
Proteomic Landscape of Human Sperm in Patients with Different Spermatogenic Impairments
 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
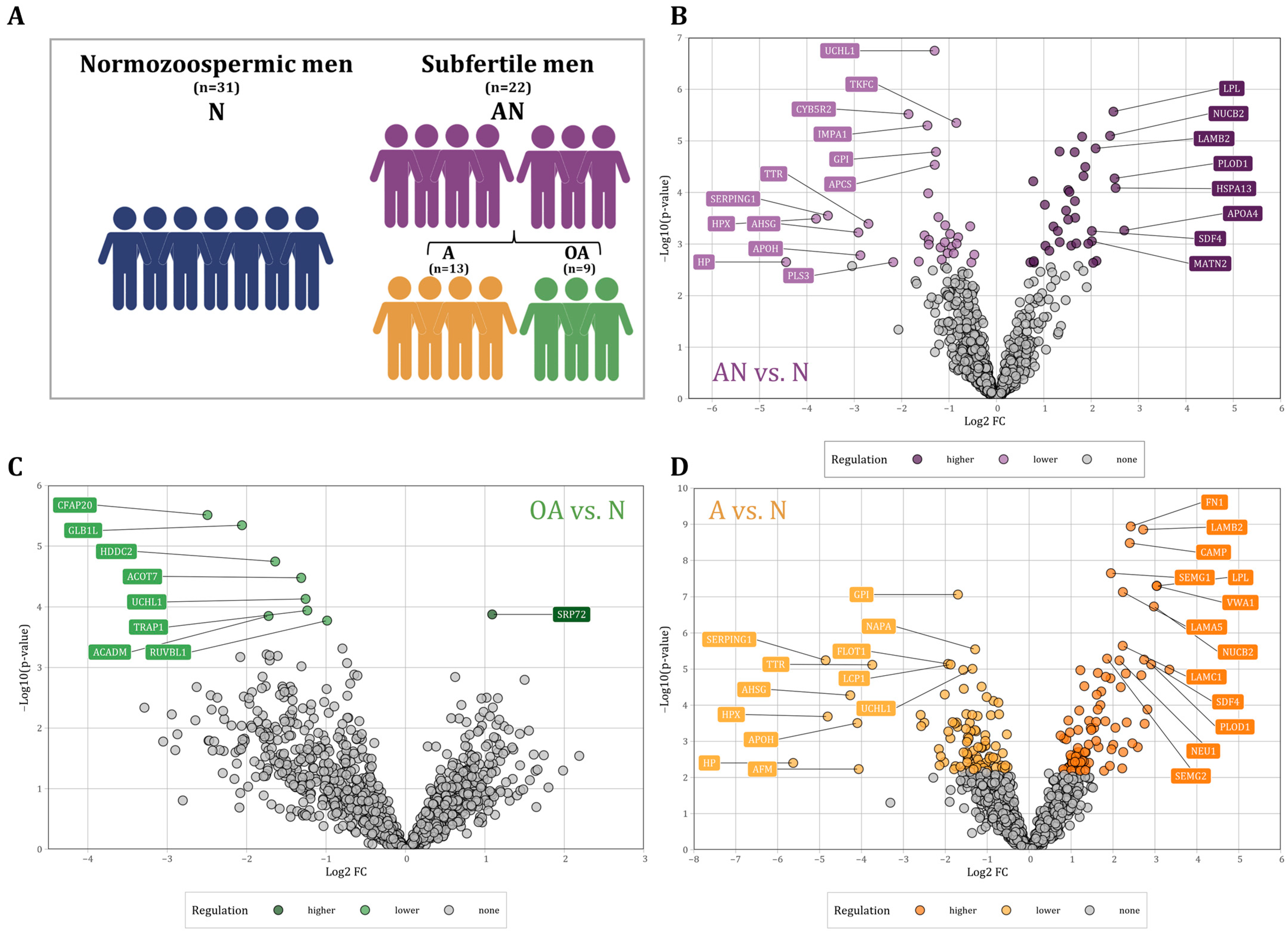
2.1. Study Population and Sample Collection
2.2. Protein Lysis
2.3. Peptide Preparation and LC-MS/MS Analysis
2.4. Data Processing and Statistical Analysis
3. Results
3.1. Basic Semen Characteristics of Patients with Subfertility Compared to Normozoospermic Men
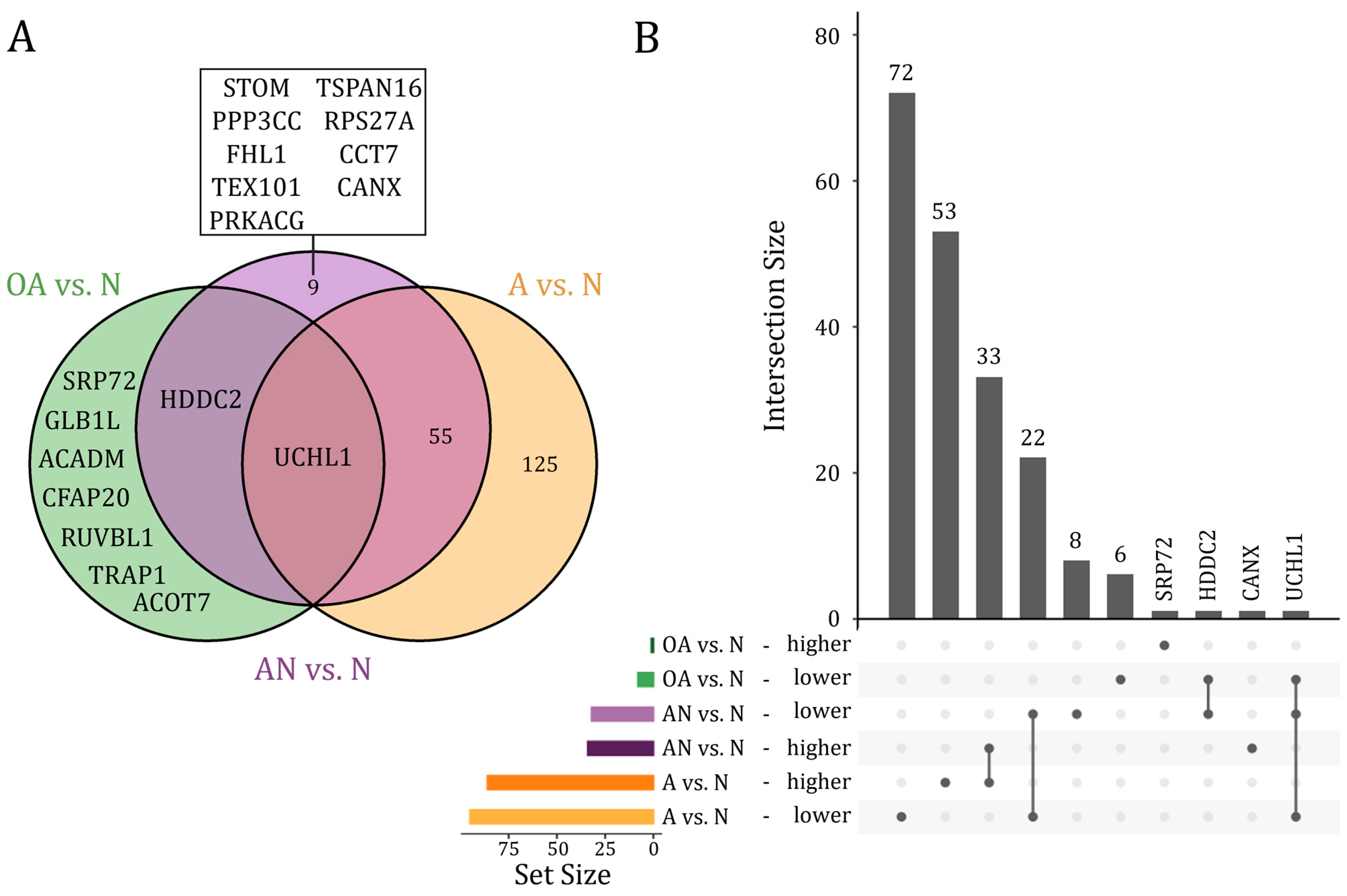
3.2. Differentially Abundant Proteins in Sperm as Determined by LC-MS/MS
3.3. Dysregulated Proteins between Subfertile and Normozoospermic Men
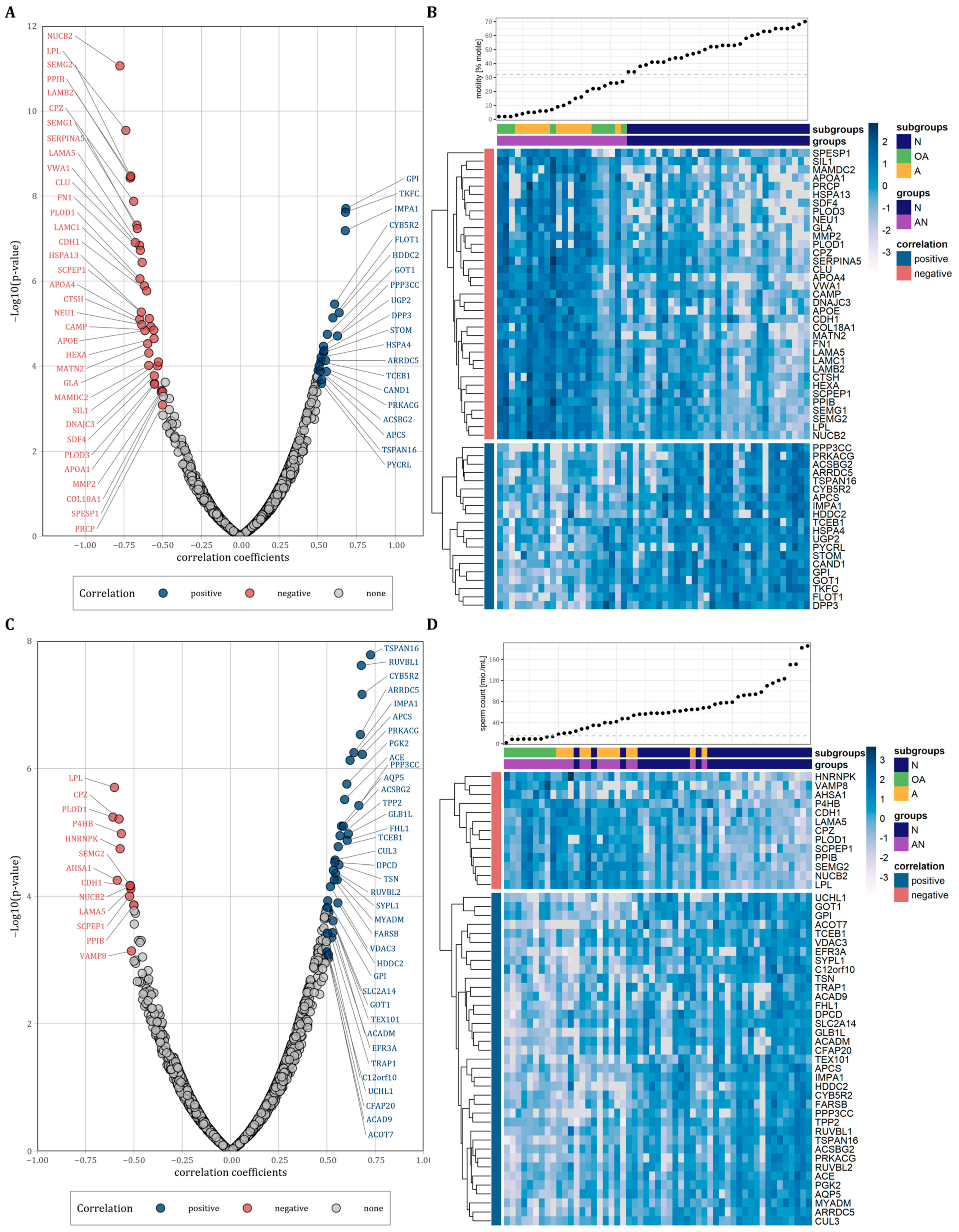
3.4. Correlation of Proteins with Progressive Sperm Motility and Sperm Count
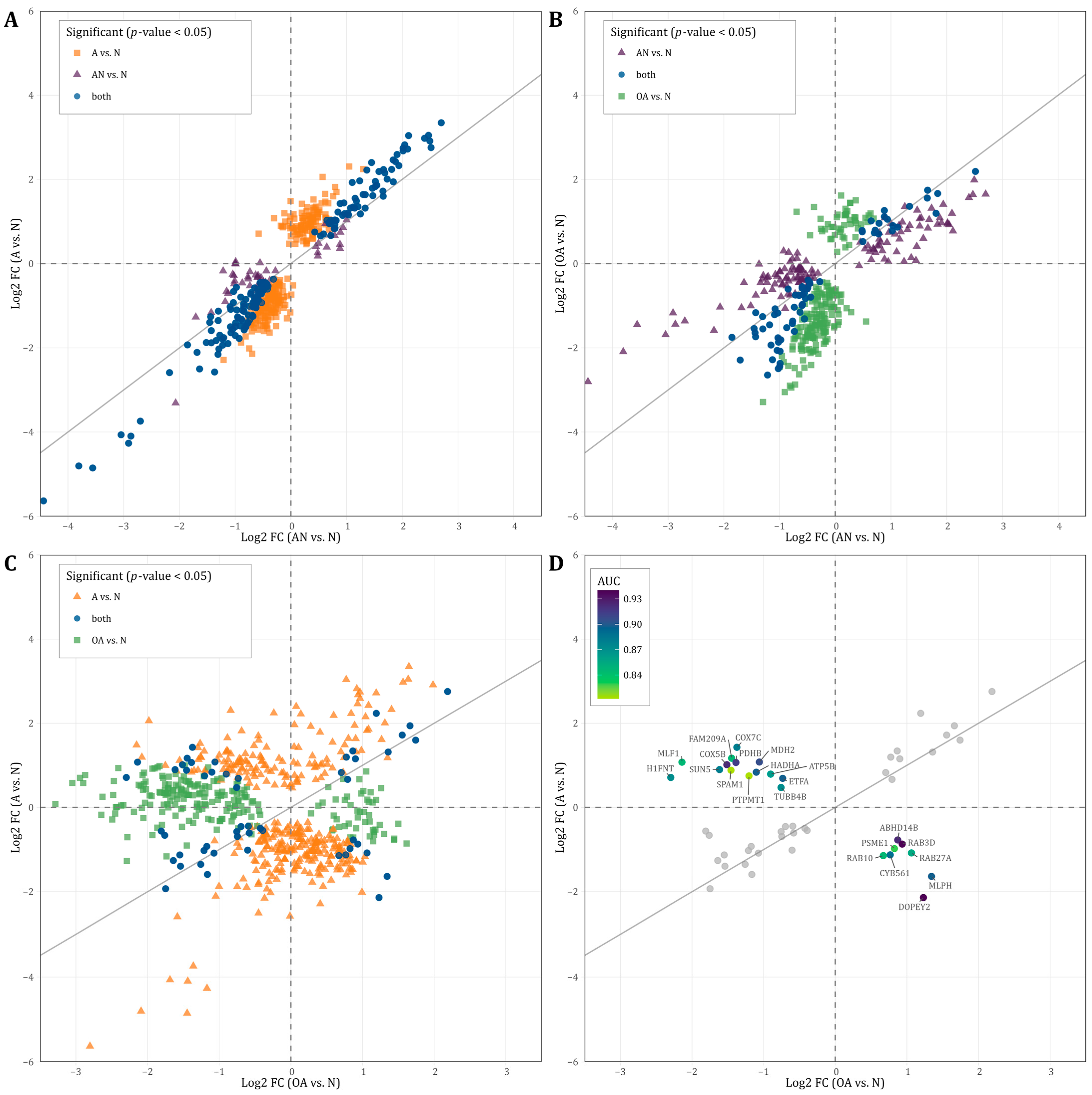
3.5. Directions of Deregulation and Evidence for Diagnostic Accuracy
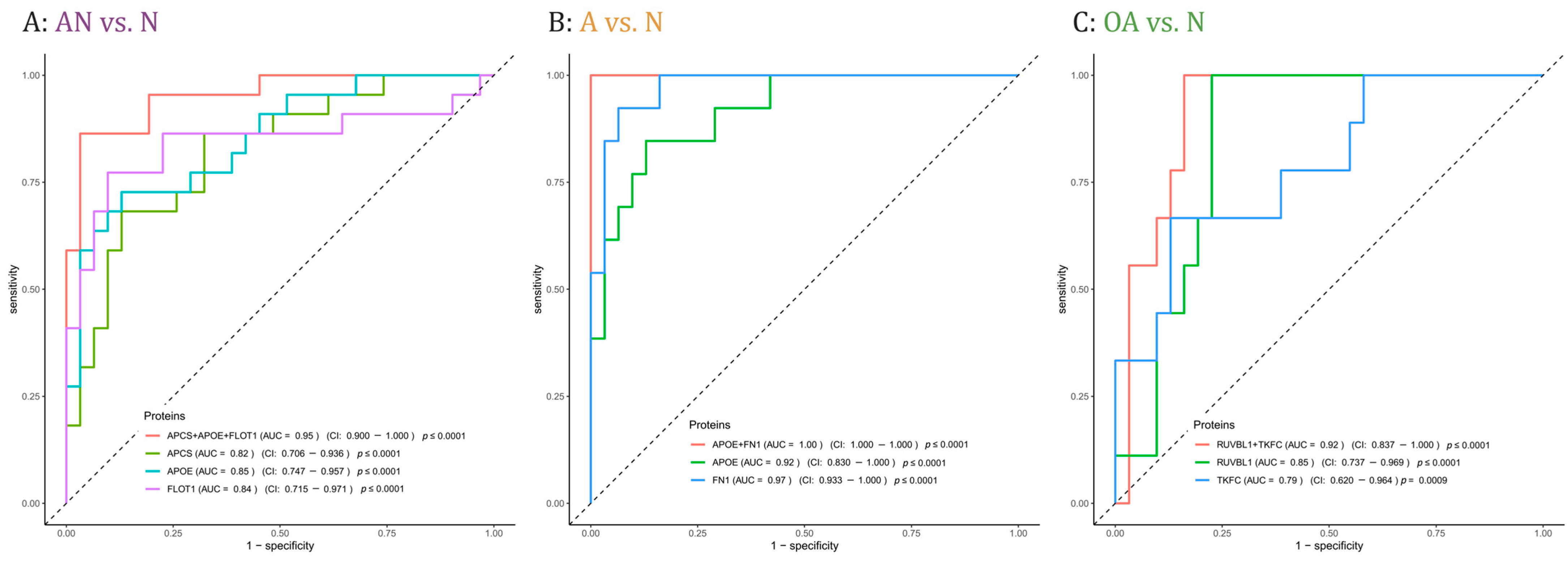
3.6. Prediction of Subfertility and Their Specific Phenotypes Oligoasthenozoospermia and Asthenozoospermia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jungwirth, A.; Diemer, T.; Kopa, Z.; Krausz, C.; Minhas, S.; Tournaye, H. EAU Guidelines on male fertility. In Proceedings of the EAU Annual Congress Barcelona, Barcelona, Spain, 15–19 March 2019. [Google Scholar]
- Abu-Halima, M.; Ayesh, B.M.; Hart, M.; Alles, J.; Fischer, U.; Hammadeh, M.; Keller, A.; Huleihel, M.; Meese, E. Differential expression of miR-23a/b-3p and its target genes in male patients with subfertility. Fertil. Steril. 2019, 112, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Abu-Halima, M.; Becker, L.S.; Al Smadi, M.A.; Kunz, L.S.; Groger, L.; Meese, E. Expression of SPAG7 and its regulatory microRNAs in seminal plasma and seminal plasma-derived extracellular vesicles of patients with subfertility. Sci. Rep. 2023, 13, 3645. [Google Scholar] [CrossRef] [PubMed]
- Abu-Halima, M.; Becker, L.S.; Ayesh, B.M.; Meese, E. MicroRNA-targeting in male infertility: Sperm microRNA-19a/b-3p and its spermatogenesis related transcripts content in men with oligoasthenozoospermia. Front. Cell Dev. Biol. 2022, 10, 973849. [Google Scholar] [CrossRef] [PubMed]
- Abu-Halima, M.; Belkacemi, A.; Ayesh, B.M.; Simone Becker, L.; Sindiani, A.M.; Fischer, U.; Hammadeh, M.; Keller, A.; Meese, E. MicroRNA-targeting in spermatogenesis: Over-expressions of microRNA-23a/b-3p and its affected targeting of the genes ODF2 and UBQLN3 in spermatozoa of patients with oligoasthenozoospermia. Andrology 2021, 9, 1137–1144. [Google Scholar] [CrossRef]
- Joshi, M.; Andrabi, S.W.; Singh, V.; Bansal, S.K.; Makker, G.C.; Mishra, G.; Gupta, G.; Rajender, S. Coding and regulatory transcriptome comparisons between fertile and infertile spermatozoa identify RNA signatures of male infertility. Andrologia 2022, 54, e14437. [Google Scholar] [CrossRef]
- Castillo, J.; Jodar, M.; Oliva, R. The contribution of human sperm proteins to the development and epigenome of the preimplantation embryo. Hum. Reprod. Update 2018, 24, 535–555. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Heredia, J.; de Mateo, S.; Vidal-Taboada, J.M.; Ballesca, J.L.; Oliva, R. Identification of proteomic differences in asthenozoospermic sperm samples. Hum. Reprod. 2008, 23, 783–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowicka-Bauer, K.; Lepczynski, A.; Ozgo, M.; Kamieniczna, M.; Fraczek, M.; Stanski, L.; Olszewska, M.; Malcher, A.; Skrzypczak, W.; Kurpisz, M.K. Sperm mitochondrial dysfunction and oxidative stress as possible reasons for isolated asthenozoospermia. J. Physiol. Pharmacol. 2018, 69. [Google Scholar] [CrossRef]
