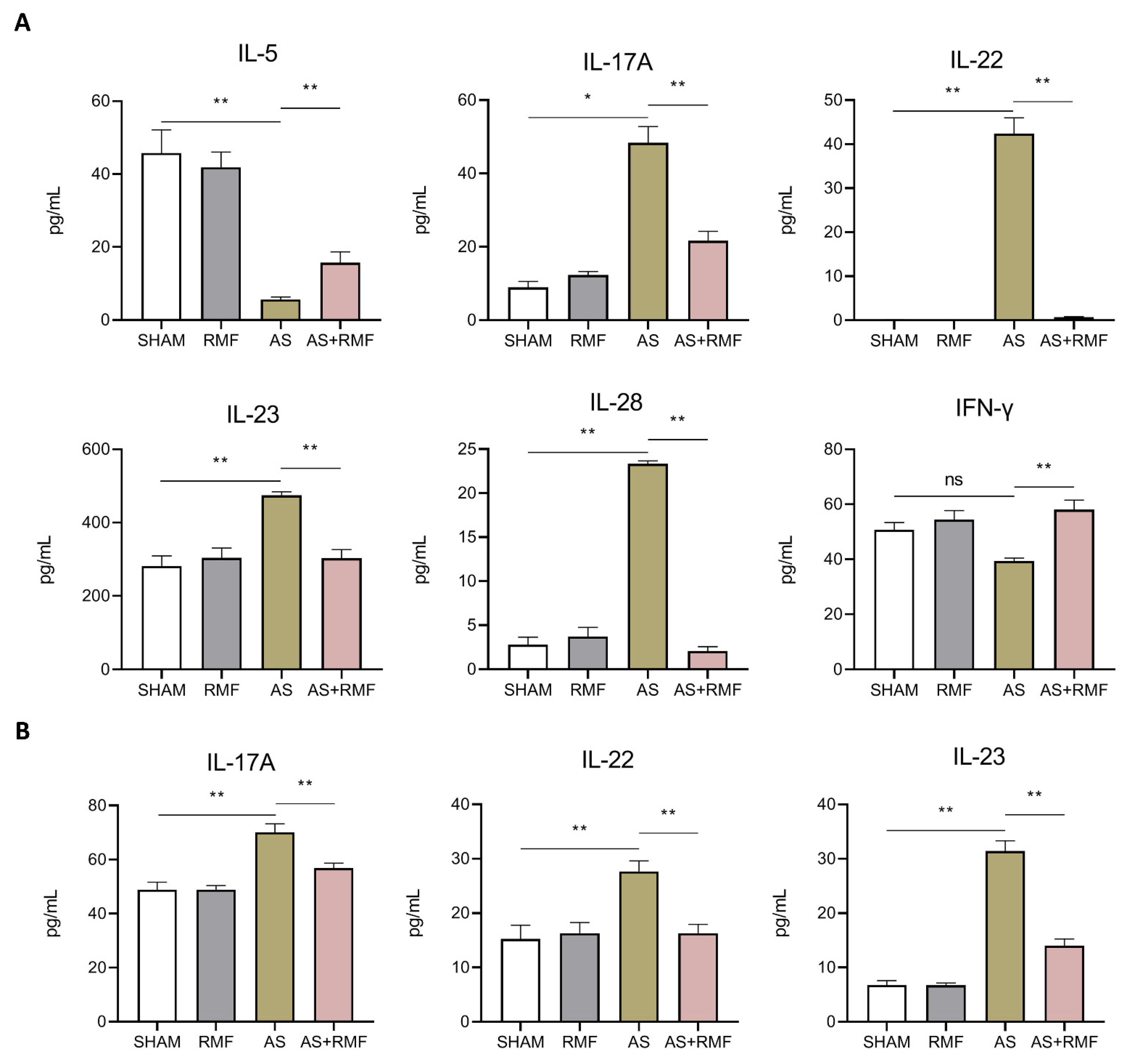
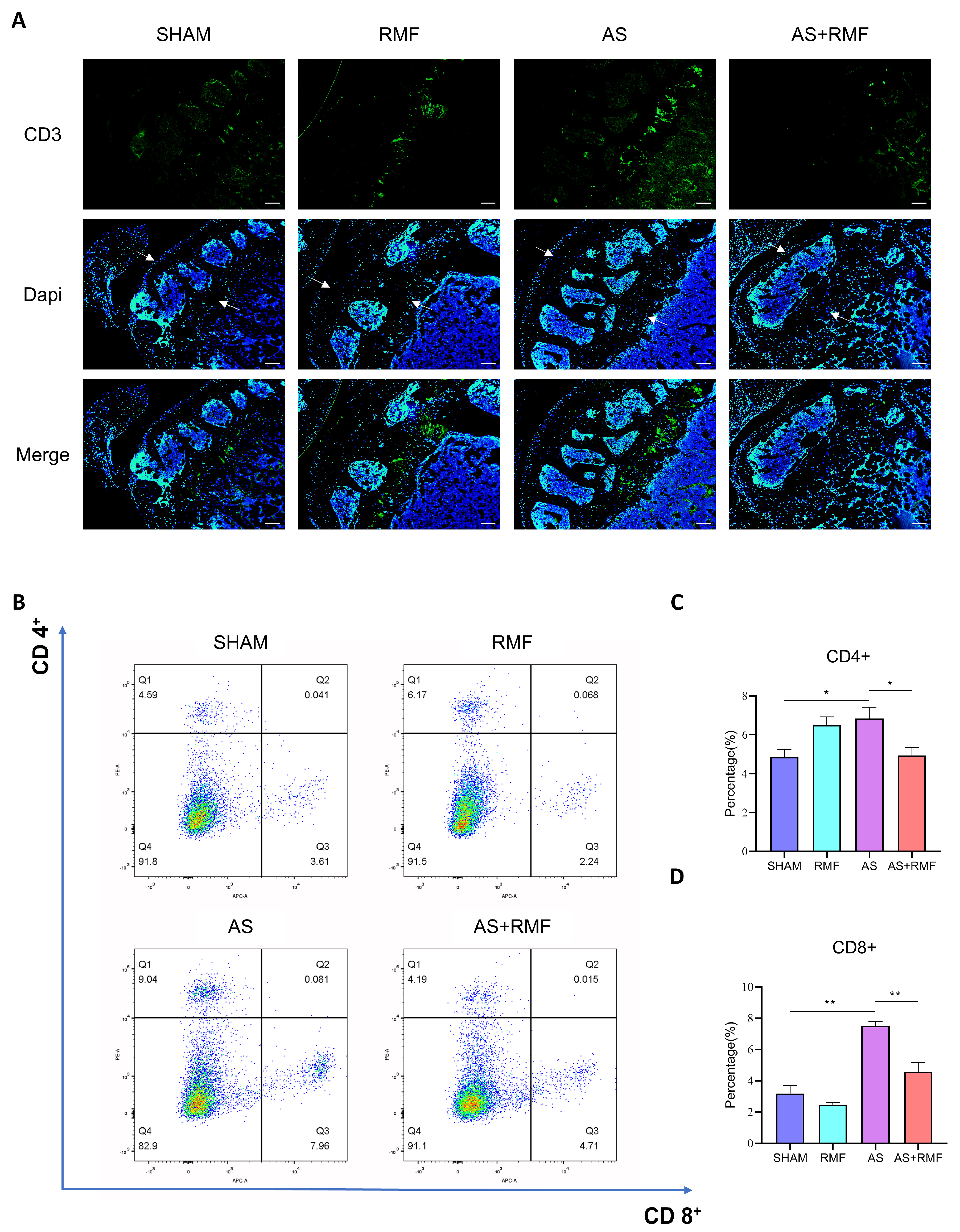
3.3. RMF Mitigates the Immune Dysfunction of AS Mice with Cartilage Deformation
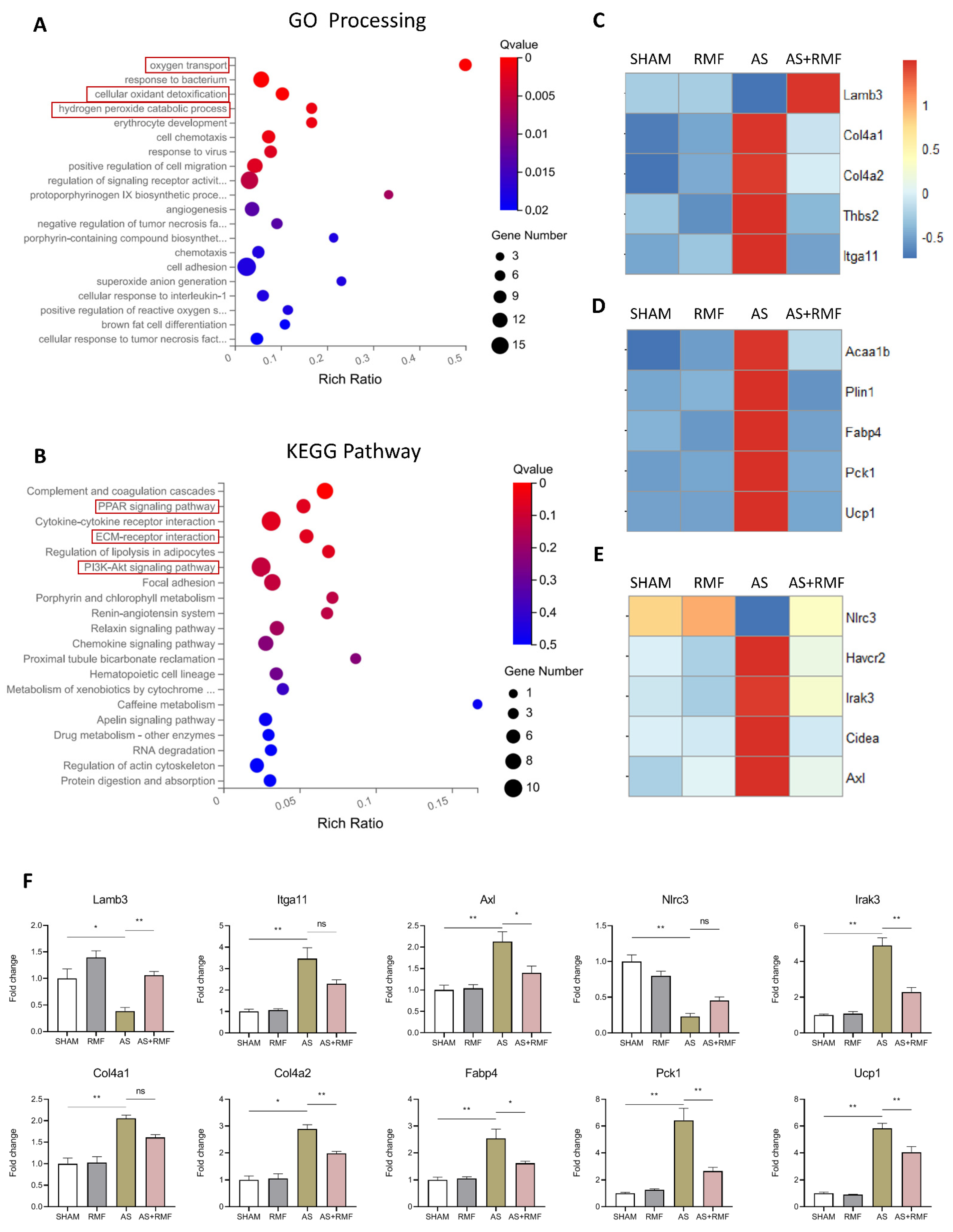
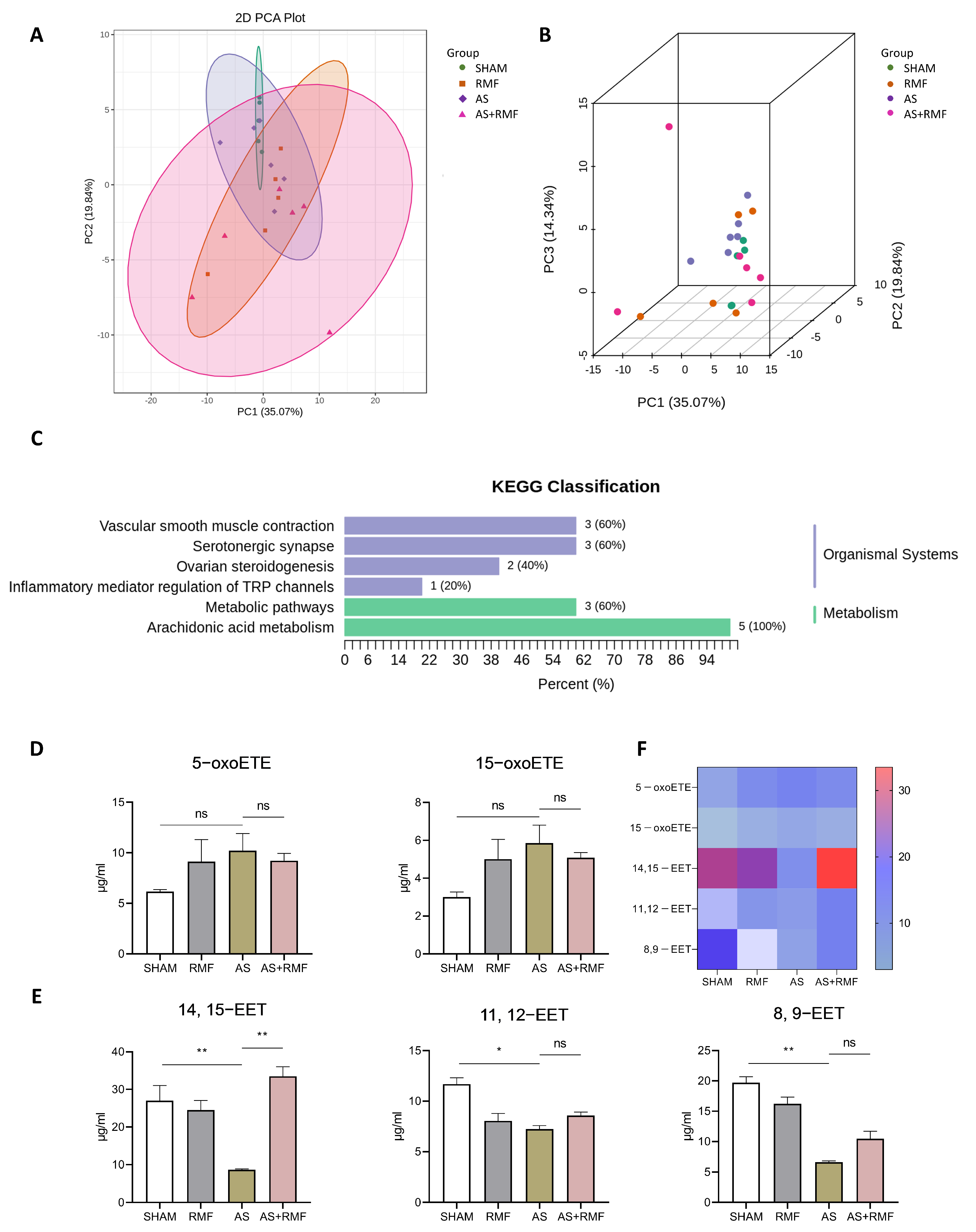
3.4. RMF Treatment Rebalances the Bone Redox System and Energy Metabolism of AS Mice
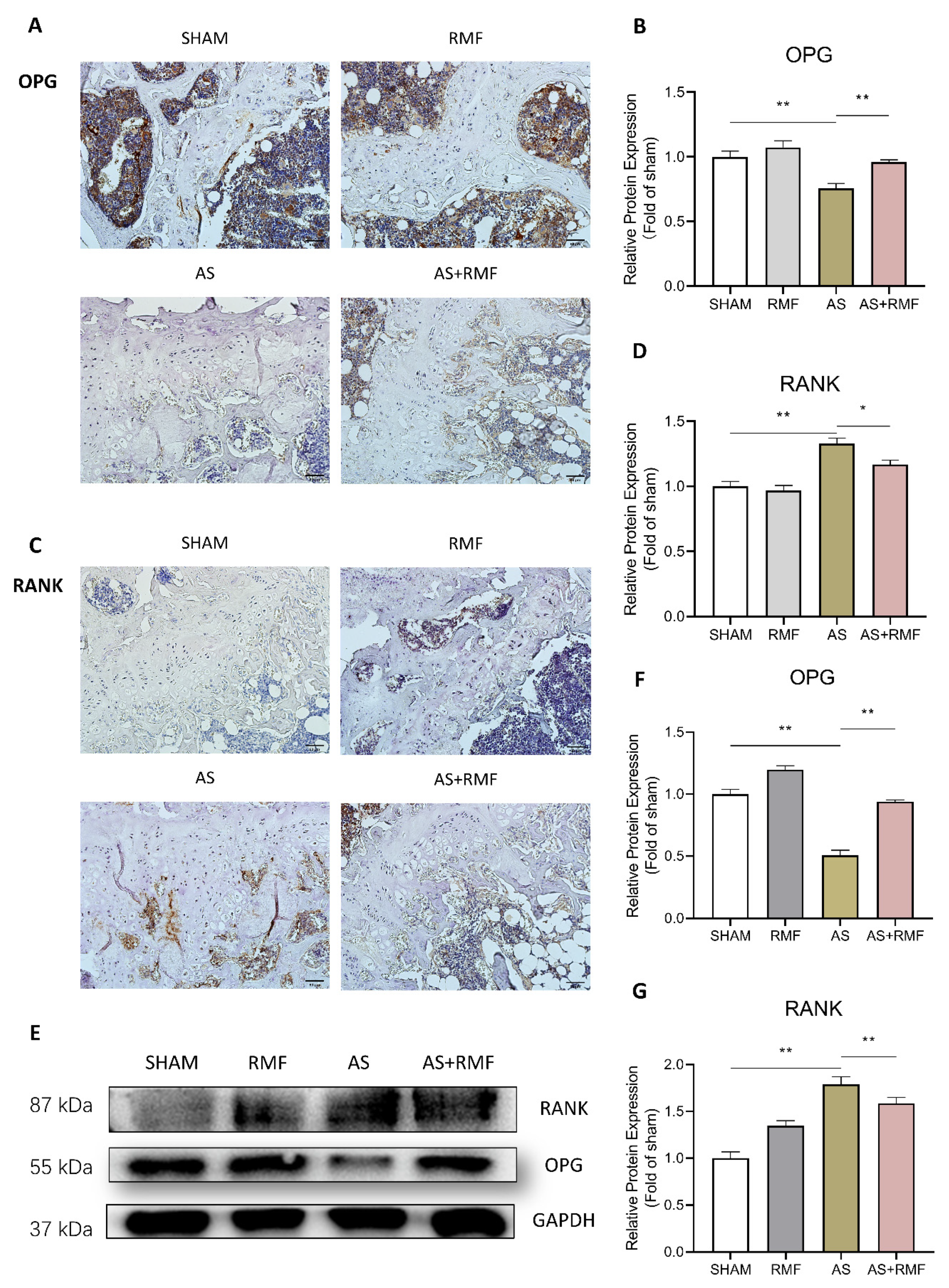
3.5. RMF Alleviates Bone and Cartilage Deterioration via the RANKL/RANK/OPG Signaling Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sieper, J.; Poddubnyy, D. Axial spondyloarthritis. LANCET 2017, 390, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Rudwaleit, M.; Landewé, R.; Sieper, J. Ankylosing Spondylitis and Axial Spondyloarthritis. N. Engl. J. Med. 2016, 375, 1302–1303. [Google Scholar] [CrossRef] [PubMed]
- Cortes, A.; Hadler, J.; Pointon, J.P.; Robinson, P.C.; Karaderi, T.; Leo, P.; Cremin, K.; Pryce, K.; Harris, J.; Lee, S.; et al. Identification of multiple risk variants for ankylosing spondylitis through high-density genotyping of immune-related loci. Nat. Genet. 2013, 45, 730–738. [Google Scholar] [CrossRef] [Green Version]
- de Vlam, K.; Lories, R.J.; Luyten, F.P. Mechanisms of pathologic new bone formation. Curr. Rheumatol. Rep. 2006, 8, 332–337. [Google Scholar] [CrossRef]
- Lindsey, R.C.; Aghajanian, P.; Mohan, S. Thyroid Hormone Signaling in the Development of the Endochondral Skeleton. Vitam. Horm. 2018, 106, 351–381. [Google Scholar] [CrossRef]
- Salhotra, A.; Shah, H.N.; Levi, B.; Longaker, M.T. Mechanisms of bone development and repair. Nat. Reviews. Mol. Cell Biol. 2020, 21, 696–711. [Google Scholar] [CrossRef]
- Hallett, S.A.; Ono, W.; Ono, N. Growth Plate Chondrocytes: Skeletal Development, Growth and Beyond. Int. J. Mol. Sci. 2019, 20, 6009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, C.K.; Seo, E.Y.; Chen, J.Y.; Lo, D.; McArdle, A.; Sinha, R.; Tevlin, R.; Seita, J.; Vincent-Tompkins, J.; Wearda, T.; et al. Identification and specification of the mouse skeletal stem cell. Cell 2015, 160, 285–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landewe, R.; Dougados, M.; Mielants, H.; van der Tempel, H.; van der Heijde, D. Physical function in ankylosing spondylitis is independently determined by both disease activity and radiographic damage of the spine. Ann. Rheum. Dis. 2009, 68, 863–867. [Google Scholar] [CrossRef]
- Ward, M.M.; Deodhar, A.; Akl, E.A.; Lui, A.; Ermann, J.; Gensler, L.S.; Smith, J.A.; Borenstein, D.; Hiratzka, J.; Weiss, P.F.; et al. American College of Rheumatology/Spondylitis Association of America/Spondyloarthritis Research and Treatment Network 2015 Recommendations for the Treatment of Ankylosing Spondylitis and Nonradiographic Axial Spondyloarthritis. Arthritis Rheumatol. 2016, 68, 282–298. [Google Scholar] [CrossRef] [Green Version]
