
Current Advancements in Spinal Cord Injury Research—Glial Scar Formation and Neural Regeneration

Abstract
:1. Introduction
1.1. Cellular Events Immediately following Trauma
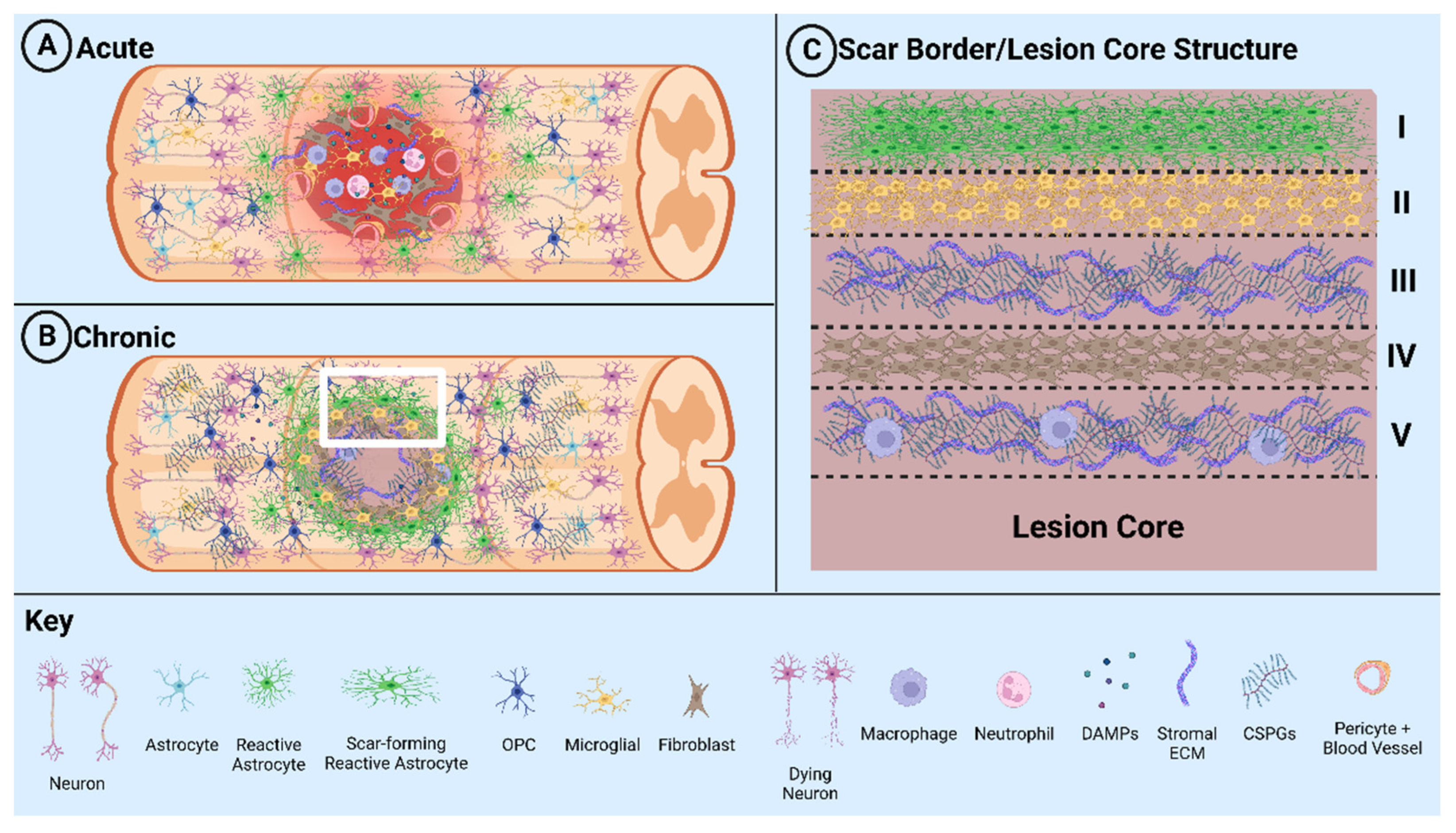
1.2. Glial Scar Formation
1.3. Positive and Negative Effects of the Glial Scar
2. Current Research in Therapeutic Interventions
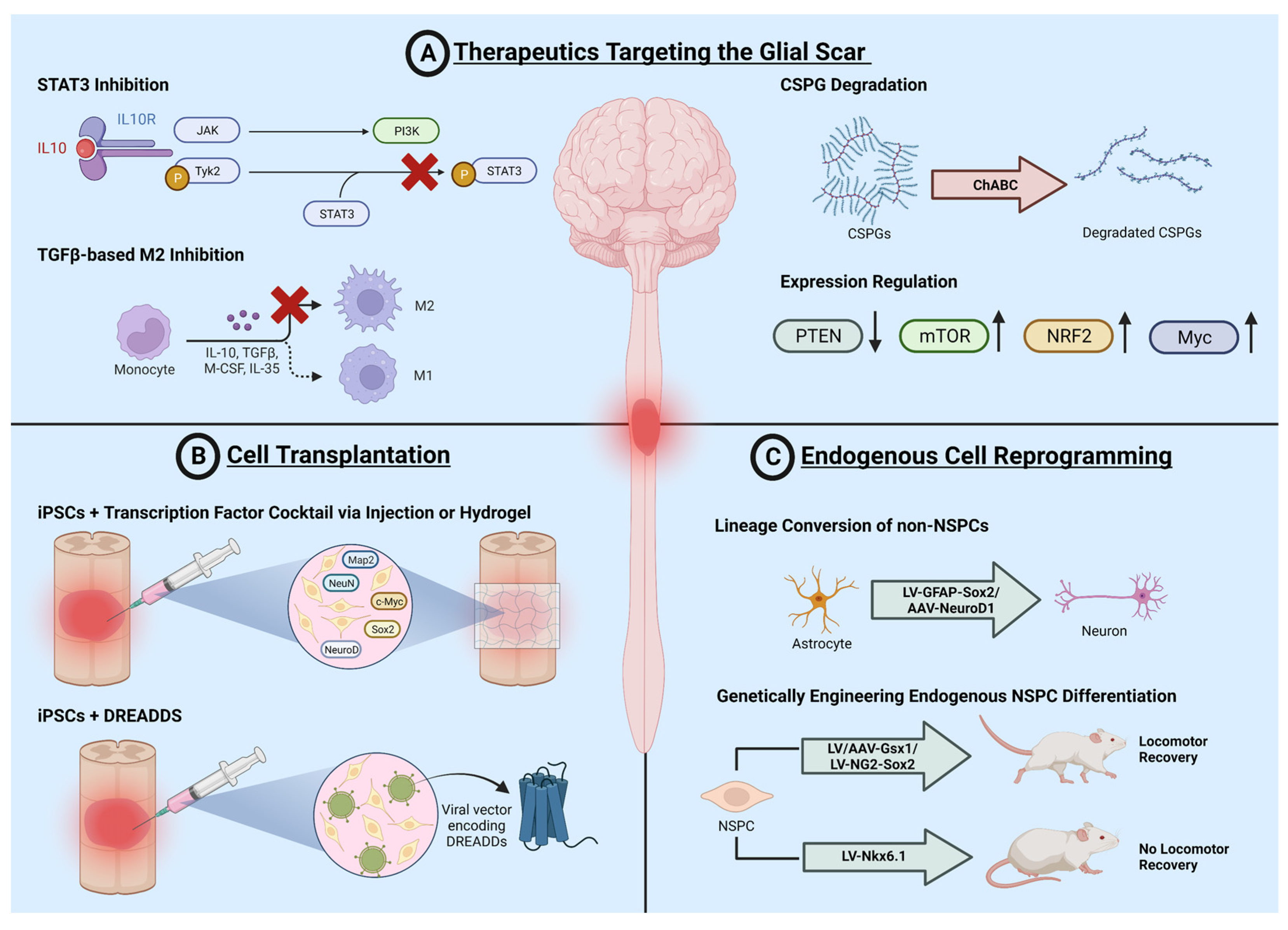
2.1. Therapeutics Targeting the Glial Scar
2.1.1. Targeting Scar Formation
2.1.2. Resolving the Glial Scar
2.2. Cell Transplantation
2.3. Endogenous Cell Reprogramming
3. Conclusions and Future Directions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ding, W.; Hu, S.; Wang, P.; Kang, H.; Peng, R.; Dong, Y.; Li, F. Spinal cord injury: The global incidence, prevalence, and disability from the global burden of disease study 2019. Spine 2022, 47, 1532–1540. [Google Scholar] [CrossRef] [PubMed]
- McRae, J.; Morgan, S.; Wallace, E.; Miles, A. Oropharyngeal dysphagia in acute cervical spinal cord injury: A literature review. Dysphagia 2022. [Google Scholar] [CrossRef] [PubMed]
- Tran, A.P.; Warren, P.M.; Silver, J. New insights into glial scar formation after spinal cord injury. Cell Tissue Res. 2022, 387, 319–336. [Google Scholar] [CrossRef] [PubMed]
- Schwab, M.E.; Strittmatter, S.M. Nogo limits neural plasticity and recovery from injury. Curr. Opin. Neurobiol. 2014, 27, 53–60. [Google Scholar] [CrossRef] [Green Version]
- McKeon, R.J.; Schreiber, R.C.; Rudge, J.S.; Silver, J. Reduction of neurite outgrowth in a model of glial scarring following CNS injury is correlated with the expression of inhibitory molecules on reactive astrocytes. J. Neurosci. 1991, 11, 3398–3411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKerracher, L.; Rosen, K.M. MAG, myelin and overcoming growth inhibition in the CNS. Front. Mol. Neurosci. 2015, 8, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Wang, D.; Jiang, Y.; Ma, H.; Li, X.; Wang, H. Dexmedetomidine postconditioning alleviates spinal cord ischemia-reperfusion injury in rats via inhibiting neutrophil infiltration, microglia activation, reactive gliosis and CXCL13/CXCR5 axis activation. Int. J. Neurosci. 2023, 133, 1–12. [Google Scholar] [CrossRef]
- Hashimoto, S.; Nagoshi, N.; Shinozaki, M.; Nakanishi, K.; Suematsu, Y.; Shibata, T.; Kawai, M.; Kitagawa, T.; Ago, K.; Kamata, Y.; et al. Microenvironmental modulation in tandem with human stem cell transplantation enhances functional recovery after chronic complete spinal cord injury. Biomaterials 2023, 295, 122002. [Google Scholar] [CrossRef]
- Fan, B.; Wei, Z.; Yao, X.; Shi, G.; Cheng, X.; Zhou, X.; Zhou, H.; Ning, G.; Kong, X.; Feng, S. Microenvironment imbalance of spinal cord injury. Cell Transplant. 2018, 27, 853–866. [Google Scholar] [CrossRef] [Green Version]
- Daniel, P.M.; Strich, S.J. Histological observations on Wallerian degeneration in the spinal cord of the baboon, Papio papio. Acta Neuropathol. 1969, 12, 314–328. [Google Scholar] [CrossRef]
- Fischer, T.; Stern, C.; Freund, P.; Schubert, M.; Sutter, R. Wallerian degeneration in cervical spinal cord tracts is commonly seen in routine T2-weighted MRI after traumatic spinal cord injury and is associated with impairment in a retrospective study. Eur. Radiol. 2021, 31, 2923–2932. [Google Scholar] [CrossRef] [PubMed]
- Totoiu, M.O.; Keirstead, H.S. Spinal cord injury is accompanied by chronic progressive demyelination. J. Comp. Neurol. 2005, 486, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Benson, M.D.; Romero, M.I.; Lush, M.E.; Lu, Q.R.; Henkemeyer, M.; Parada, L.F. Ephrin-B3 is a myelin-based inhibitor of neurite outgrowth. Proc. Natl. Acad. Sci. USA 2005, 102, 10694–10699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahuja, C.S.; Nori, S.; Tetreault, L.; Wilson, J.; Kwon, B.; Harrop, J.; Choi, D.; Fehlings, M.G. Traumatic Spinal Cord Injury-Repair and Regeneration. Neurosurgery 2017, 80, S9–S22. [Google Scholar] [CrossRef]
- Anjum, A.; Yazid, M.D.; Fauzi Daud, M.; Idris, J.; Ng, A.M.H.; Selvi Naicker, A.; Ismail, O.H.R.; Athi Kumar, R.K.; Lokanathan, Y. Spinal cord injury: Pathophysiology, multimolecular interactions, and underlying recovery mechanisms. Int. J. Mol. Sci. 2020, 21, 7533. [Google Scholar] [CrossRef]
- Bradbury, E.J.; Burnside, E.R. Moving beyond the glial scar for spinal cord repair. Nat. Commun. 2019, 10, 3879. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.J.; Vadakkan, K.I.; Zhuo, M. ATP-induced chemotaxis of microglial processes requires P2Y receptor-activated initiation of outward potassium currents. Glia 2007, 55, 810–821. [Google Scholar] [CrossRef]
- Davalos, D.; Grutzendler, J.; Yang, G.; Kim, J.V.; Zuo, Y.; Jung, S.; Littman, D.R.; Dustin, M.L.; Gan, W.B. ATP mediates rapid microglial response to local brain injury in vivo. Nat. Neurosci. 2005, 8, 752–758. [Google Scholar] [CrossRef]
- Fu, J.; Mu, G.; Liu, X.; Ou, C.; Zhao, J. Ischemic postconditioning reduces spinal cord ischemia-reperfusion injury through ATP-sensitive potassium channel. Spinal Cord 2022, 60, 326–331. [Google Scholar] [CrossRef]
- Franke, H.; Krugel, U.; Schmidt, R.; Grosche, J.; Reichenbach, A.; Illes, P. P2 receptor-types involved in astrogliosis in vivo. Br. J. Pharmacol. 2001, 134, 1180–1189. [Google Scholar] [CrossRef] [Green Version]
- Shinozaki, Y.; Shibata, K.; Yoshida, K.; Shigetomi, E.; Gachet, C.; Ikenaka, K.; Tanaka, K.F.; Koizumi, S. Transformation of Astrocytes to a Neuroprotective Phenotype by Microglia via P2Y(1) Receptor Downregulation. Cell Rep. 2017, 19, 1151–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabelstrom, H.; Stenudd, M.; Reu, P.; Dias, D.O.; Elfineh, M.; Zdunek, S.; Damberg, P.; Goritz, C.; Frisen, J. Resident neural stem cells restrict tissue damage and neuronal loss after spinal cord injury in mice. Science 2013, 342, 637–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, C.; Arnold, P.M.; Zoubine, M.N.; Citron, B.A.; Watanabe, I.; Berman, N.E.; Festoff, B.W. Apoptosis in cellular compartments of rat spinal cord after severe contusion injury. J. Neurotrauma 1998, 15, 459–472. [Google Scholar] [CrossRef]