- Parte, P.P.; Rao, P.; Redij, S.; Lobo, V.; D’Souza, S.J.; Gajbhiye, R.; Kulkarni, V. Sperm phosphoproteome profiling by ultra performance liquid chromatography followed by data independent analysis (LC-MS(E)) reveals altered proteomic signatures in asthenozoospermia. J. Proteom. 2012, 75, 5861–5871. [Google Scholar] [CrossRef]
- Saraswat, M.; Joenvaara, S.; Jain, T.; Tomar, A.K.; Sinha, A.; Singh, S.; Yadav, S.; Renkonen, R. Human Spermatozoa Quantitative Proteomic Signature Classifies Normo- and Asthenozoospermia. Mol. Cell. Proteom. 2017, 16, 57–72. [Google Scholar] [CrossRef] [Green Version]
- Sinha, A.; Singh, V.; Singh, S.; Yadav, S. Proteomic analyses reveal lower expression of TEX40 and ATP6V0A2 proteins related to calcium ion entry and acrosomal acidification in asthenozoospermic males. Life Sci. 2019, 218, 81–88. [Google Scholar] [CrossRef]
- Siva, A.B.; Kameshwari, D.B.; Singh, V.; Pavani, K.; Sundaram, C.S.; Rangaraj, N.; Deenadayal, M.; Shivaji, S. Proteomics-based study on asthenozoospermia: Differential expression of proteasome alpha complex. Mol. Hum. Reprod. 2010, 16, 452–462. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Huo, R.; Wang, F.Q.; Lin, M.; Zhou, Z.M.; Sha, J.H. Identification of several proteins involved in regulation of sperm motility by proteomic analysis. Fertil. Steril. 2007, 87, 436–438. [Google Scholar] [CrossRef]
- Agarwal, A.; Panner Selvam, M.K.; Baskaran, S. Proteomic Analyses of Human Sperm Cells: Understanding the Role of Proteins and Molecular Pathways Affecting Male Reproductive Health. Int. J. Mol. Sci. 2020, 21, 1621. [Google Scholar] [CrossRef] [Green Version]
- Hashemitabar, M.; Sabbagh, S.; Orazizadeh, M.; Ghadiri, A.; Bahmanzadeh, M. A proteomic analysis on human sperm tail: Comparison between normozoospermia and asthenozoospermia. J. Assist. Reprod. Genet. 2015, 32, 853–863. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Cui, Y.; Zhang, X.; Lou, J.; Zhou, J.; Bei, H.; Wei, R. Proteomic profile of human spermatozoa in healthy and asthenozoospermic individuals. Reprod. Biol. Endocrinol. 2018, 16, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gahlay, G.K.; Rajput, N. The enigmatic sperm proteins in mammalian fertilization: An overviewdagger. Biol. Reprod. 2020, 103, 1171–1185. [Google Scholar] [CrossRef] [PubMed]
- Abu-Halima, M.; Hammadeh, M.; Backes, C.; Fischer, U.; Leidinger, P.; Lubbad, A.M.; Keller, A.; Meese, E. Panel of five microRNAs as potential biomarkers for the diagnosis and assessment of male infertility. Fertil. Steril. 2014, 102, 989–997.e1. [Google Scholar] [CrossRef]