- Baeten, D.; Sieper, J.; Braun, J.; Baraliakos, X.; Dougados, M.; Emery, P.; Deodhar, A.; Porter, B.; Martin, R.; Andersson, M.; et al. Secukinumab, an Interleukin-17A Inhibitor, in Ankylosing Spondylitis. N. Engl. J. Med. 2015, 373, 2534–2548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baraliakos, X.; Listing, J.; Brandt, J.; Zink, A.; Alten, R.; Burmester, G.; Gromnica-Ihle, E.; Kellner, H.; Schneider, M.; Sörensen, H.; et al. Clinical response to discontinuation of anti-TNF therapy in patients with ankylosing spondylitis after 3 years of continuous treatment with infliximab. Arthritis Res. Ther. 2005, 7, R439–R444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baraliakos, X.; Listing, J.; Rudwaleit, M.; Sieper, J.; Braun, J. The relationship between inflammation and new bone formation in patients with ankylosing spondylitis. Arthritis Res. Ther. 2008, 10, R104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherlock, J.P.; Joyce-Shaikh, B.; Turner, S.P.; Chao, C.C.; Sathe, M.; Grein, J.; Gorman, D.M.; Bowman, E.P.; McClanahan, T.K.; Yearley, J.H.; et al. IL-23 induces spondyloarthropathy by acting on ROR-γt+ CD3+CD4-CD8- entheseal resident T cells. Nat. Med. 2012, 18, 1069–1076. [Google Scholar] [CrossRef]
- Li, X.; Wang, J.; Zhan, Z.; Li, S.; Zheng, Z.; Wang, T.; Zhang, K.; Pan, H.; Li, Z.; Zhang, N.; et al. Inflammation Intensity-Dependent Expression of Osteoinductive Wnt Proteins Is Critical for Ectopic New Bone Formation in Ankylosing Spondylitis. Arthritis Rheumatol. 2018, 70, 1056–1070. [Google Scholar] [CrossRef]
- van der Heijde, D.; Machado, P.; Braun, J.; Hermann, K.G.; Baraliakos, X.; Hsu, B.; Baker, D.; Landewé, R. MRI inflammation at the vertebral unit only marginally predicts new syndesmophyte formation: A multilevel analysis in patients with ankylosing spondylitis. Ann. Rheum. Dis. 2012, 71, 369–373. [Google Scholar] [CrossRef]
- Chiowchanwisawakit, P.; Lambert, R.G.; Conner-Spady, B.; Maksymowych, W.P. Focal fat lesions at vertebral corners on magnetic resonance imaging predict the development of new syndesmophytes in ankylosing spondylitis. Arthritis Rheum. 2011, 63, 2215–2225. [Google Scholar] [CrossRef]
- Diarra, D.; Stolina, M.; Polzer, K.; Zwerina, J.; Ominsky, M.S.; Dwyer, D.; Korb, A.; Smolen, J.; Hoffmann, M.; Scheinecker, C.; et al. Dickkopf-1 is a master regulator of joint remodeling. Nat. Med. 2007, 13, 156–163. [Google Scholar] [CrossRef]
- Yu, T.; Zhang, J.; Zhu, W.; Wang, X.; Bai, Y.; Feng, B.; Zhuang, Q.; Han, C.; Wang, S.; Hu, Q.; et al. Chondrogenesis mediates progression of ankylosing spondylitis through heterotopic ossification. Bone Res. 2021, 9, 19. [Google Scholar] [CrossRef]
- Klingberg, E.; Lorentzon, M.; Mellström, D.; Geijer, M.; Göthlin, J.; Hilme, E.; Hedberg, M.; Carlsten, H.; Forsblad-d’Elia, H. Osteoporosis in ankylosing spondylitis - prevalence, risk factors and methods of assessment. Arthritis Res. Ther. 2012, 14, R108. [Google Scholar] [CrossRef] [Green Version]
- Toussirot, E.; Wendling, D. Osteoporosis in ankylosing spondylitis. Presse Med. 1996, 25, 720–724. [Google Scholar] [PubMed]