- Cheng, R.; Zhu, G.; Ni, C.; Wang, R.; Sun, P.; Tian, L.; Zhang, L.; Zhang, J.; Ye, X.; Luo, B. P2Y2 receptor mediated neuronal regeneration and angiogenesis to affect functional recovery in rats with spinal cord injury. Neural. Plast. 2022, 2022, 2191011. [Google Scholar] [CrossRef] [PubMed]
- Wanner, I.B.; Anderson, M.A.; Song, B.; Levine, J.; Fernandez, A.; Gray-Thompson, Z.; Ao, Y.; Sofroniew, M.V. Glial scar borders are formed by newly proliferated, elongated astrocytes that interact to corral inflammatory and fibrotic cells via STAT3-dependent mechanisms after spinal cord injury. J. Neurosci. 2013, 33, 12870–12886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartus, K.; Burnside, E.R.; Galino, J.; James, N.D.; Bennett, D.L.H.; Bradbury, E.J. ErbB receptor signaling directly controls oligodendrocyte progenitor cell transformation and spontaneous remyelination after spinal cord injury. Glia 2019, 67, 1036–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assinck, P.; Duncan, G.J.; Plemel, J.R.; Lee, M.J.; Stratton, J.A.; Manesh, S.B.; Liu, J.; Ramer, L.M.; Kang, S.H.; Bergles, D.E.; et al. Myelinogenic plasticity of oligodendrocyte precursor cells following spinal cord contusion injury. J. Neurosci. 2017, 37, 8635–8654. [Google Scholar] [CrossRef]
- Levine, J. The reactions and role of NG2 glia in spinal cord injury. Brain Res. 2016, 1638, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Dias, D.O.; Goritz, C. Fibrotic scarring following lesions to the central nervous system. Matrix Biol. 2018, 68–69, 561–570. [Google Scholar] [CrossRef]
- Yu, P.; Wang, H.; Katagiri, Y.; Geller, H.M. An in vitro model of reactive astrogliosis and its effect on neuronal growth. Methods Mol. Biol. 2012, 814, 327–340. [Google Scholar] [CrossRef] [Green Version]
- Noristani, H.N.; Sabourin, J.C.; Boukhaddaoui, H.; Chan-Seng, E.; Gerber, Y.N.; Perrin, F.E. Spinal cord injury induces astroglial conversion towards neuronal lineage. Mol. Neurodegener. 2016, 11, 68. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Bi, H.; Ge, Q.; Teng, H.; Wan, G.; Yu, B.; Jiang, Q.; Gu, X. Heterogeneity analysis of astrocytes following spinal cord injury at single-cell resolution. FASEB J. 2022, 36, e22442. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Geoffroy, C.G.; Meves, J.M.; Narang, A.; Li, Y.; Nguyen, M.T.; Khai, V.S.; Kong, X.; Steinke, C.L.; Carolino, K.I.; et al. Leucine zipper-bearing kinase is a critical regulator of astrocyte reactivity in the adult mammalian CNS. Cell Rep. 2018, 22, 3587–3597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrmann, J.E.; Imura, T.; Song, B.; Qi, J.; Ao, Y.; Nguyen, T.K.; Korsak, R.A.; Takeda, K.; Akira, S.; Sofroniew, M.V. STAT3 is a critical regulator of astrogliosis and scar formation after spinal cord injury. J. Neurosci. 2008, 28, 7231–7243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, C.; Shen, X.; Xu, X.; Liu, H.; Li, F.; Lu, S.; Gao, Z.; Zhang, J.; Wu, Q.; Yang, D.; et al. Astrocytic YAP promotes the formation of glia scars and neural regeneration after spinal cord injury. J. Neurosci. 2020, 40, 2644–2662. [Google Scholar] [CrossRef]
- Anderson, M.A.; Burda, J.E.; Ren, Y.; Ao, Y.; O’Shea, T.M.; Kawaguchi, R.; Coppola, G.; Khakh, B.S.; Deming, T.J.; Sofroniew, M.V. Astrocyte scar formation aids central nervous system axon regeneration. Nature 2016, 532, 195–200. [Google Scholar] [CrossRef] [Green Version]
- Filous, A.R.; Tran, A.; Howell, C.J.; Busch, S.A.; Evans, T.A.; Stallcup, W.B.; Kang, S.H.; Bergles, D.E.; Lee, S.I.; Levine, J.M.; et al. Entrapment via synaptic-like connections between NG2 proteoglycan+ cells and dystrophic axons in the lesion plays a role in regeneration failure after spinal cord injury. J. Neurosci. 2014, 34, 16369–16384. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Soderblom, C.; Trojanowsky, M.; Lee, D.H.; Lee, J.K. Fibronectin matrix assembly after spinal cord injury. J. Neurotrauma 2015, 32, 1158–1167. [Google Scholar] [CrossRef] [Green Version]
- McKillop, W.M.; Dragan, M.; Schedl, A.; Brown, A. Conditional Sox9 ablation reduces chondroitin sulfate proteoglycan levels and improves motor function following spinal cord injury. Glia 2013, 61, 164–177. [Google Scholar] [CrossRef] [Green Version]
- Lein, P.J.; Higgins, D.; Turner, D.C.; Flier, L.A.; Terranova, V.P. The NC1 domain of type IV collagen promotes axonal growth in sympathetic neurons through interaction with the alpha 1 beta 1 integrin. J. Cell Biol. 1991, 113, 417–428. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Goldberg, D.J. Bundling of microtubules in the growth cone induced by laminin. Mol. Cell Neurosci. 2000, 15, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Kobayakawa, K.; Ohkawa, Y.; Kumamaru, H.; Yokota, K.; Saito, T.; Kijima, K.; Yoshizaki, S.; Harimaya, K.; Nakashima, Y.; et al. Interaction of reactive astrocytes with type I collagen induces astrocytic scar formation through the integrin-N-cadherin pathway after spinal cord injury. Nat. Med. 2017, 23, 818–828. [Google Scholar] [CrossRef] [PubMed]
- Yokota, K.; Kobayakawa, K.; Saito, T.; Hara, M.; Kijima, K.; Ohkawa, Y.; Harada, A.; Okazaki, K.; Ishihara, K.; Yoshida, S.; et al. Periostin promotes scar formation through the interaction between pericytes and infiltrating monocytes/macrophages after spinal cord injury. Am. J. Pathol. 2017, 187, 639–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goritz, C.; Dias, D.O.; Tomilin, N.; Barbacid, M.; Shupliakov, O.; Frisen, J. A pericyte origin of spinal cord scar tissue. Science 2011, 333, 238–242. [Google Scholar] [CrossRef]