- Abu-Halima, M.; Hammadeh, M.; Schmitt, J.; Leidinger, P.; Keller, A.; Meese, E.; Backes, C. Altered microRNA expression profiles of human spermatozoa in patients with different spermatogenic impairments. Fertil. Steril. 2013, 99, 1249–1255.e16. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Rappsilber, J.; Mann, M.; Ishihama, Y. Protocol for micro-purification, enrichment, pre-fractionation and storage of peptides for proteomics using StageTips. Nat. Protoc. 2007, 2, 1896–1906. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell. Proteom. 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; Garcia-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate—A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Burton, J.; Wojewodzic, M.W.; Rounge, T.B.; Haugen, T.B. A Role of the TEX101 Interactome in the Common Aetiology Behind Male Subfertility and Testicular Germ Cell Tumor. Front. Oncol. 2022, 12, 892043. [Google Scholar] [CrossRef]
- Endo, S.; Yoshitake, H.; Tsukamoto, H.; Matsuura, H.; Kato, K.; Sakuraba, M.; Takamori, K.; Fujiwara, H.; Takeda, S.; Araki, Y. TEX101, a glycoprotein essential for sperm fertility, is required for stable expression of Ly6k on testicular germ cells. Sci. Rep. 2016, 6, 23616. [Google Scholar] [CrossRef] [Green Version]
- Miyata, H.; Satouh, Y.; Mashiko, D.; Muto, M.; Nozawa, K.; Shiba, K.; Fujihara, Y.; Isotani, A.; Inaba, K.; Ikawa, M. Sperm calcineurin inhibition prevents mouse fertility with implications for male contraceptive. Science 2015, 350, 442–445. [Google Scholar] [CrossRef]
- Beebe, S.J.; Oyen, O.; Sandberg, M.; Froysa, A.; Hansson, V.; Jahnsen, T. Molecular cloning of a tissue-specific protein kinase (C gamma) from human testis--representing a third isoform for the catalytic subunit of cAMP-dependent protein kinase. Mol. Endocrinol. 1990, 4, 465–475. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Y.; Sun, P.B.; Li, K.; Gao, T.; Zheng, D.W.; Wu, F.P.; Liu, A.J.; Ni, Y. Protein kinases regulate hyperactivated motility of human sperm. Chin. Med. J. 2021, 134, 2412–2414. [Google Scholar] [CrossRef]
- Karlsson, M.; Zhang, C.; Mear, L.; Zhong, W.; Digre, A.; Katona, B.; Sjostedt, E.; Butler, L.; Odeberg, J.; Dusart, P.; et al. A single-cell type transcriptomics map of human tissues. Sci. Adv. 2021, 7, eabh2169. [Google Scholar] [CrossRef] [PubMed]
- Ostermeier, G.C.; Miller, D.; Huntriss, J.D.; Diamond, M.P.; Krawetz, S.A. Reproductive biology: Delivering spermatozoan RNA to the oocyte. Nature 2004, 429, 154. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Schuster, A.; Tang, C.; Yu, T.; Ortogero, N.; Bao, J.; Zheng, H.; Yan, W. Sperm-borne miRNAs and endo-siRNAs are important for fertilization and preimplantation embryonic development. Development 2016, 143, 635–647. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, M.; Jassal, B.; Stephan, R.; Milacic, M.; Rothfels, K.; Senff-Ribeiro, A.; Griss, J.; Sevilla, C.; Matthews, L.; Gong, C.; et al. The reactome pathway knowledgebase 2022. Nucleic Acids Res. 2022, 50, D687–D692. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Gene Ontology Consortium. The Gene Ontology resource: Enriching a GOld mine. Nucleic Acids Res. 2021, 49, D325–D334. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.W.; Yi, J.K.; Jeong, E.J.; Lee, W.J.; Hwang, J.M.; Kim, D.H.; Ha, J.J.; Kwon, W.S. Ras-related proteins (Rab) play significant roles in sperm motility and capacitation status. Reprod. Biol. 2022, 22, 100617. [Google Scholar] [CrossRef]
- Strom, M.; Hume, A.N.; Tarafder, A.K.; Barkagianni, E.; Seabra, M.C. A family of Rab27-binding proteins. Melanophilin links Rab27a and myosin Va function in melanosome transport. J. Biol. Chem. 2002, 277, 25423–25430. [Google Scholar] [CrossRef] [Green Version]