- Appel, H.; Kuhne, M.; Spiekermann, S.; Köhler, D.; Zacher, J.; Stein, H.; Sieper, J.; Loddenkemper, C. Immunohistochemical analysis of hip arthritis in ankylosing spondylitis: Evaluation of the bone-cartilage interface and subchondral bone marrow. Arthritis Rheum. 2006, 54, 1805–1813. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.N.; LaMarche, M.J.; Chan, H.M.; Fekkes, P.; Garcia-Fortanet, J.; Acker, M.G.; Antonakos, B.; Chen, C.H.; Chen, Z.; Cooke, V.G.; et al. Allosteric inhibition of SHP2 phosphatase inhibits cancers driven by receptor tyrosine kinases. Nature 2016, 535, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Saxton, T.M.; Henkemeyer, M.; Gasca, S.; Shen, R.; Rossi, D.J.; Shalaby, F.; Feng, G.S.; Pawson, T. Abnormal mesoderm patterning in mouse embryos mutant for the SH2 tyrosine phosphatase Shp-2. EMBO J. 1997, 16, 2352–2364. [Google Scholar] [CrossRef]
- Niogret, C.; Birchmeier, W.; Guarda, G. SHP-2 in Lymphocytes’ Cytokine and Inhibitory Receptor Signaling. Front. Immunol. 2019, 10, 2468. [Google Scholar] [CrossRef]
- Yokosuka, T.; Takamatsu, M.; Kobayashi-Imanishi, W.; Hashimoto-Tane, A.; Azuma, M.; Saito, T. Programmed cell death 1 forms negative costimulatory microclusters that directly inhibit T cell receptor signaling by recruiting phosphatase SHP2. J. Exp. Med. 2012, 209, 1201–1217. [Google Scholar] [CrossRef] [Green Version]
- Kan, C.; Yang, F.; Wang, S. SHP2-Mediated Signal Networks in Stem Cell Homeostasis and Dysfunction. Stem Cells Int. 2018, 2018, 8351374. [Google Scholar] [CrossRef] [PubMed]
- Chan, G.; Kalaitzidis, D.; Neel, B.G. The tyrosine phosphatase Shp2 (PTPN11) in cancer. Cancer Metastasis Rev. 2008, 27, 179–192. [Google Scholar] [CrossRef]
- Tartaglia, M.; Mehler, E.L.; Goldberg, R.; Zampino, G.; Brunner, H.G.; Kremer, H.; van der Burgt, I.; Crosby, A.H.; Ion, A.; Jeffery, S.; et al. Mutations in PTPN11, encoding the protein tyrosine phosphatase SHP-2, cause Noonan syndrome. Nat. Genet. 2001, 29, 465–468. [Google Scholar] [CrossRef]
- Bowen, M.E.; Boyden, E.D.; Holm, I.A.; Campos-Xavier, B.; Bonafé, L.; Superti-Furga, A.; Ikegawa, S.; Cormier-Daire, V.; Bovée, J.V.; Pansuriya, T.C.; et al. Loss-of-function mutations in PTPN11 cause metachondromatosis, but not Ollier disease or Maffucci syndrome. PLoS Genet. 2011, 7, e1002050. [Google Scholar] [CrossRef]
- Yang, W.; Wang, J.; Moore, D.C.; Liang, H.; Dooner, M.; Wu, Q.; Terek, R.; Chen, Q.; Ehrlich, M.G.; Quesenberry, P.J.; et al. Ptpn11 deletion in a novel progenitor causes metachondromatosis by inducing hedgehog signalling. Nature 2013, 499, 491–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, H.; Zhao, W.; Hou, J.; Zhang, Y.; Xie, Y.; Zheng, Y.; Xu, H.; Qian, C.; Zhou, J.; Yu, Y.; et al. SHP-2 phosphatase negatively regulates the TRIF adaptor protein-dependent type I interferon and proinflammatory cytokine production. Immunity 2006, 25, 919–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.K.; Feng, G.S.; Chen, D.; King, P.D.; Kamiya, N. Targeted disruption of Shp2 in chondrocytes leads to metachondromatosis with multiple cartilaginous protrusions. J. Bone Miner. Res. 2014, 29, 761–769. [Google Scholar] [CrossRef] [Green Version]