- Soderblom, C.; Luo, X.; Blumenthal, E.; Bray, E.; Lyapichev, K.; Ramos, J.; Krishnan, V.; Lai-Hsu, C.; Park, K.K.; Tsoulfas, P.; et al. Perivascular fibroblasts form the fibrotic scar after contusive spinal cord injury. J. Neurosci. 2013, 33, 13882–13887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reier, P.J.; Houle, J.D. The glial scar: Its bearing on axonal elongation and transplantation approaches to CNS repair. Adv. Neurol. 1988, 47, 87–138. [Google Scholar]
- Bellver-Landete, V.; Bretheau, F.; Mailhot, B.; Vallieres, N.; Lessard, M.; Janelle, M.E.; Vernoux, N.; Tremblay, M.E.; Fuehrmann, T.; Shoichet, M.S.; et al. Microglia are an essential component of the neuroprotective scar that forms after spinal cord injury. Nat. Commun. 2019, 10, 518. [Google Scholar] [CrossRef] [Green Version]
- David, S.; Kroner, A. Repertoire of microglial and macrophage responses after spinal cord injury. Nat. Rev. Neurosci. 2011, 12, 388–399. [Google Scholar] [CrossRef]
- Zhou, X.; Wahane, S.; Friedl, M.S.; Kluge, M.; Friedel, C.C.; Avrampou, K.; Zachariou, V.; Guo, L.; Zhang, B.; He, X.; et al. Microglia and macrophages promote corralling, wound compaction and recovery after spinal cord injury via Plexin-B2. Nat. Neurosci. 2020, 23, 337–350. [Google Scholar] [CrossRef]
- Yoshizaki, S.; Tamaru, T.; Hara, M.; Kijima, K.; Tanaka, M.; Konno, D.J.; Matsumoto, Y.; Nakashima, Y.; Okada, S. Microglial inflammation after chronic spinal cord injury is enhanced by reactive astrocytes via the fibronectin/beta1 integrin pathway. J. Neuroinflamm. 2021, 18, 12. [Google Scholar] [CrossRef]
- Kuboyama, T.; Kominato, S.; Nagumo, M.; Tohda, C. Recovery from spinal cord injury via M2 microglial polarization induced by Polygalae Radix. Phytomedicine 2021, 82, 153452. [Google Scholar] [CrossRef] [PubMed]
- Greenhalgh, A.D.; Zarruk, J.G.; Healy, L.M.; Baskar Jesudasan, S.J.; Jhelum, P.; Salmon, C.K.; Formanek, A.; Russo, M.V.; Antel, J.P.; McGavern, D.B.; et al. Peripherally derived macrophages modulate microglial function to reduce inflammation after CNS injury. PLoS Biol. 2018, 16, e2005264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mildner, A.; Mack, M.; Schmidt, H.; Bruck, W.; Djukic, M.; Zabel, M.D.; Hille, A.; Priller, J.; Prinz, M. CCR2 + Ly-6Chi monocytes are crucial for the effector phase of autoimmunity in the central nervous system. Brain 2009, 132, 2487–2500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shechter, R.; Miller, O.; Yovel, G.; Rosenzweig, N.; London, A.; Ruckh, J.; Kim, K.W.; Klein, E.; Kalchenko, V.; Bendel, P.; et al. Recruitment of beneficial M2 macrophages to injured spinal cord is orchestrated by remote brain choroid plexus. Immunity 2013, 38, 555–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Guo, W.; Zhu, Z.; Hu, Y.; Wang, Y.; Zhang, X.; Wang, W.; Du, N.; Song, T.; Yang, K.; et al. Macrophage migration inhibitory factor facilitates production of CCL5 in astrocytes following rat spinal cord injury. J. Neuroinflamm. 2018, 15, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miron, V.E.; Boyd, A.; Zhao, J.W.; Yuen, T.J.; Ruckh, J.M.; Shadrach, J.L.; van Wijngaarden, P.; Wagers, A.J.; Williams, A.; Franklin, R.J.M.; et al. M2 microglia and macrophages drive oligodendrocyte differentiation during CNS remyelination. Nat. Neurosci. 2013, 16, 1211–1218. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Cao, K.; Sun, X.; Chen, Y.; Duan, Z.; Sun, L.; Guo, L.; Bai, P.; Sun, D.; Fan, J.; et al. Macrophages in spinal cord injury: Phenotypic and functional change from exposure to myelin debris. Glia 2015, 63, 635–651. [Google Scholar] [CrossRef]
- Zhu, Y.; Soderblom, C.; Krishnan, V.; Ashbaugh, J.; Bethea, J.R.; Lee, J.K. Hematogenous macrophage depletion reduces the fibrotic scar and increases axonal growth after spinal cord injury. Neurobiol. Dis. 2015, 74, 114–125. [Google Scholar] [CrossRef] [Green Version]
- Finkel, Z.; Esteban, F.; Rodriguez, B.; Fu, T.; Ai, X.; Cai, L. Diversity of adult neural stem and progenitor cells in physiology and disease. Cells 2021, 10, 2045. [Google Scholar] [CrossRef]
- Mothe, A.J.; Tator, C.H. Proliferation, migration, and differentiation of endogenous ependymal region stem/progenitor cells following minimal spinal cord injury in the adult rat. Neuroscience 2005, 131, 177–187. [Google Scholar] [CrossRef]
- Meletis, K.; Barnabe-Heider, F.; Carlen, M.; Evergren, E.; Tomilin, N.; Shupliakov, O.; Frisen, J. Spinal cord injury reveals multilineage differentiation of ependymal cells. PLoS Biol. 2008, 6, e182. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, A.; Komitova, M.; Suzuki, R.; Zhu, X. Polydendrocytes (NG2 cells): Multifunctional cells with lineage plasticity. Nat. Rev. Neurosci. 2009, 10, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Hughes, E.G.; Kang, S.H.; Fukaya, M.; Bergles, D.E. Oligodendrocyte progenitors balance growth with self-repulsion to achieve homeostasis in the adult brain. Nat. Neurosci. 2013, 16, 668–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hesp, Z.C.; Yoseph, R.Y.; Suzuki, R.; Jukkola, P.; Wilson, C.; Nishiyama, A.; McTigue, D.M. Proliferating NG2-cell-dependent angiogenesis and scar formation alter axon growth and functional recovery after spinal cord injury in mice. J. Neurosci. 2018, 38, 1366–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukovic, D.; Stojkovic, M.; Moreno-Manzano, V.; Jendelova, P.; Sykova, E.; Bhattacharya, S.S.; Erceg, S. Concise review: Reactive astrocytes and stem cells in spinal cord injury: Good guys or bad guys? Stem Cells 2015, 33, 1036–1041. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Barres, B.A. Regeneration: Not everything is scary about a glial scar. Nature 2016, 532, 182–183. [Google Scholar] [CrossRef] [Green Version]
- Faulkner, J.R.; Herrmann, J.E.; Woo, M.J.; Tansey, K.E.; Doan, N.B.; Sofroniew, M.V. Reactive astrocytes protect tissue and preserve function after spinal cord injury. J. Neurosci. 2004, 24, 2143–2155. [Google Scholar] [CrossRef] [Green Version]