- Mann, U.; Shiff, B.; Patel, P. Reasons for worldwide decline in male fertility. Curr. Opin. Urol. 2020, 30, 296–301. [Google Scholar] [CrossRef]
- Hamilton, J.A.; Cissen, M.; Brandes, M.; Smeenk, J.M.; de Bruin, J.P.; Kremer, J.A.; Nelen, W.L.; Hamilton, C.J. Total motile sperm count: A better indicator for the severity of male factor infertility than the WHO sperm classification system. Hum. Reprod. 2015, 30, 1110–1121. [Google Scholar] [CrossRef] [Green Version]
- Tiegs, A.W.; Landis, J.; Garrido, N.; Scott, R.T., Jr.; Hotaling, J.M. Total Motile Sperm Count Trend Over Time: Evaluation of Semen Analyses From 119,972 Men From Subfertile Couples. Urology 2019, 132, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Salonia, A.; Bettocchi, C.; Capogrosso, P.; Carvalho, J.; Corona, G.; Hatzichristodoulou, G.; Jones, T.H.; Kadioglu, A.; Martinez-Salamanca, J.I.; Minhas, S.; et al. EAU Guidelines on Sexual and Reproductive Health. In Proceedings of the EAU Annual Congress, Milan, Italy, 10–13 March 2023. [Google Scholar]
- Sonesson, A.; Malm, J.; Rylander, L.; Giwercman, A.; Hillarp, A. Serum amyloid P component: A new biomarker for low sperm concentration? Asian J. Androl. 2021, 23, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Malm, J.; Sonesson, A.; Hellman, J.; Bjartell, A.; Frohm, B.; Hillarp, A. The pentraxin serum amyloid P component is found in the male genital tract and attached to spermatozoa. Int. J. Androl. 2008, 31, 508–517. [Google Scholar] [CrossRef]
- Maldonado-Garcia, D.; Salgado-Lucio, M.L.; Roa-Espitia, A.L.; Reyes-Miguel, T.; Hernandez-Gonzalez, E.O. Calpain inhibition prevents flotillin re-ordering and Src family activation during capacitation. Cell Tissue Res. 2017, 369, 395–412. [Google Scholar] [CrossRef] [PubMed]
- Boerke, A.; Brouwers, J.F.; Olkkonen, V.M.; van de Lest, C.H.; Sostaric, E.; Schoevers, E.J.; Helms, J.B.; Gadella, B.M. Involvement of bicarbonate-induced radical signaling in oxysterol formation and sterol depletion of capacitating mammalian sperm during in vitro fertilization. Biol. Reprod. 2013, 88, 21. [Google Scholar] [CrossRef] [Green Version]
- Angeles-Floriano, T.; Roa-Espitia, A.L.; Baltierrez-Hoyos, R.; Cordero-Martinez, J.; Elizondo, G.; Hernandez-Gonzalez, E.O. Absence of aryl hydrocarbon receptor alters CDC42 expression and prevents actin polymerization during capacitation. Mol. Reprod. Dev. 2016, 83, 1015–1026. [Google Scholar] [CrossRef]
- Kacperczyk, M.; Kmieciak, A.; Kratz, E.M. The Role of ApoE Expression and Variability of Its Glycosylation in Human Reproductive Health in the Light of Current Information. Int. J. Mol. Sci. 2021, 22, 7197. [Google Scholar] [CrossRef]
- Corbo, R.M.; Scacchi, R.; Cresta, M. Differential reproductive efficiency associated with common apolipoprotein e alleles in postreproductive-aged subjects. Fertil. Steril. 2004, 81, 104–107. [Google Scholar] [CrossRef]
- Setarehbadi, R.; Vatannejad, A.; Vaisi-Raygani, A.; Amiri, I.; Esfahani, M.; Fattahi, A.; Tavilani, H. Apolipoprotein E genotypes of fertile and infertile men. Syst. Biol. Reprod. Med. 2012, 58, 263–267. [Google Scholar] [CrossRef]
- Liu, H.; Wang, D.; Ma, Y.; Sun, H.; Wang, L.; Shi, Y.; Wang, J.; Chen, X. Hyperbaric Oxygen Therapy Ameliorates Sperm Parameters in Apolipoprotein E Knockout Mice Testes by Attenuating Oxidative Stress and Inflammation. Reprod. Sci. 2023, 1–11. [Google Scholar] [CrossRef]
