
Targeting Underlying Inflammation in Carcinoma Is Essential for the Resolution of Depressiveness

 , , , , and
, , , , and
Abstract
:1. Introduction
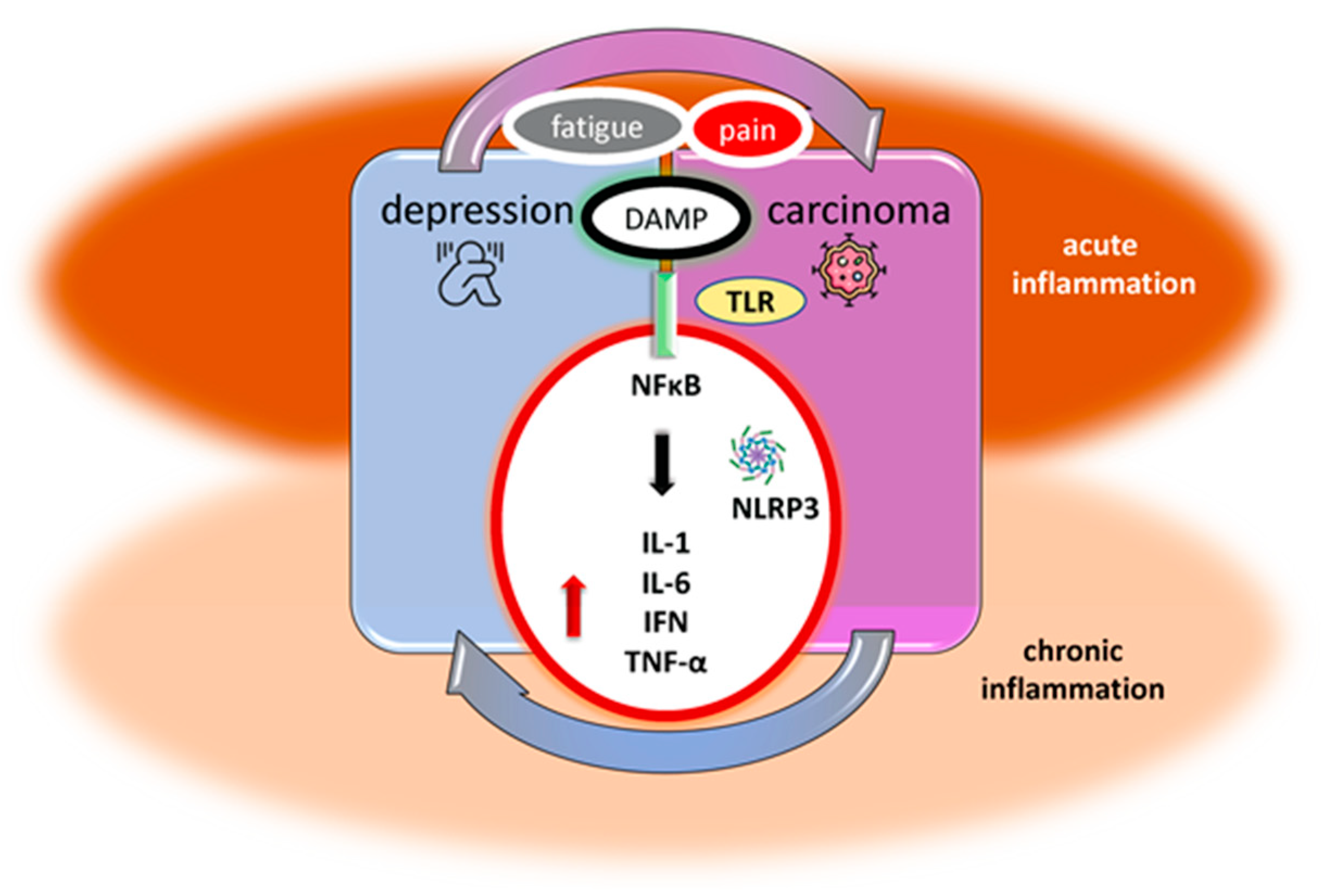
2. Acute and Chronic Inflammation in Carcinoma
3. Acute and Chronic Inflammation in Depression
4. Animal Models of Inflammatory-Induced Depression in Carcinoma
5. Underlying Inflammatory Disturbances in Carcinoma and Depression Co-Occurrence
6. Potential for New Anti-Inflammatory Strategies in Cooccurrence of Carcinoma and Depression
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uchikawa, H.; Fujii, K.; Shiohama, T.; Nakazato, M.; Shimizu, E.; Miyashita, T.; Shimojo, N. Specific temperament in patients with nevoid basal cell carcinoma syndrome. Pediatr. Int. 2021, 63, 177–182. [Google Scholar] [CrossRef]
- Verma, M.; Paik, J.M.; Younossi, I.; Tan, D.; Abdelaal, H.; Younossi, Z.M. The impact of hepatocellular carcinoma diagnosis on patients’ health-related quality of life. Cancer Med. 2021, 10, 6273–6281. [Google Scholar] [CrossRef]
- Pham, H.; Torres, H.; Sharma, P. Mental health implications in bladder cancer patients: A review. Urol. Oncol. 2019, 37, 97–107. [Google Scholar] [CrossRef]
- Oh, H.M.; Son, C.G. The Risk of Psychological Stress on Cancer Recurrence: A Systematic Review. Cancers 2021, 13, 5816. [Google Scholar] [CrossRef] [PubMed]
- Kelley, K.W.; Bluthé, R.M.; Dantzer, R.; Zhou, J.H.; Shen, W.H.; Johnson, R.W.; Broussard, S.R. Cytokine-induced sickness behavior. Brain Behav. Immun. 2003, 1, 12–118. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Gotoh, N. Inflammatory cytokines-enriched microenvironment plays key roles for the development of breast cancers. Cancer Sci. 2023; epub ahead of print. [Google Scholar] [CrossRef]
- Bauer, M.E.; Teixeira, A.L. Inflammation in psychiatric disorders: What comes first? Ann. N. Y. Acad. Sci. 2019, 1437, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Poole, L.; Dickens, C.; Steptoe, A. The puzzle of depression and acute coronary syndrome: Reviewing the role of acute inflammation. J. Psychosom. Res. 2011, 71, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Beurel, E.; Toups, M.; Nemeroff, C.B. The Bidirectional Relationship of Depression and Inflammation: Double Trouble. Neuron. 2020, 107, 234–256. [Google Scholar] [CrossRef] [PubMed]
- Rohleder, N. Stress and inflammation—The need to address the gap in the transition between acute and chronic stress effects. Psychoneuroendocrinology 2019, 105, 164–171. [Google Scholar] [CrossRef]
- Lu, H.; Ouyang, W.; Huang, C. Inflammation, a key event in cancer development. Mol. Cancer Res. 2006, 4, 221–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freyermuth-Trujillo, X.; Segura-Uribe, J.J.; Salgado-Ceballos, H.; Orozco-Barrios, C.E.; Coyoy-Salgado, A. Inflammation: A Target for Treatment in Spinal Cord Injury. Cells 2022, 11, 2692. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.; Ward, P.; Gilroy, D. Fundamentals of Inflammation; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar] [CrossRef] [Green Version]