- Ge, M.H.; Tian, H.; Mao, L.; Li, D.Y.; Lin, J.Q.; Hu, H.S.; Huang, S.C.; Zhang, C.J.; Mei, X.F. Zinc attenuates ferroptosis and promotes functional recovery in contusion spinal cord injury by activating Nrf2/GPX4 defense pathway. CNS Neurosci. Ther. 2021, 27, 1023–1040. [Google Scholar] [CrossRef]
- Tamaru, T.; Kobayakawa, K.; Saiwai, H.; Konno, D.; Kijima, K.; Yoshizaki, S.; Hata, K.; Iura, H.; Ono, G.; Haruta, Y.; et al. Glial scar survives until the chronic phase by recruiting scar-forming astrocytes after spinal cord injury. Exp. Neurol. 2023, 359, 114264. [Google Scholar] [CrossRef]
- Wang, R.; Zhou, R.; Chen, Z.; Gao, S.; Zhou, F. The glial cells respond to spinal cord injury. Front. Neurol. 2022, 13, 844497. [Google Scholar] [CrossRef]
- Eroglu, C.; Barres, B.A. Regulation of synaptic connectivity by glia. Nature 2010, 468, 223–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhoog, Q.P.; Holtman, L.; Aronica, E.; van Vliet, E.A. Astrocytes as guardians of neuronal excitability: Mechanisms underlying epileptogenesis. Front. Neurol. 2020, 11, 591690. [Google Scholar] [CrossRef] [PubMed]
- Hayatdavoudi, P.; Hosseini, M.; Hajali, V.; Hosseini, A.; Rajabian, A. The role of astrocytes in epileptic disorders. Physiol. Rep. 2022, 10, e15239. [Google Scholar] [CrossRef] [PubMed]
- Popovich, P.G.; Horner, P.J.; Mullin, B.B.; Stokes, B.T. A quantitative spatial analysis of the blood-spinal cord barrier. I. Permeability changes after experimental spinal contusion injury. Exp. Neurol. 1996, 142, 258–275. [Google Scholar] [CrossRef] [PubMed]
- Didangelos, A.; Puglia, M.; Iberl, M.; Sanchez-Bellot, C.; Roschitzki, B.; Bradbury, E.J. High-throughput proteomics reveal alarmins as amplifiers of tissue pathology and inflammation after spinal cord injury. Sci. Rep. 2016, 6, 21607. [Google Scholar] [CrossRef] [Green Version]
- Hackett, A.R.; Lee, J.K. Understanding the NG2 glial scar after spinal cord injury. Front. Neurol. 2016, 7, 199. [Google Scholar] [CrossRef] [Green Version]
- Pizzorusso, T.; Medini, P.; Berardi, N.; Chierzi, S.; Fawcett, J.W.; Maffei, L. Reactivation of ocular dominance plasticity in the adult visual cortex. Science 2002, 298, 1248–1251. [Google Scholar] [CrossRef] [Green Version]
- Schachtrup, C.; Ryu, J.K.; Helmrick, M.J.; Vagena, E.; Galanakis, D.K.; Degen, J.L.; Margolis, R.U.; Akassoglou, K. Fibrinogen triggers astrocyte scar formation by promoting the availability of active TGF-beta after vascular damage. J. Neurosci. 2010, 30, 5843–5854. [Google Scholar] [CrossRef] [Green Version]
- Jia, Z.; Zhu, H.; Li, J.; Wang, X.; Misra, H.; Li, Y. Oxidative stress in spinal cord injury and antioxidant-based intervention. Spinal Cord. 2012, 50, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Coyoy-Salgado, A.; Segura-Uribe, J.J.; Guerra-Araiza, C.; Orozco-Suarez, S.; Salgado-Ceballos, H.; Feria-Romero, I.A.; Gallardo, J.M.; Orozco-Barrios, C.E. The importance of natural antioxidants in the treatment of spinal cord injury in animal models: An overview. Oxid. Med. Cell Longev. 2019, 2019, 3642491. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Yao, X.; Jiang, W.; Li, W.; Zhu, S.; Liao, C.; Zou, L.; Ding, R.; Chen, J. Advanced oxidation protein products induce microglia-mediated neuroinflammation via MAPKs-NF-kappaB signaling pathway and pyroptosis after secondary spinal cord injury. J. Neuroinflamm. 2020, 17, 90. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M.; Kohro, Y.; Yano, T.; Tsujikawa, T.; Kitano, J.; Tozaki-Saitoh, H.; Koyanagi, S.; Ohdo, S.; Ji, R.R.; Salter, M.W.; et al. JAK-STAT3 pathway regulates spinal astrocyte proliferation and neuropathic pain maintenance in rats. Brain 2011, 134, 1127–1139. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, E.; Rivat, C.; Pommier, B.; Mauborgne, A.; Pohl, M. JAK/STAT3 pathway is activated in spinal cord microglia after peripheral nerve injury and contributes to neuropathic pain development in rat. J. Neurochem. 2008, 107, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Stipursky, J.; Gomes, F.C. TGF-beta1/SMAD signaling induces astrocyte fate commitment in vitro: Implications for radial glia development. Glia 2007, 55, 1023–1033. [Google Scholar] [CrossRef] [PubMed]
- Susarla, B.T.; Laing, E.D.; Yu, P.; Katagiri, Y.; Geller, H.M.; Symes, A.J. Smad proteins differentially regulate transforming growth factor-beta-mediated induction of chondroitin sulfate proteoglycans. J. Neurochem. 2011, 119, 868–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, D.O.; Kim, H.; Holl, D.; Werne Solnestam, B.; Lundeberg, J.; Carlen, M.; Goritz, C.; Frisen, J. Reducing pericyte-derived scarring promotes recovery after spinal cord injury. Cell 2018, 173, 153–165.e122. [Google Scholar] [CrossRef] [Green Version]
- Guimaraes-Camboa, N.; Cattaneo, P.; Sun, Y.; Moore-Morris, T.; Gu, Y.; Dalton, N.D.; Rockenstein, E.; Masliah, E.; Peterson, K.L.; Stallcup, W.B.; et al. Pericytes of multiple organs do not behave as mesenchymal stem cells in vivo. Cell Stem Cell 2017, 20, 345–359.e345. [Google Scholar] [CrossRef] [Green Version]
- Hellal, F.; Hurtado, A.; Ruschel, J.; Flynn, K.C.; Laskowski, C.J.; Umlauf, M.; Kapitein, L.C.; Strikis, D.; Lemmon, V.; Bixby, J.; et al. Microtubule stabilization reduces scarring and causes axon regeneration after spinal cord injury. Science 2011, 331, 928–931. [Google Scholar] [CrossRef] [Green Version]
- Okada, M.; Miyamoto, O.; Shibuya, S.; Zhang, X.; Yamamoto, T.; Itano, T. Expression and role of type I collagen in a rat spinal cord contusion injury model. Neurosci. Res. 2007, 58, 371–377. [Google Scholar] [CrossRef]
- Miao, Q.L.; Ye, Q.; Zhang, X.H. Perineuronal net, CSPG receptor and their regulation of neural plasticity. Sheng Li Xue Bao 2014, 66, 387–397. [Google Scholar]