- Lee, Y.; Kockx, M.; Raftery, M.J.; Jessup, W.; Griffith, R.; Kritharides, L. Glycosylation and sialylation of macrophage-derived human apolipoprotein E analyzed by SDS-PAGE and mass spectrometry: Evidence for a novel site of glycosylation on Ser290. Mol. Cell. Proteom. 2010, 9, 1968–1981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, R.; Xin, M.; Hao, Z.; You, S.; Xu, Y.; Wu, J.; Dang, L.; Zhang, X.; Sun, S. Biological Functions and Large-Scale Profiling of Protein Glycosylation in Human Semen. J. Proteome Res. 2020, 19, 3877–3889. [Google Scholar] [CrossRef] [PubMed]
- Andersen, O.M.; Yeung, C.H.; Vorum, H.; Wellner, M.; Andreassen, T.K.; Erdmann, B.; Mueller, E.C.; Herz, J.; Otto, A.; Cooper, T.G.; et al. Essential role of the apolipoprotein E receptor-2 in sperm development. J. Biol. Chem. 2003, 278, 23989–23995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wennemuth, G.; Schiemann, P.J.; Krause, W.; Gressner, A.M.; Aumuller, G. Influence of fibronectin on the motility of human spermatozoa. Int. J. Androl. 1997, 20, 10–16. [Google Scholar] [CrossRef]
- Heidari, M.; Darbandi, S.; Darbandi, M.; Akhondi, M.M.; Sadeghi, M.R. Fibronectin as a new biomarker for human sperm selection in assisted reproductive technology. Turk. J. Urol. 2019, 45, 83–90. [Google Scholar] [CrossRef]
- Martinez-Leon, E.; Osycka-Salut, C.; Signorelli, J.; Kong, M.; Morales, P.; Perez-Martinez, S.; Diaz, E.S. Fibronectin modulates the endocannabinoid system through the cAMP/PKA pathway during human sperm capacitation. Mol. Reprod. Dev. 2019, 86, 224–238. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Leon, E.; Osycka-Salut, C.; Signorelli, J.; Pozo, P.; Perez, B.; Kong, M.; Morales, P.; Perez-Martinez, S.; Diaz, E.S. Fibronectin stimulates human sperm capacitation through the cyclic AMP/protein kinase A pathway. Hum. Reprod. 2015, 30, 2138–2151. [Google Scholar] [CrossRef] [Green Version]
- Calle-Guisado, V.; Hurtado de Llera, A.; Gonzalez-Fernandez, L.; Bragado, M.J.; Garcia-Marin, L.J. Human sperm motility is downregulated by the AMPK activator A769662. Andrology 2017, 5, 1131–1140. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Perrote, A.; Hug, N.; Gonzalez-Corpas, A.; Rodriguez, C.F.; Serna, M.; Garcia-Martin, C.; Boskovic, J.; Fernandez-Leiro, R.; Caceres, J.F.; Llorca, O. Regulation of RUVBL1-RUVBL2 AAA-ATPases by the nonsense-mediated mRNA decay factor DHX34, as evidenced by Cryo-EM. Elife 2020, 9, e63042. [Google Scholar] [CrossRef]
- Puri, T.; Wendler, P.; Sigala, B.; Saibil, H.; Tsaneva, I.R. Dodecameric structure and ATPase activity of the human TIP48/TIP49 complex. J. Mol. Biol. 2007, 366, 179–192. [Google Scholar] [CrossRef]
- Doyon, Y.; Selleck, W.; Lane, W.S.; Tan, S.; Cote, J. Structural and functional conservation of the NuA4 histone acetyltransferase complex from yeast to humans. Mol. Cell. Biol. 2004, 24, 1884–1896. [Google Scholar] [CrossRef] [Green Version]