- Munn, L. Cancer and Inflammation. Wiley Interdiscip. Rev. Syst. Biol. Med. 2017, 9, e1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matzinger, P. Tolerance, danger, and the extended family. Annu. Rev. Immunol. 1994, 12, 991–1045. [Google Scholar] [CrossRef]
- Gallucci, S.; Matzinger, P. Danger signals: SOS to the immune system. Curr. Opin. Immunol. 2001, 13, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Han, Z.; Oppenheim, J.J. Alarmins and immunity. Immunol. Rev. 2017, 280, 41–56. [Google Scholar] [CrossRef]
- Chalmers, S.; Khawaja, A.; Wieruszewski, P.M.; Gajic, O.; Odeyemi, Y. Diagnosis and treatment of acute pulmonary inflammation in critically ill patients: The role of inflammatory biomarkers. World J. Crit. Care Medl. 2019, 8, 59–71. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Abbas, A.K.; Aster, J.C. Robbins Basic Pathology; Elsevier Health Sciences: Philadephia, PA, USA, 2013. [Google Scholar]
- Dinarello, C.A. Proinflammatory cytokines. Chest 2000, 118, 503–508. [Google Scholar] [CrossRef]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.M.; An, J. Cytokines, inflammation and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Philip, M.; Rowley, D.A.; Schreiber, H. Inflammation as a tumor promoter in cancer induction. Semin. Cancer Biol. 2004, 14, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Korniluk, A.; Koper, O.; Kemona, H.; Dymicka-Piekarska, V. From inflammation to cancer. Ir. J. Med. Sci. 2017, 186, 57–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef]
- Jovanovic, I.P.; Pejnovic, N.N.; Radosavljevic, G.D.; Arsenijevic, N.N.; Lukic, M.L. IL-33/ST2 axis in innate and acquired immunity to tumors. Oncoimmunology 2012, 1, 229–231. [Google Scholar] [CrossRef] [Green Version]
- Pollard, J.W. Tumour-educated macrophages promote tumour progression and metastasis. Nat. Rev. Cancer 2004, 4, 71–78. [Google Scholar] [CrossRef]
- Schetter, A.J.; Heegaard, N.H.; Harris, C.C. Inflammation and cancer: Interweaving microRNA, free radical, cytokine and p53 pathways. Carcinogenesis 2010, 31, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Nettis, M.A.; Pariante, C.M. Is there neuroinflammation in depression? Understanding the link between the brain and the peripheral immune system in depression. Int. Rev. Neurobiol. 2020, 152, 23–40. [Google Scholar] [CrossRef]
- Carlessi, A.S.; Borba, L.A.; Zugno, A.I.; Quevedo, J.; Réus, G.Z. Gut microbiota-brain axis in depression: The role of neuroinflammation. Eur. J. Neurosci. 2021, 53, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Majd, M.; Saunders, E.F.H.; Engeland, C.G. Inflammation and the dimensions of depression: A review. Front. Neuroendocrinol. 2020, 56, 100800. [Google Scholar] [CrossRef] [PubMed]
- Capuron, L.; Gumnick, J.F.; Musselman, D.L.; Lawson, D.H.; Reemsnyder, A.; Nemeroff, C.B.; Miller, A.H. Neurobehavioral effects of interferon-alpha in cancer patients: Phenomenology and paroxetine responsiveness of symptom dimensions. Neuropsychopharmacology 2002, 26, 643–652. [Google Scholar] [CrossRef] [PubMed]
- Loftis, J.M.; Huckans, M.; Morasco, B.J. Neuroimmune mechanisms of cytokine-induced depression: Current theories and novel treatment strategies. Neurobiol. Dis. 2010, 37, 519–533. [Google Scholar] [CrossRef] [Green Version]