- Irvine, S.F.; Kwok, J.C.F. Perineuronal nets in spinal motoneurones: Chondroitin sulphate proteoglycan around alpha motoneurones. Int. J. Mol. Sci. 2018, 19, 1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussein, R.K.; Mencio, C.P.; Katagiri, Y.; Brake, A.M.; Geller, H.M. Role of chondroitin sulfation following spinal cord injury. Front. Cell. Neurosci. 2020, 14, 208. [Google Scholar] [CrossRef]
- Lang, B.T.; Cregg, J.M.; DePaul, M.A.; Tran, A.P.; Xu, K.; Dyck, S.M.; Madalena, K.M.; Brown, B.P.; Weng, Y.L.; Li, S.; et al. Modulation of the proteoglycan receptor PTPsigma promotes recovery after spinal cord injury. Nature 2015, 518, 404–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, N.; Nandi, S.; Garg, S.; Ghosh, S.; Ghosh, S.; Samat, R.; Ghosh, S. Targeting chondroitin sulfate proteoglycans: An emerging therapeutic strategy to treat CNS injury. ACS Chem. Neurosci. 2020, 11, 231–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharifi, A.; Zandieh, A.; Behroozi, Z.; Hamblin, M.R.; Mayahi, S.; Yousefifard, M.; Ramezani, F. Sustained delivery of chABC improves functional recovery after a spine injury. BMC Neurosci. 2022, 23, 60. [Google Scholar] [CrossRef]
- Kosuri, S.; Borca, C.H.; Mugnier, H.; Tamasi, M.; Patel, R.A.; Perez, I.; Kumar, S.; Finkel, Z.; Schloss, R.; Cai, L.; et al. Machine-assisted discovery of chondroitinase ABC complexes toward sustained neural regeneration. Adv. Healthc. Mater. 2022, 11, e2102101. [Google Scholar] [CrossRef]
- Danilov, C.A.; Steward, O. Conditional genetic deletion of PTEN after a spinal cord injury enhances regenerative growth of CST axons and motor function recovery in mice. Exp. Neurol. 2015, 266, 147–160. [Google Scholar] [CrossRef] [Green Version]
- Park, K.K.; Liu, K.; Hu, Y.; Smith, P.D.; Wang, C.; Cai, B.; Xu, B.; Connolly, L.; Kramvis, I.; Sahin, M.; et al. Promoting axon regeneration in the adult CNS by modulation of the PTEN/mTOR pathway. Science 2008, 322, 963–966. [Google Scholar] [CrossRef] [Green Version]
- Tyler, W.A.; Gangoli, N.; Gokina, P.; Kim, H.A.; Covey, M.; Levison, S.W.; Wood, T.L. Activation of the mammalian target of rapamycin (mTOR) is essential for oligodendrocyte differentiation. J. Neurosci. 2009, 29, 6367–6378. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Stoica, B.A.; Dinizo, M.; Pajoohesh-Ganji, A.; Piao, C.; Faden, A.I. Delayed cell cycle pathway modulation facilitates recovery after spinal cord injury. Cell Cycle 2012, 11, 1782–1795. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.Y.; Kwon, M.J.; Lee, E.M.; Kim, K.; Oh, Y.J.; Kim, H.S.; Hwang, D.H.; Kim, B.G. Role of Myc proto-oncogene as a transcriptional hub to regulate the expression of regeneration-associated genes following preconditioning peripheral nerve injury. J. Neurosci. 2021, 41, 446–460. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.A.; Patel, K.D.; Goncalves, C.M.; Grutzendler, J.; Nishiyama, A. Modulation of oligodendrocyte generation during a critical temporal window after NG2 cell division. Nat. Neurosci. 2014, 17, 1518–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.H.; Chen, J.; Zhou, J.; Nong, F.; Lv, J.H.; Liu, J. Reduced inflammatory cell recruitment and tissue damage in spinal cord injury by acellular spinal cord scaffold seeded with mesenchymal stem cells. Exp. Ther. Med. 2017, 13, 203–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Q.S.; Zhuang, D.L.; Nasser, M.I.; Sai, X.; Deng, G.; Li, G.; Zhu, P. Stem cell therapies for restorative treatments of central nervous system ischemia-reperfusion injury. Cell. Mol. Neurobiol. 2023, 43, 491–510. [Google Scholar] [CrossRef]
- Davies, J.E.; Huang, C.; Proschel, C.; Noble, M.; Mayer-Proschel, M.; Davies, S.J. Astrocytes derived from glial-restricted precursors promote spinal cord repair. J. Biol. 2006, 5, 7. [Google Scholar] [CrossRef] [Green Version]
- Kumamaru, H.; Kadoya, K.; Adler, A.F.; Takashima, Y.; Graham, L.; Coppola, G.; Tuszynski, M.H. Generation and post-injury integration of human spinal cord neural stem cells. Nat. Methods 2018, 15, 723–731. [Google Scholar] [CrossRef]
- Kumamaru, H.; Lu, P.; Rosenzweig, E.S.; Kadoya, K.; Tuszynski, M.H. Regenerating corticospinal axons innervate phenotypically appropriate neurons within neural stem cell grafts. Cell Rep. 2019, 26, 2329–2339. [Google Scholar] [CrossRef] [Green Version]
- Fischer, I.; Dulin, J.N.; Lane, M.A. Transplanting neural progenitor cells to restore connectivity after spinal cord injury. Nat. Rev. Neurosci. 2020, 21, 366–383. [Google Scholar] [CrossRef]
- Salegio, E.A.; Bresnahan, J.C.; Sparrey, C.J.; Camisa, W.; Fischer, J.; Leasure, J.; Buckley, J.; Nout-Lomas, Y.S.; Rosenzweig, E.S.; Moseanko, R.; et al. A unilateral cervical spinal cord contusion injury model in non-human primates (Macaca mulatta). J. Neurotrauma 2016, 33, 439–459. [Google Scholar] [CrossRef] [Green Version]
- Brock, J.H.; Graham, L.; Staufenberg, E.; Im, S.; Tuszynski, M.H. Rodent neural progenitor cells support functional recovery after cervical spinal cord contusion. J. Neurotrauma 2018, 35, 1069–1078. [Google Scholar] [CrossRef]
- Rosenzweig, E.S.; Brock, J.H.; Lu, P.; Kumamaru, H.; Salegio, E.A.; Kadoya, K.; Weber, J.L.; Liang, J.J.; Moseanko, R.; Hawbecker, S.; et al. Restorative effects of human neural stem cell grafts on the primate spinal cord. Nat. Med. 2018, 24, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagoshi, N.; Okano, H.; Nakamura, M. Regenerative therapy for spinal cord injury using iPSC technology. Inflamm. Regen. 2020, 40, 40. [Google Scholar] [CrossRef]
- Lu, P.; Woodruff, G.; Wang, Y.; Graham, L.; Hunt, M.; Wu, D.; Boehle, E.; Ahmad, R.; Poplawski, G.; Brock, J.; et al. Long-distance axonal growth from human induced pluripotent stem cells after spinal cord injury. Neuron 2014, 83, 789–796. [Google Scholar] [CrossRef] [Green Version]