- Bowen, M.E.; Ayturk, U.M.; Kurek, K.C.; Yang, W.; Warman, M.L. SHP2 regulates chondrocyte terminal differentiation, growth plate architecture and skeletal cell fates. PLoS Genet. 2014, 10, e1004364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.K.; Aruwajoye, O.; Sucato, D.; Richards, B.S.; Feng, G.S.; Chen, D.; King, P.D.; Kamiya, N. Induction of SHP2 deficiency in chondrocytes causes severe scoliosis and kyphosis in mice. Spine 2013, 38, E1307–E1312. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Huang, J.; Moore, D.C.; Zuo, C.; Wu, Q.; Xie, L.; von der Mark, K.; Yuan, X.; Chen, D.; Warman, M.L.; et al. SHP2 Regulates the Osteogenic Fate of Growth Plate Hypertrophic Chondrocytes. Sci. Rep. 2017, 7, 12699. [Google Scholar] [CrossRef] [Green Version]
- Zuo, C.; Wang, L.; Kamalesh, R.M.; Bowen, M.E.; Moore, D.C.; Dooner, M.S.; Reginato, A.M.; Wu, Q.; Schorl, C.; Song, Y.; et al. SHP2 regulates skeletal cell fate by modifying SOX9 expression and transcriptional activity. Bone Res. 2018, 6, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miah, S.M.S.; Jayasuriya, C.T.; Salter, A.I.; Reilly, E.C.; Fugere, C.; Yang, W.; Chen, Q.; Brossay, L. Ptpn11 Deletion in CD4(+) Cells Does Not Affect T Cell Development and Functions but Causes Cartilage Tumors in a T Cell-Independent Manner. Front. Immunol. 2017, 8, 1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guittard, G.; Gallardo, D.L.; Li, W.; Melis, N.; Lui, J.C.; Kortum, R.L.; Shakarishvili, N.G.; Huh, S.; Baron, J.; Weigert, R.; et al. Unexpected Cartilage Phenotype in CD4-Cre-Conditional SOS-Deficient Mice. Front. Immunol. 2017, 8, 343. [Google Scholar] [CrossRef] [Green Version]
- Wehenkel, M.; Corr, M.; Guy, C.S.; Edwards, B.A.; Castellaw, A.H.; Calabrese, C.; Pagès, G.; Pouysségur, J.; Vogel, P.; McGargill, M.A. Extracellular Signal-Regulated Kinase Signaling in CD4-Expressing Cells Inhibits Osteochondromas. Front. Immunol. 2017, 8, 482. [Google Scholar] [CrossRef] [Green Version]
- Markov, M.S. Magnetic field therapy: A review. Electromagn. Biol. Med. 2007, 26, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Hubley, M.J.; Rosanske, R.C.; Moerland, T.S. Diffusion coefficients of ATP and creatine phosphate in isolated muscle: Pulsed gradient 31P NMR of small biological samples. NMR Biomed. 1995, 8, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Harden, R.N.; Remble, T.A.; Houle, T.T.; Long, J.F.; Markov, M.S.; Gallizzi, M.A. Prospective, randomized, single-blind, sham treatment-controlled study of the safety and efficacy of an electromagnetic field device for the treatment of chronic low back pain: A pilot study. Pain Pract. 2007, 7, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Heyers, D.; Zapka, M.; Hoffmeister, M.; Wild, J.M.; Mouritsen, H. Magnetic field changes activate the trigeminal brainstem complex in a migratory bird. Proc. Natl. Acad. Sci. USA 2010, 107, 9394–9399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanderstraeten, J.; Gailly, P.; Malkemper, E.P. Low-Light Dependence of the Magnetic Field Effect on Cryptochromes: Possible Relevance to Plant Ecology. Front. Plant Sci. 2018, 9, 121. [Google Scholar] [CrossRef] [Green Version]