- Cindric Vranesic, A.; Reiche, J.; Hoischen, C.; Wohlmann, A.; Bratsch, J.; Friedrich, K.; Gunes, B.; Cappallo-Obermann, H.; Kirchhoff, C.; Diekmann, S.; et al. Characterization of SKAP/kinastrin isoforms: The N-terminus defines tissue specificity and Pontin binding. Hum. Mol. Genet. 2016, 25, 2838–2852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulon, S.; Bertrand, E.; Pradet-Balade, B. HSP90 and the R2TP co-chaperone complex: Building multi-protein machineries essential for cell growth and gene expression. RNA Biol. 2012, 9, 148–154. [Google Scholar] [CrossRef] [Green Version]
- Sharpe, R.M. Environment, lifestyle and male infertility. Baillieres Best Pract. Res. Clin. Endocrinol. Metab. 2000, 14, 489–503. [Google Scholar] [CrossRef]
- Male Infertility Best Practice Policy Committee of the American Urological Association; Practice Committee of the American Society for Reproductive Medicine. Report on management of obstructive azoospermia. Fertil. Steril. 2004, 82 (Suppl. S1), S137–S141. [Google Scholar] [CrossRef] [PubMed]
- Adomaitis, R.; Vincel, B.; Eidukaite, A.; Ostaneviciute, E.; Kirka, R.; Bilius, V.; Malcius, D.; Verkauskas, G.; Hadziselimovic, F. Consequences of bilateral cryptorchidism in adults. Andrologia 2016, 48, 933–938. [Google Scholar] [CrossRef] [PubMed]
- Wosnitzer, M.S.; Goldstein, M. Obstructive azoospermia. Urol. Clin. N. Am. 2014, 41, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.J.; Kim, B.S.; Kim, H.J.; Cho, Y.D.; Shin, H.L.; Yoon, H.I.; Lee, Y.S.; Baek, J.H.; Woo, K.M.; Ryoo, H.M. Intratesticular Peptidyl Prolyl Isomerase 1 Protein Delivery Using Cationic Lipid-Coated Fibroin Nanoparticle Complexes Rescues Male Infertility in Mice. ACS Nano 2020, 14, 13217–13231. [Google Scholar] [CrossRef]
- Leader, B.; Baca, Q.J.; Golan, D.E. Protein therapeutics: A summary and pharmacological classification. Nat. Rev. Drug. Discov. 2008, 7, 21–39. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | N (n = 31) | AN (n = 22) | OA (n = 9) | A (n = 13) | N vs. AN | N vs. OA | N vs. A |
|---|---|---|---|---|---|---|---|
| Age | 35.8 ± 7.3 | 33.5 ± 8.3 | 33.2 ± 7.2 | 33.7 ± 9.3 | n.s. | n.s. | n.s. |
| Volume (mL) | 3.3 ± 1.8 | 4.1 ± 2.8 | 4.5 ± 3.4 | 3.8 ± 2.4 | n.s. | n.s. | n.s. |
| Count (106/mL) | 86.7 ± 39.6 | 26.9 ± 19.6 | 9.1 ± 3.4 | 39.2 ± 16.3 | *** | *** | *** |
| Motility (% motile) | 52.0 ± 10.6 | 12.3 ± 9.0 | 14.9 ± 11.3 | 10.5 ± 7.0 | *** | *** | *** |
| Morphology (%) | 7.0 ± 8.2 | 9.6 ± 5.7 | 6.2 ± 3.8 | 11.9 ± 5.8 | n.s. | n.s. | * |
| A: Negatively Correlated Proteins | |||||||
| Experimental ID | Gene Symbol | Motility (% motile) | Count (106/mL) | ||||
| r | p-Value | Correlation | r | p-Value | Correlation | ||
| 3707 | NUCB2 | −0.78 | 8.65 × 10−12 | negative | −0.52 | 6.90 × 10−5 | negative |
| 2530 | LPL | −0.74 | 2.86 × 10−10 | negative | −0.6 | 1.97 × 10−6 | negative |
| 3510 | LAMB2 | −0.71 | 3.75 × 10−9 | negative | −0.5 | 1.70 × 10−4 | - |
| 2938 | PPIB | −0.71 | 3.53 × 10−9 | negative | −0.5 | 1.37 × 10−4 | negative |
| 3774 | SEMG2 | −0.71 | 3.38 × 10−9 | negative | −0.52 | 6.75 × 10−5 | negative |
| 4530 | CPZ | −0.69 | 1.33 × 10−8 | negative | −0.58 | 6.18 × 10−6 | negative |
| 4609 | VWA1 | −0.68 | 1.24 × 10−7 | negative | −0.42 | 3.04 × 10−3 | - |
| 2475 | SEMG1 | −0.67 | 4.83 × 10−8 | negative | −0.49 | 1.84 × 10−4 | - |
| 2497 | SERPINA5 | −0.66 | 5.81 × 10−8 | negative | −0.5 | 1.38 × 10−4 | - |
| 2521 | APOA4 | −0.65 | 7.96 × 10−6 | negative | −0.39 | 1.50 × 10−2 | - |
| 1961 | LAMA5 | −0.65 | 1.45 × 10−7 | negative | −0.52 | 7.64 × 10−5 | negative |