- Doulames, V.M.; Plant, G.W. Induced pluripotent stem cell therapies for cervical spinal cord injury. Int. J. Mol. Sci. 2016, 17, 530. [Google Scholar] [CrossRef] [Green Version]
- Linaro, D.; Vermaercke, B.; Iwata, R.; Ramaswamy, A.; Libe-Philippot, B.; Boubakar, L.; Davis, B.A.; Wierda, K.; Davie, K.; Poovathingal, S.; et al. Xenotransplanted human cortical neurons reveal species-specific development and functional integration into mouse visual circuits. Neuron 2019, 104, 972–986. [Google Scholar] [CrossRef] [Green Version]
- Ceto, S.; Sekiguchi, K.J.; Takashima, Y.; Nimmerjahn, A.; Tuszynski, M.H. Neural stem cell grafts form extensive synaptic networks that integrate with host circuits after spinal cord injury. Cell Stem Cell 2020, 27, 430–440.e435. [Google Scholar] [CrossRef]
- Kawai, M.; Imaizumi, K.; Ishikawa, M.; Shibata, S.; Shinozaki, M.; Shibata, T.; Hashimoto, S.; Kitagawa, T.; Ago, K.; Kajikawa, K.; et al. Long-term selective stimulation of transplanted neural stem/progenitor cells for spinal cord injury improves locomotor function. Cell Rep. 2021, 37, 110019. [Google Scholar] [CrossRef]
- Kitagawa, T.; Nagoshi, N.; Kamata, Y.; Kawai, M.; Ago, K.; Kajikawa, K.; Shibata, R.; Sato, Y.; Imaizumi, K.; Shindo, T.; et al. Modulation by DREADD reveals the therapeutic effect of human iPSC-derived neuronal activity on functional recovery after spinal cord injury. Stem Cell Rep. 2022, 17, 127–142. [Google Scholar] [CrossRef]
- Frantz, S. Embryonic stem cell pioneer Geron exits field, cuts losses. Nat. Biotechnol. 2012, 30, 12–13. [Google Scholar] [CrossRef] [PubMed]
- Gazdic, M.; Volarevic, V.; Harrell, C.R.; Fellabaum, C.; Jovicic, N.; Arsenijevic, N.; Stojkovic, M. Stem cells therapy for spinal cord injury. Int. J. Mol. Sci. 2018, 19, 1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cofano, F.; Boido, M.; Monticelli, M.; Zenga, F.; Ducati, A.; Vercelli, A.; Garbossa, D. Mesenchymal stem cells for spinal cord injury: Current options, limitations, and future of cell therapy. Int. J. Mol. Sci. 2019, 20, 2698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, J.M.; Barry, F.P.; Murphy, J.M.; Mahon, B.P. Mesenchymal stem cells avoid allogeneic rejection. J. Inflamm. 2005, 2, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, J.; Zhang, H. Roles of Mesenchymal stem cells in spinal cord injury. Stem Cells Int. 2017, 2017, 5251313. [Google Scholar] [CrossRef] [Green Version]
- Pelagalli, A.; Nardelli, A.; Lucarelli, E.; Zannetti, A.; Brunetti, A. Autocrine signals increase ovine mesenchymal stem cells migration through Aquaporin-1 and CXCR4 overexpression. J. Cell. Physiol. 2018, 233, 6241–6249. [Google Scholar] [CrossRef] [PubMed]
- Vizoso, F.J.; Eiro, N.; Cid, S.; Schneider, J.; Perez-Fernandez, R. Mesenchymal stem cell secretome: Toward cell-free therapeutic strategies in regenerative medicine. Int. J. Mol. Sci. 2017, 18, 1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mead, B.; Logan, A.; Berry, M.; Leadbeater, W.; Scheven, B.A. Paracrine-mediated neuroprotection and neuritogenesis of axotomised retinal ganglion cells by human dental pulp stem cells: Comparison with human bone marrow and adipose-derived mesenchymal stem cells. PLoS ONE 2014, 9, e109305. [Google Scholar] [CrossRef]
- De Luca, A.; Gallo, M.; Aldinucci, D.; Ribatti, D.; Lamura, L.; D’Alessio, A.; De Filippi, R.; Pinto, A.; Normanno, N. Role of the EGFR ligand/receptor system in the secretion of angiogenic factors in mesenchymal stem cells. J. Cell. Physiol. 2011, 226, 2131–2138. [Google Scholar] [CrossRef]
- Zanotti, L.; Angioni, R.; Cali, B.; Soldani, C.; Ploia, C.; Moalli, F.; Gargesha, M.; D’Amico, G.; Elliman, S.; Tedeschi, G.; et al. Mouse mesenchymal stem cells inhibit high endothelial cell activation and lymphocyte homing to lymph nodes by releasing TIMP-1. Leukemia 2016, 30, 1143–1154. [Google Scholar] [CrossRef]
- El-Kheir, W.A.; Gabr, H.; Awad, M.R.; Ghannam, O.; Barakat, Y.; Farghali, H.A.; El Maadawi, Z.M.; Ewes, I.; Sabaawy, H.E. Autologous bone marrow-derived cell therapy combined with physical therapy induces functional improvement in chronic spinal cord injury patients. Cell Transplant. 2014, 23, 729–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, G.; Liu, X.; Zhang, Z.; Yang, Z.; Dai, Y.; Xu, R. Transplantation of autologous bone marrow mesenchymal stem cells in the treatment of complete and chronic cervical spinal cord injury. Brain Res. 2013, 1533, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Geffner, L.F.; Santacruz, P.; Izurieta, M.; Flor, L.; Maldonado, B.; Auad, A.H.; Montenegro, X.; Gonzalez, R.; Silva, F. Administration of autologous bone marrow stem cells into spinal cord injury patients via multiple routes is safe and improves their quality of life: Comprehensive case studies. Cell Transplant. 2008, 17, 1277–1293. [Google Scholar] [CrossRef] [Green Version]
- Mendonca, M.V.; Larocca, T.F.; de Freitas Souza, B.S.; Villarreal, C.F.; Silva, L.F.; Matos, A.C.; Novaes, M.A.; Bahia, C.M.; de Oliveira Melo Martinez, A.C.; Kaneto, C.M.; et al. Safety and neurological assessments after autologous transplantation of bone marrow mesenchymal stem cells in subjects with chronic spinal cord injury. Stem Cell Res. Ther. 2014, 5, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karamouzian, S.; Nematollahi-Mahani, S.N.; Nakhaee, N.; Eskandary, H. Clinical safety and primary efficacy of bone marrow mesenchymal cell transplantation in subacute spinal cord injured patients. Clin. Neurol. Neurosurg. 2012, 114, 935–939. [Google Scholar] [CrossRef]
- Deng, J.; Petersen, B.E.; Steindler, D.A.; Jorgensen, M.L.; Laywell, E.D. Mesenchymal stem cells spontaneously express neural proteins in culture and are neurogenic after transplantation. Stem Cells 2006, 24, 1054–1064. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, T.; Ding, J.; Gu, H.; Wang, Q.; Wang, Y.; Zhang, D.; Gao, C. A reactive oxygen species-responsive hydrogel encapsulated with bone marrow derived stem cells promotes repair and regeneration of spinal cord injury. Bioact. Mater. 2023, 19, 550–568. [Google Scholar] [CrossRef]