- Struk, M.; Grygorcewicz, B.; Nawrotek, P.; Augustyniak, A.; Konopacki, M.; Kordas, M.; Rakoczy, R. Enhancing effect of 50 Hz rotating magnetic field on induction of Shiga toxin-converting lambdoid prophages. Microb. Pathog. 2017, 109, 4–7. [Google Scholar] [CrossRef]
- Du, L.; Fan, H.; Miao, H.; Zhao, G.; Hou, Y. Extremely Low Frequency Magnetic Fields Inhibit Adipogenesis of Human Mesenchymal Stem Cells. Bioelectromagnetics 2014, 35, 519–530. [Google Scholar] [CrossRef]
- Nie, Y.; Du, L.; Mou, Y.; Xu, Z.; Weng, L.; Du, Y.; Zhu, Y.; Hou, Y.; Wang, T. Effect of low frequency magnetic fields on melanoma: Tumor inhibition and immune modulation. BMC Cancer 2013, 13, 582. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.Y.; Xue, Y.; Zhang, Y. Effects of 0.4 T rotating magnetic field exposure on density, strength, calcium and metabolism of rat thigh bones. Bioelectromagnetics 2006, 27, 1–9. [Google Scholar] [CrossRef]
- Goraca, A.; Ciejka, E.; Piechota, A. Effects of extremely low frequency magnetic field on the parameters of oxidative stress in heart. J. Physiol. Pharmacol. 2010, 61, 333–338. [Google Scholar] [CrossRef]
- Sirav, B.; Kuzay, D.; Delen, K.; Oruc, S.; Ozer, C. Oxidative Effects of Electromagnetic Radiation on Liver, Lung, Kidney and Heart Tissues of Diabetic and Normal Rats. Gazi Med. J. 2021, 32, 397–402. [Google Scholar] [CrossRef]
- Zhu, J.; Yang, J.; Yang, X.; Li, Y.; Xiao, S.; Guo, Z.; Yu, M. Effects of Low Frequency Rotating Permanent Magnetic Field on Human Body Surface Thermal Distribution, Blood Pressure and Heart Rate. Space Med. Med. Eng. 2012, 25, 417–421. [Google Scholar] [CrossRef]
- Wu, X.; Guo, W.; Wu, L.; Gu, Y.; Gu, L.; Xu, S.; Wu, X.; Shen, Y.; Ke, Y.; Tan, R.; et al. Selective sequestration of STAT1 in the cytoplasm via phosphorylated SHP-2 ameliorates murine experimental colitis. J. Immunol. 2012, 189, 3497–3507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, F.; Liu, Q.; Zhu, Y.; Fan, Z.; Chen, W.; Liu, S.; Li, X.; Guo, W.; Feng, G.-S.; Yu, H.; et al. Targeting chondrocytes for arresting bony fusion in ankylosing spondylitis. Nat. Commun. 2021, 12, 6540. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Lin, G.M.; Wu, N.; Tang, S.W.; Zheng, Z.J.; Lin, M.C.; Xu, G.X.; Liu, H.; Deng, Y.Y.; Zhang, X.Y.; et al. Early exposure of rotating magnetic fields promotes central nervous regeneration in planarian Girardia sinensis. Bioelectromagnetics 2016, 37, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Glasson, S.S.; Chambers, M.G.; Van den Berg, W.B.; Little, C.B. The OARSI histopathology initiative - recommendations for histological assessments of osteoarthritis in the mouse. Osteoarthr. Cartil. 2010, 18, S17–S23. [Google Scholar] [CrossRef] [Green Version]
- Reilly, E.C.; Thompson, E.A.; Aspeslagh, S.; Wands, J.R.; Elewaut, D.; Brossay, L. Activated iNKT cells promote memory CD8+ T cell differentiation during viral infection. PloS ONE 2012, 7, e37991. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Abel, E.D.; Long, F.X. Glucose metabolism induced by Bmp signaling is essential for murine skeletal development. Nat. Commun. 2018, 9, 4831. [Google Scholar] [CrossRef] [Green Version]