| 3782 | PLOD1 | −0.65 | 8.84 × 10−7 | negative | −0.61 | 5.77 × 10−6 | negative |
| 2673 | CLU | −0.64 | 1.89 × 10−7 | negative | −0.4 | 3.29 × 10−3 | - |
| 3308 | HSPA13 | −0.64 | 5.39 × 10−6 | negative | −0.49 | 1.00 × 10−3 | - |
| 5763 | NEU1 | −0.64 | 1.07 × 10−5 | negative | −0.5 | 1.04 × 10−3 | - |
| 2431 | FN1 | −0.63 | 3.64 × 10−7 | negative | −0.46 | 4.95 × 10−4 | - |
| 2420 | APOE | −0.62 | 1.47 × 10−5 | negative | −0.33 | 3.44 × 10−2 | - |
| 2675 | LAMC1 | −0.62 | 1.30 × 10−6 | negative | −0.42 | 1.99 × 10−3 | - |
| 2722 | CDH1 | −0.6 | 1.71 × 10−6 | negative | −0.52 | 6.87 × 10−5 | negative |
| 1832 | MATN2 | −0.6 | 2.98 × 10−5 | negative | −0.33 | 3.12 × 10−2 | - |
| 2516 | GLA | −0.59 | 4.95 × 10−5 | negative | −0.37 | 1.60 × 10−2 | - |
| 4767 | MAMDC2 | −0.59 | 9.70 × 10−5 | negative | −0.45 | 4.17 × 10−3 | - |
| 6191 | SCPEP1 | −0.59 | 7.62 × 10−6 | negative | −0.52 | 9.98 × 10−5 | negative |
| 131 | CTSH | −0.58 | 1.19 × 10−5 | negative | −0.47 | 4.93 × 10−4 | - |
| 1642 | CAMP | −0.56 | 1.42 × 10−5 | negative | −0.28 | 4.04 × 10−2 | - |
| 2531 | HEXA | −0.56 | 2.26 × 10−5 | negative | −0.47 | 5.30 × 10−4 | - |
| 2419 | APOA1 | −0.55 | 2.68 × 10−4 | negative | −0.32 | 4.91 × 10−2 | - |
| 2096 | PLOD3 | −0.55 | 2.57 × 10−4 | negative | −0.42 | 7.00 × 10−3 | - |
| 5836 | SDF4 | −0.55 | 1.70 × 10−4 | negative | −0.26 | 1.06 × 10−1 | - |
| 3918 | DNAJC3 | −0.53 | 9.86 × 10−5 | negative | −0.41 | 3.37 × 10−3 | - |
| 6045 | SIL1 | −0.53 | 8.01 × 10−5 | negative | −0.46 | 8.97 × 10−4 | - |
| 3180 | COL18A1 | −0.51 | 4.09 × 10−4 | negative | −0.32 | 3.42 × 10−2 | - |
| 2572 | MMP2 | −0.51 | 3.76 × 10−4 | negative | −0.26 | 8.85 × 10−2 | - |
| 3228 | PRCP | −0.5 | 8.23 × 10−4 | negative | −0.43 | 5.04 × 10−3 | - |
| 4643 | SPESP1 | −0.5 | 4.02 × 10−4 | negative | −0.11 | 4.58 × 10−1 | - |
| 2536 | P4HB | −0.48 | 2.41 × 10−4 | - | −0.56 | 1.05 × 10−5 | negative |
| 5898 | VAMP8 | −0.4 | 1.04 × 10−2 | - | −0.51 | 7.20 × 10−4 | negative |
| 3601 | HNRNPK | −0.38 | 7.34 × 10−3 | - | −0.57 | 1.81 × 10−5 | negative |
| 2304 | AHSA1 | −0.35 | 2.47 × 10−2 | - | −0.59 | 5.60 × 10−5 | negative |
| B: positively correlated proteins | |||||||
| Experimental ID | Gene symbol | Progressive sperm motility | Sperm count | ||||
| r | p-value | Correlation | r | p-value | Correlation | ||
| 4245 | TKFC | 0.68 | 2.41 × 10−8 | positive | 0.49 | 1.83 × 10−4 | - |
| 2526 | GPI | 0.68 | 1.97 × 10−8 | positive | 0.5 | 1.18 × 10−4 | positive |
| 3026 | IMPA1 | 0.68 | 6.53 × 10−8 | positive | 0.64 | 5.63 × 10−7 | positive |
| 4778 | HDDC2 | 0.64 | 5.55 × 10−6 | positive | 0.56 | 1.27 × 10−4 | positive |
| 3299 | PPP3CC | 0.63 | 1.96 × 10−5 | positive | 0.67 | 3.80 × 10−6 | positive |
| 4544 | CYB5R2 | 0.61 | 3.49 × 10−6 | positive | 0.68 | 6.85 × 10−8 | positive |
| 2227 | FLOT1 | 0.6 | 7.35 × 10−6 | positive | 0.3 | 3.84 × 10−2 | - |
| 2823 | GOT1 | 0.56 | 1.80 × 10−5 | positive | 0.51 | 1.55 × 10−4 | positive |
| 2923 | PRKACG | 0.56 | 1.34 × 10−4 | positive | 0.68 | 5.98 × 10−7 | positive |
| 632 | ARRDC5 | 0.55 | 7.40 × 10−5 | positive | 0.67 | 2.93 × 10−7 | positive |
| 6391 | DPP3 | 0.54 | 4.40 × 10−5 | positive | 0.28 | 5.02 × 10−2 | - |
| 2997 | STOM | 0.54 | 4.85 × 10−5 | positive | 0.46 | 7.69 × 10−4 | - |