- Papa, S.; Vismara, I.; Mariani, A.; Barilani, M.; Rimondo, S.; De Paola, M.; Panini, N.; Erba, E.; Mauri, E.; Rossi, F.; et al. Mesenchymal stem cells encapsulated into biomimetic hydrogel scaffold gradually release CCL2 chemokine in situ preserving cytoarchitecture and promoting functional recovery in spinal cord injury. J. Control. Release 2018, 278, 49–56. [Google Scholar] [CrossRef]
- Koffler, J.; Zhu, W.; Qu, X.; Platoshyn, O.; Dulin, J.N.; Brock, J.; Graham, L.; Lu, P.; Sakamoto, J.; Marsala, M.; et al. Biomimetic 3D-printed scaffolds for spinal cord injury repair. Nat. Med. 2019, 25, 263–269. [Google Scholar] [CrossRef]
- Zipser, C.M.; Cragg, J.J.; Guest, J.D.; Fehlings, M.G.; Jutzeler, C.R.; Anderson, A.J.; Curt, A. Cell-based and stem-cell-based treatments for spinal cord injury: Evidence from clinical trials. Lancet Neurol. 2022, 21, 659–670. [Google Scholar] [CrossRef]
- Pinelli, F.; Pizzetti, F.; Veneruso, V.; Petillo, E.; Raghunath, M.; Perale, G.; Veglianese, P.; Rossi, F. Biomaterial-mediated factor delivery for spinal cord injury treatment. Biomedicines 2022, 10, 1673. [Google Scholar] [CrossRef]
- Lu, T.; Peng, W.; Liang, Y.; Li, M.; Li, D.S.; Du, K.H.; Zhu, J.H.; Wu, J.H. PTEN-silencing combined with ChABC-overexpression in adipose-derived stem cells promotes functional recovery of spinal cord injury in rats. Biochem. Biophys. Res. Commun. 2020, 532, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.; Li, Y.; Anderson, J.; Castro-Pedrido, S.; Skinner, R.; Lei, S.; Finkel, Z.; Rodriguez, B.; Esteban, F.; Lee, K.B.; et al. Gsx1 promotes locomotor functional recovery after spinal cord injury. Mol. Ther. 2021, 29, 2469–2482. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Li, Y.; Shen, P.P.; Wang, B. Gene-modified stem cells for spinal cord injury: A promising better alternative therapy. Stem Cell Rev. Rep. 2022, 18, 2662–2682. [Google Scholar] [CrossRef] [PubMed]
- Puls, B.; Ding, Y.; Zhang, F.; Pan, M.; Lei, Z.; Pei, Z.; Jiang, M.; Bai, Y.; Forsyth, C.; Metzger, M.; et al. Regeneration of functional neurons after spinal cord injury via in situ NeuroD1-mediated astrocyte-to-neuron conversion. Front. Cell Dev. Biol. 2020, 8, 591883. [Google Scholar] [CrossRef]
- Su, Z.; Niu, W.; Liu, M.L.; Zou, Y.; Zhang, C.L. In vivo conversion of astrocytes to neurons in the injured adult spinal cord. Nat. Commun. 2014, 5, 3338. [Google Scholar] [CrossRef] [Green Version]
- Tai, W.; Wu, W.; Wang, L.L.; Ni, H.; Chen, C.; Yang, J.; Zang, T.; Zou, Y.; Xu, X.M.; Zhang, C.L. In vivo reprogramming of NG2 glia enables adult neurogenesis and functional recovery following spinal cord injury. Cell Stem Cell 2021, 28, 923–937. [Google Scholar] [CrossRef]
- Li, Y.; Tzatzalos, E.; Kwan, K.Y.; Grumet, M.; Cai, L. Transcriptional regulation of Notch1 expression by Nkx6.1 in neural stem/progenitor cells during ventral spinal cord development. Sci. Rep. 2016, 6, 38665. [Google Scholar] [CrossRef] [Green Version]
- Tzatzalos, E.; Smith, S.M.; Doh, S.T.; Hao, H.; Li, Y.; Wu, A.; Grumet, M.; Cai, L. A cis-element in the Notch1 locus is involved in the regulation of gene expression in interneuron progenitors. Dev. Biol. 2012, 372, 217–228. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.; Anderson, J.; Lei, S.; Finkel, Z.; Rodriguez, B.; Esteban, F.; Risman, R.; Li, Y.; Lee, K.B.; Lyu, Y.L.; et al. Nkx6.1 enhances neural stem cell activation and attenuates glial scar formation and neuroinflammation in the adult injured spinal cord. Exp. Neurol. 2021, 345, 113826. [Google Scholar] [CrossRef]
- Finkel, Z.; Cai, L. Transcription factors promote neural regeneration after spinal cord injury. Neural. Regen. Res. 2022, 17, 2439–2440. [Google Scholar] [CrossRef] [PubMed]
- Brommer, B.; He, M.; Zhang, Z.; Yang, Z.; Page, J.C.; Su, J.; Zhang, Y.; Zhu, J.; Gouy, E.; Tang, J.; et al. Improving hindlimb locomotor function by Non-invasive AAV-mediated manipulations of propriospinal neurons in mice with complete spinal cord injury. Nat. Commun. 2021, 12, 781. [Google Scholar] [CrossRef] [PubMed]
- Schlimgen, R.; Howard, J.; Wooley, D.; Thompson, M.; Baden, L.R.; Yang, O.O.; Christiani, D.C.; Mostoslavsky, G.; Diamond, D.V.; Duane, E.G.; et al. Risks associated with lentiviral vector exposures and prevention strategies. J. Occup. Environ. Med. 2016, 58, 1159–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naso, M.F.; Tomkowicz, B.; Perry, W.L., 3rd; Strohl, W.R. Adeno-Associated Virus (AAV) as a vector for gene therapy. BioDrugs 2017, 31, 317–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kathe, C.; Skinnider, M.A.; Hutson, T.H.; Regazzi, N.; Gautier, M.; Demesmaeker, R.; Komi, S.; Ceto, S.; James, N.D.; Cho, N.; et al. The neurons that restore walking after paralysis. Nature 2022, 611, 540–547. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Positive Effects: | References: |
|---|---|
| Uptakes excess glutamate preventing chronic glutamate neurotoxicity | [70,71] |
| Macrophages improve overall tissue quality through removal of cell debris | [43,55,57] |
| Prolonged tissue repair signaling | [16,36,49] |
| Maintains body’s natural excitation/inhibition ratio; helps prevent irregular signaling | [45,72,73] |
| Physical barrier to protect remaining functional tissue | [14,29,37] |
| Negative Effects: | |
| Non-resolving auto-immune response that leads to fueling fibrosis; over stimulation of inflammation leads to damaged surrounding tissues | [16,74] |
| Over stimulation of inflammation leads to damaged surrounding tissues | [16,50,75] |
| Inhibits differentiation of OPCs | [26,76] |
| Inhibits axogenesis, plasticity of the neuron, and myelin generation | [4,5,6,13,77] |