- McNamara, J.T.; Huntington, K.E.; Borys, S.; Jayasuriya, C.T.; Brossay, L. SHP-2 deletion in CD4Cre expressing chondrocyte precursors leads to tumor development with wrist tropism. Sci. Rep. 2021, 11, 20006. [Google Scholar] [CrossRef]
- Zhang, S.; Fan, Z.; Ouyang, Z.; Sun, H.; Song, Y.; Yu, H.; Xu, Q.; Liu, S.; Sun, Y.; Shao, F. Purine metabolites promote ectopic new bone formation in ankylosing spondylitis. Int. Immunopharmacol. 2023, 116, 109810. [Google Scholar] [CrossRef]
- Qin, X.H.; Wu, Z.; Dong, J.H.; Zeng, Y.N.; Xiong, W.C.; Liu, C.; Wang, M.Y.; Zhu, M.Z.; Chen, W.J.; Zhang, Y.; et al. Liver Soluble Epoxide Hydrolase Regulates Behavioral and Cellular Effects of Chronic Stress. Cell Rep. 2019, 29, 3223–3234.e3226. [Google Scholar] [CrossRef] [Green Version]
- Magrey, M.N.; Khan, M.A. The Paradox of Bone Formation and Bone Loss in Ankylosing Spondylitis: Evolving New Concepts of Bone Formation and Future Trends in Management. Curr. Rheumatol. Rep. 2017, 19, 17. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, S.; Doyle, D.V.; Denton, A.; Rolfe, I.; McCloskey, E.V.; Spector, T.D. Bone mineral density and vertebral compression fracture rates in ankylosing spondylitis. Ann. Rheum. Dis. 1994, 53, 117–121. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Shang, P. A review of bioeffects of static magnetic field on rodent models. Prog. Biophys. Mol. Biol. 2014, 114, 14–24. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Ke, Y.; Zhang, E.E.; Feng, G.S. Conditional deletion of Shp2 tyrosine phosphatase in thymocytes suppresses both pre-TCR and TCR signals. J. Immunol. 2006, 177, 5990–5996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labusca, L.; Herea, D.D.; Danceanu, C.M.; Minuti, A.E.; Stavila, C.; Grigoras, M.; Gherca, D.; Stoian, G.; Ababei, G.; Chiriac, H.; et al. The effect of magnetic field exposure on differentiation of magnetite nanoparticle-loaded adipose-derived stem cells. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 109, 110652. [Google Scholar] [CrossRef]
- Pan, X.H.; Xiao, D.M.; Zhang, X.Y.; Huang, Y.S.; Lin, B.W. Study of rotating permanent magnetic field to treat steroid-induced osteonecrosis of femoral head. Int. Orthop. 2009, 33, 617–623. [Google Scholar] [CrossRef] [Green Version]
- Gregorczyk, I.; Jasiecka-Mikolajczyk, A.; Maslanka, T. Blockade of NF-kappa B Translocation and of RANKL/RANK Interaction Decreases the Frequency of Th2 and Th17 Cells Capable of IL-4 and IL-17 Production, Respectively, in a Mouse Model of Allergic Asthma. Molecules 2021, 26, 3117. [Google Scholar] [CrossRef]
- Xiao, Y.; Huang, Y.; Chen, Z.Y. Distribution, depletion and recovery of docosahexaenoic acid are region-specific in rat brain. Br. J. Nutr. 2005, 94, 544–550. [Google Scholar] [CrossRef] [Green Version]
- Spector, A.A. Arachidonic acid cytochrome P450 epoxygenase pathway. J. Lipid Res. 2009, 50, S52–S56. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Hao, J.Q.; Zeng, J.; Sauter, E.R. SnapShot: FABP Functions. Cell 2020, 182, 1066. [Google Scholar] [CrossRef]