| 4198 | UGP2 | 0.54 | 3.44 × 10−5 | positive | 0.37 | 5.79 × 10−3 | - |
| 6619 | TSPAN16 | 0.53 | 2.09 × 10−4 | positive | 0.73 | 1.64 × 10−8 | positive |
| 3116 | HSPA4 | 0.52 | 6.24 × 10−5 | positive | 0.4 | 2.70 × 10−3 | - |
| 4301 | PYCRL | 0.52 | 2.58 × 10−4 | positive | 0.29 | 5.51 × 10−2 | - |
| 1079 | TCEB1 | 0.52 | 7.86 × 10−5 | positive | 0.56 | 1.68 × 10−5 | positive |
| 4332 | ACSBG2 | 0.51 | 1.31 × 10−4 | positive | 0.59 | 8.01 × 10−6 | positive |
| 4838 | CAND1 | 0.51 | 9.96 × 10−5 | positive | 0.4 | 3.24 × 10−3 | - |
| 2426 | APCS | 0.5 | 1.27 × 10−4 | positive | 0.62 | 7.41 × 10−7 | positive |
| 3976 | FHL1 | 0.49 | 7.70 × 10−4 | - | 0.61 | 1.34 × 10−5 | positive |
| 5742 | MYADM | 0.49 | 4.10 × 10−4 | - | 0.55 | 5.50 × 10−5 | positive |
| 3025 | TPP2 | 0.48 | 3.03 × 10−4 | - | 0.57 | 1.13 × 10−5 | positive |
| 3972 | CUL3 | 0.47 | 3.59 × 10−4 | - | 0.54 | 2.72 × 10−5 | positive |
| 4179 | SYPL1 | 0.47 | 5.68 × 10−4 | - | 0.54 | 4.37 × 10−5 | positive |
| 4655 | GLB1L | 0.46 | 1.67 × 10−3 | - | 0.61 | 1.06 × 10−5 | positive |
| 6705 | RUVBL1 | 0.45 | 8.14 × 10−4 | - | 0.68 | 2.41 × 10−8 | positive |
| 5910 | DPCD | 0.43 | 1.28 × 10−3 | - | 0.54 | 2.94 × 10−5 | positive |
| 6313 | FARSB | 0.43 | 1.94 × 10−3 | - | 0.54 | 5.55 × 10−5 | positive |
| 2616 | UCHL1 | 0.43 | 5.71 × 10−3 | - | 0.51 | 8.25 × 10−4 | positive |
| 1810 | ACOT7 | 0.41 | 9.95 × 10−3 | - | 0.51 | 9.30 × 10−4 | positive |
| 6860 | CFAP20 | 0.4 | 1.00 × 10−2 | - | 0.51 | 8.26 × 10−4 | positive |
| 5950 | TEX101 | 0.4 | 7.97 × 10−3 | - | 0.53 | 2.43 × 10−4 | positive |
| 1189 | TSN | 0.39 | 6.80 × 10−3 | - | 0.56 | 3.21 × 10−5 | positive |
| 6190 | C12orf10 | 0.37 | 1.04 × 10−2 | - | 0.5 | 3.79 × 10−4 | positive |
| 2535 | PGK2 | 0.37 | 5.69 × 10−3 | - | 0.6 | 1.75 × 10−6 | positive |
| 6698 | RUVBL2 | 0.36 | 7.23 × 10−3 | - | 0.53 | 3.91 × 10−5 | positive |
| 2721 | ACE | 0.34 | 1.32 × 10−2 | - | 0.59 | 3.06 × 10−6 | positive |
| 436 | EFR3A | 0.34 | 2.81 × 10−2 | - | 0.53 | 3.71 × 10−4 | positive |
| 6708 | VDAC3 | 0.34 | 1.20 × 10−2 | - | 0.52 | 7.06 × 10−5 | positive |
| 5290 | SLC2A14 | 0.33 | 1.81 × 10−2 | - | 0.5 | 1.49 × 10−4 | positive |
| 2690 | ACADM | 0.31 | 2.80 × 10−2 | - | 0.51 | 1.79 × 10−4 | positive |
| 3883 | TRAP1 | 0.29 | 6.17 × 10−2 | - | 0.52 | 4.47 × 10−4 | positive |
| 3502 | AQP5 | 0.24 | 9.10 × 10−2 | - | 0.58 | 8.00 × 10−6 | positive |
| 6150 | ACAD9 | 0.23 | 1.49 × 10−1 | - | 0.5 | 7.45 × 10−4 | positive |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Becker, L.S.; Al Smadi, M.A.; Raeschle, M.; Rishik, S.; Abdul-Khaliq, H.; Meese, E.; Abu-Halima, M. Proteomic Landscape of Human Sperm in Patients with Different Spermatogenic Impairments. Cells 2023, 12, 1017. https://doi.org/10.3390/cells12071017
Becker LS, Al Smadi MA, Raeschle M, Rishik S, Abdul-Khaliq H, Meese E, Abu-Halima M. Proteomic Landscape of Human Sperm in Patients with Different Spermatogenic Impairments. Cells. 2023; 12(7):1017. https://doi.org/10.3390/cells12071017
Chicago/Turabian StyleBecker, Lea Simone, Mohammad A. Al Smadi, Markus Raeschle, Shusruto Rishik, Hashim Abdul-Khaliq, Eckart Meese, and Masood Abu-Halima. 2023. "Proteomic Landscape of Human Sperm in Patients with Different Spermatogenic Impairments" Cells 12, no. 7: 1017. https://doi.org/10.3390/cells12071017