| Physical barrier making transplanted/endogenous cell migration difficult | [29,35,45,78] |
| Excessive production of free radicals, reactive oxygen species (ROS), and glutamate, as well as ion imbalance | [14,79,80,81] |
| Study Title | Intervention | Duration | Phase | Status | Sponsor |
|---|---|---|---|---|---|
| Intrathecal Transplantation of Autologous Bone Marrow-derived Mononuclear Cells for Treating Traumatic Acute Spinal Cord Injury | Lumbar injection Transplantation of autologous bone-marrow-derived mononuclear cells | 2020–2023 | II | recruiting | Shanghai Changzheng Hospital |
| Assessment of Safety and Effectiveness of Mesenchymal Stem Cells in the Treatment of Spinal Cord Injury (SCI) Patients | Autologous bone-marrow-derived MSCs and Wharton-jelly-derived mesenchymal stem cells | 2017–2020 | I | completed | University of Jordan |
| Subarachnoid Administrations of Adults Autologous Mesenchymal Stromal Cells in SCI | Adult autologous mesenchymal bone marrow cells | 2019 | I | completed | Puerta de Hierro University Hospital |
| Intrathecal Administration (Pattern 100/3) of Expanded Autologous Adult Bone Marrow Mesenchymal Stem Cells in Established Chronic Spinal Cord Injuries | Autologous mesenchymal bone marrow cell injection | 2015–2017 | II | completed | Puerta de Hierro University Hospital |
| Autologous Bone Marrow Cell Transplantation in Persons with Acute Spinal Cord Injury—An Indian Pilot Study | Autologous mesenchymal bone marrow cells | 2011–2017 | I/II | completed | Indian Spinal Injuries Centre |
| Comparative Evaluation of Safety and Effectiveness of Autologous Bone Marrow Derived Mesenchymal Stem Cells (BM-MSC) vs. Adipose Tissue Derived Mesenchymal Stem Cells (AT-MSC) in the Treatment of Spinal Cord Injury (SCI) Patient | Intrathecal injection of autologous mesenchymal stem cells | 2016–2018 | I/II | completed | University of Jordan |
| CELLTOP Part II: A Phase II Clinical Trial of Autologous Adipose Derived Mesenchymal Stem Cells in the Treatment of Paralysis Due to Traumatic Spinal Cord Injury | Intrathecal transplantation of autologous adipose-derived mesenchymal stem cells | 2020–2024 | II | recruiting | Mayo Clinic |
| Phase I Clinical Trial of Autologous Adipose Derived Mesenchymal Stem Cells in the Treatment of Paralysis Due to Traumatic Spinal Cord Injury | Intrathecal delivery of autologous, adipose-derived mesenchymal stem cells | 2017–2019 | I | completed | Mohamad Bydon |
| Safety of Cultured Allogeneic Adult Umbilical Cord Derived Mesenchymal Stem Cells for SCI | Cultured allogeneic adult umbilical-cord-derived mesenchymal stem cells | 2022–2026 | I | recruiting | Foundation for Orthopaedics and Regenerative Medicine |
| A Randomized Controlled Phase II, Two-Arm Study of Umbilical Cord Blood Cell Transplant (MC001) Into Injured Spinal Cord Followed by the Locomotor Training for Patients with Chronic Complete Spinal Cord Injuries (SCI) | Umbilical cord blood mononuclear stem cell (UCBMSC) transplant | 2022–2023 | II | recruiting | StemCyte, Inc. |
| Allogeneic Cord Blood for Neurological Diseases in Adults | Allogeneic umbilical cord blood therapy | 2022 | I | Not yet recruiting | The Medical Pavilion, Bahamas |
| Repeated Subarachnoid Administrations of Human Umbilical Cord Mesenchymal Stem Cells in Treating Spinal Cord Injury | Intrathecal administration of human umbilical cord mesenchymal stem cells | 2018–2020 | I/II | completed | Limin Rong, Third Affiliated Hospital, Sun Yat-Sen University |
| Allogeneic Mononuclear Umbilical Cord Blood Systemic Infusions for Adult Patients with Severe Acute Contusion Spinal Cord Injury: Phase I Safety Study and Phase IIa Primary Efficiency Study | I.V. infusions of human allogeneic umbilical cord blood mononuclear cells | 2013–2018 | I/IIa | completed | Sklifosovsky Institute of Emergency Care |
| A Single Center, Open Label, Single Group, Phase 1/2a Clinical Study to Evaluate the Safety and Exploratory Efficacy of Transplantation Therapy Using PSA-NCAM(+) NPC Derived From hESC Line in AIS-A Level of Sub-acute SCI(From 7 to 60 Days) | Neural precursor cells derived from human embryonic stem cell line | 2021–2023 | I/IIa | recruiting | S.Biomedics Co., Ltd. |
| A Multi-center, Double-blind, Randomized, Placebo-controlled, Delayed Start Phase II/III Study to Assess the Efficacy and Safety of Neuro-Cells in (Sub)Acute Spinal Cord Injury Patients | Intrathecal intervention with neuro-cells | 2022 | II/III | recruiting | Neuroplast |
| A 3 Months Open Phase I Study to Assess the Safety of the Intrathecal Application of Neuro-Cells in End Stage (Chronic) Traumatic Spinal Cord Injury Patients | Intrathecal application of neuro-cells | 2020–2021 | I | active, not recruiting | Neuroplast |
| The Safety of Autologous Human Schwann Cells (ahSC) in Subjects with Chronic Spinal Cord Injury (SCI) Receiving Rehabilitation | Autologous human Schwann cell transplantation | 2015–2019 | I | completed | W. Dalton Dietrich, University of Miami, Miami Project |
| Dose Escalation Study of AST-OPC1 in Spinal Cord Injury | AST-OPC1 injection | 2015–2018 | I/IIa | completed | Lineage Cell Therapeutics, Inc. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clifford, T.; Finkel, Z.; Rodriguez, B.; Joseph, A.; Cai, L. Current Advancements in Spinal Cord Injury Research—Glial Scar Formation and Neural Regeneration. Cells 2023, 12, 853. https://doi.org/10.3390/cells12060853
Clifford T, Finkel Z, Rodriguez B, Joseph A, Cai L. Current Advancements in Spinal Cord Injury Research—Glial Scar Formation and Neural Regeneration. Cells. 2023; 12(6):853. https://doi.org/10.3390/cells12060853
Chicago/Turabian StyleClifford, Tanner, Zachary Finkel, Brianna Rodriguez, Adelina Joseph, and Li Cai. 2023. "Current Advancements in Spinal Cord Injury Research—Glial Scar Formation and Neural Regeneration" Cells 12, no. 6: 853. https://doi.org/10.3390/cells12060853