- Wan, Y.; Chong, L.-W.; Evans, R.M. PPAR-gamma regulates osteoclastogenesis in mice. Nat. Med. 2007, 13, 1496–1503. [Google Scholar] [CrossRef]
- Kasonga, A.; Kruger, M.C.; Coetzee, M. Activation of PPARs Modulates Signalling Pathways and Expression of Regulatory Genes in Osteoclasts Derived from Human CD14+Monocytes. Int. J. Mol. Sci. 2019, 20, 1798. [Google Scholar] [CrossRef] [Green Version]
- Hynes, R.O. Integrins: Bidirectional, allosteric signaling machines. Cell 2002, 110, 673–687. [Google Scholar] [CrossRef] [Green Version]
- Robinson, W.H.; Lepus, C.M.; Wang, Q.; Raghu, H.; Mao, R.; Lindstrom, T.M.; Sokolove, J. Low-grade inflammation as a key mediator of the pathogenesis of osteoarthritis. Nat. Rev. Rheumatol. 2016, 12, 580–592. [Google Scholar] [CrossRef]
- Lin, D.P.; Li, L.; Sun, Y.; Wang, W.D.; Wang, X.Q.; Ye, Y.; Chen, X.; Xu, Y. Interleukin-17 regulates the expressions of RANKL and OPG in human periodontal ligament cells via TRAF6/TBK1-JNK/NF-kappa B pathways. Immunology 2015, 144, 472–485. [Google Scholar] [CrossRef]
- Dudakov, J.A.; Hanash, A.M.; van den Brink, M.R.M. Interleukin-22: Immunobiology and Pathology. Annu. Rev. Immunol. 2015, 33, 747–785. [Google Scholar] [CrossRef] [Green Version]
- Riol-Blanco, L.; Ordovas-Montanes, J.; Perro, M.; Naval, E.; Thiriot, A.; Alvarez, D.; Paust, S.; Wood, J.N.; von Andrian, U.H. Nociceptive sensory neurons drive interleukin-23-mediated psoriasiform skin inflammation. Nature 2014, 510, 157–161. [Google Scholar] [CrossRef] [Green Version]
- Ono, T.; Hayashi, M.; Sasaki, F.; Nakashima, T. RANKL biology: Bone metabolism, the immune system, and beyond. Inflamm. Regen. 2020, 40, 2. [Google Scholar] [CrossRef] [Green Version]
- Alippe, Y.; Mbalaviele, G. Omnipresence of inflammasome activities in inflammatory bone diseases. Semin. Immunopathol. 2019, 41, 607–618. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.Y.; Xu, J.Q.; Zhang, W.Y.; Song, C.Y.; Gao, C.G.; He, Y.J.; Shang, Y. Negative regulator NLRC3: Its potential role and regulatory mechanism in immune response and immune-related diseases. Front. Immunol. 2022, 13, 1012459. [Google Scholar] [CrossRef] [PubMed]
- Uchimura, T.; Oyama, Y.; Deng, M.; Guo, H.; Wilson, J.E.; Rampanelli, E.; Cook, K.D.; Misumi, I.; Tan, X.; Chen, L.; et al. The Innate Immune Sensor NLRC3 Acts as a Rheostat that Fine-Tunes T Cell Responses in Infection and Autoimmunity. Immunity 2018, 49, 1049–1061. [Google Scholar] [CrossRef]
- Tao, T.; Zhang, Y.K.; Wei, H.; Heng, K. Downregulation of IRAK3 by miR-33b-3p relieves chondrocyte inflammation and apoptosis in an in vitro osteoarthritis model. Biosci. Biotechnol. Biochem. 2021, 85, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Chinju, A.; Moriyama, M.; Kakizoe-Ishiguro, N.; Chen, H.; Miyahara, Y.; Haque, A.; Furusho, K.; Sakamoto, M.; Kai, K.; Kibe, K.; et al. CD163+M2 Macrophages Promote Fibrosis in IgG4-Related Disease Via Toll-Like Receptor 7/Interleukin-1 Receptor-Associated Kinase 4/NF-kappa B Signaling. Arthritis Rheumatol. 2022, 74, 892–901. [Google Scholar] [CrossRef]
- Gomes da Silva, I.I.F.; Barbosa, A.D.; Souto, F.O.; Maia, M.M.D.; Crovella, S.; Souza, P.R.E.; Sandrin-Garcia, P. MYD88, IRAK3 and Rheumatoid Arthritis pathogenesis: Analysis of differential gene expression in CD14 + monocytes and the inflammatory cytokine levels. Immunobiology 2021, 226, 152152. [Google Scholar] [CrossRef]
- Yu, J.; Qin, Y.; Zhou, N. Knockdown of Circ_SLC39A8 protects against the progression of osteoarthritis by regulating miR-591/IRAK3 axis. J. Orthop. Surg. Res. 2021, 16, 170. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score | Description |
|---|---|
| 0 | None |
| 1 | Enlarged pelvic incidence angle |
| 2 | Slight stiffness of knee joints |
| 3 | Scoliosis, kyphosis, and moderate stiffness of knee joints |
| 4 | Marked scoliosis and kyphosis, ankylosis of knee and hip joints |
| 5 | Ankylosis of the lower part of the body and spine |
| Score | Description |
|---|---|
| 1 | Normal movement |
| 2 | Mild dyskinesia |
| 3 | Inferior athletic capacity |
| 4 | Severe dyskinesia or motor incapacity |
| Score | Crawling Time |
|---|---|
| 1 | ≥5 min |
| 1.5 | 4.5–5 min |
| 2 | 4–4.5 min |
| 2.5 | 3.5–4 min |
| 3 | 3–3.5 min |
| 3.5 | 2.5–3 min |
| 4 | <2.5 min |
| Score | Number of Electric Shocks |
|---|---|
| 1 | ≤2 |
| 1.5 | 3–5 |
| 2 | 6–8 |
| 2.5 | 9–11 |
| 3 | 12–14 |
| 3.5 | 15–17 |
| 4 | >17 |
| Primer | Sequence |
|---|---|
| β-actin-F | AATCGTGCGTGACATCAAA |
| β-actin-R | ATGGATGCCACAGGATTCCATACCC |
| Lamb3-F | TTCGCTGCCTGACTTGACA |
| Lamb3-R | AAGCGACGGACTGAACTGT |
| Itga11-F | CACGGCATTTGGCATTGAA |
| Itga11-R | GTGCCGTAAACCGTAACTT |
| Axl-F | GAGCCAACCGTGGAAAGAG |
| Axl-R | CTCGGTTGGCACCTTTCTC |
| Nlrc3-F | CAGGCGGAGCCCTTTAGCA |
| Nlrc3-R | GTCCGCCTCGGGAAATCGT |
| Irak3-F | AAGACCCACGATGGACGAA |
| Irak3-R | AGAGGTCCAGGGTCGTTTT |
| Col4a1-F | GGACAAATCGGACCCACTG |
| Col4a1-R | CCTGTTTAGCCTGGGTGAC |
| Col4a2-F | CCTCCAGGACAGGGCTTAC |
| Col4a2-R | GGAGGTCCTGTCCCGAATG |
| Fabp4-F | AGTGGGAGTGGGCTTTGCC |
| Fabp4-R | TCCTGTCGTCTGCGGTGAT |
| Pck1-F | CAGTCATCATCACCCAAGAG |
| Pck1-R | ATAGGGCGAGTCTGTCAGTT |
| Ucp1-F | GTACCGAGCTGTGCGATGT |
| Ucp1-R | TTCCTCCAAGTTGCTTATGTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Y.; Yang, H.; Hua, Z.; Nie, S.; Xu, S.; Zhou, C.; Chen, F.; Li, M.; Yu, Q.; Sun, Y.; et al. Rotating Magnetic Field Mitigates Ankylosing Spondylitis Targeting Osteocytes and Chondrocytes via Ameliorating Immune Dysfunctions. Cells 2023, 12, 972. https://doi.org/10.3390/cells12070972
Han Y, Yang H, Hua Z, Nie S, Xu S, Zhou C, Chen F, Li M, Yu Q, Sun Y, et al. Rotating Magnetic Field Mitigates Ankylosing Spondylitis Targeting Osteocytes and Chondrocytes via Ameliorating Immune Dysfunctions. Cells. 2023; 12(7):972. https://doi.org/10.3390/cells12070972
Chicago/Turabian StyleHan, Yu, Hua Yang, Zhongke Hua, Shenglan Nie, Shuling Xu, Cai Zhou, Fengyi Chen, Mengqing Li, Qinyao Yu, Yang Sun, and et al. 2023. "Rotating Magnetic Field Mitigates Ankylosing Spondylitis Targeting Osteocytes and Chondrocytes via Ameliorating Immune Dysfunctions" Cells 12, no. 7: 972. https://doi.org/10.3390/cells12070972