- Lynall, M.E.; Turner, L.; Bhatti, J.; Cavanagh, J.; de Boer, P.; Mondelli, V.; Jones, D.; Drevets, W.C.; Cowen, P.; Harrison, N.A.; et al. Neuroimmunology of Mood Disorders and Alzheimer’s Disease (NIMA) Consortium. Peripheral Blood Cell-Stratified Subgroups of Inflamed Depression. Biol. Psychiatry 2020, 88, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Borovcanin, M.M.; Vesic, K.; Balcioglu, Y.H.; Mijailović, N.R. Prescription of selective serotonin reuptake inhibitors in COVID-19 infection needs caution. Front. Psychiatry 2022, 13, 1052710. [Google Scholar] [CrossRef]
- Dantzer, R.; O’Connor, J.C.; Freund, G.G.; Johnson, R.W.; Kelley, K.W. From inflammation to sickness and depression: When the immune system subjugates the brain. Nat. Rev. Neurosci. 2008, 9, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Kiraly, D.D.; Horn, S.R.; Van Dam, N.T.; Costi, S.; Schwartz, J.; Kim-Schulze, S.; Patel, M.; Hodes, G.E.; Russo, S.J.; Merad, M.; et al. Altered peripheral immune profiles in treatment-resistant depression: Response to ketamine and prediction of treatment outcome. Transl. Psychiatry 2017, 7, e1065. [Google Scholar] [CrossRef] [Green Version]
- Dudek, K.A.; Dion-Albert, L.; Kaufmann, F.N.; Tuck, E.; Lebel, M.; Menard, C. Neurobiology of resilience in depression: Immune and vascular insights from human and animal studies. Eur. J. Neurosci. 2021, 53, 183–221. [Google Scholar] [CrossRef] [Green Version]
- Mac Giollabhui, N.; Ng, T.H.; Ellman, L.M.; Alloy, L.B. The longitudinal associations of inflammatory biomarkers and depression revisited: Systematic review, meta-analysis, and meta-regression. Mol. Psychiatry 2021, 26, 3302–3314. [Google Scholar] [CrossRef]
- Del Giudice, M.; Gangestad, S.W. Rethinking IL-6 and CRP: Why they are more than inflammatory biomarkers, and why it matters. Brain Behav. Immun. 2018, 70, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Vegas, O.; Beitia, G.; Sánchez-Martin, J.R.; Arregi, A.; Azpiroz, A. Behavioral and neurochemical responses in mice bearing tumors submitted to social stress. Behav. Brain Res. 2004, 155, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, W.; Lu, T.; Li, J.; Zheng, Y.; Kong, L. Morphological and genetic characterization of a cultivated Cordyceps sinensis fungus and its polysaccharide component possessing antioxidant property in H22 tumor-bearing mice. Life Sci. 2006, 78, 2742–2748. [Google Scholar] [CrossRef]
- Papiez, M.A.; Dybala, M.; Sowa-Kucma, M.; Krzysciak, W.; Taha, H.; Jozkowicz, A.; Nowak, G. Evaluation of oxidative status and depression-like responses in Brown Norway rats with acute myeloid leukemia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2009, 33, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.; Ma, J.; Liu, Y.M.; Yang, L.; Peng, L.; Wang, H.; Chen, H.Z. Allostatic tumor-burden induces depression-associated changes in hepatoma-bearing mice. J. Neuro-Oncol. 2009, 94, 367–372. [Google Scholar] [CrossRef]
- Nashed, M.G.; Seidlitz, E.P.; Frey, B.N.; Singh, G. Depressive-like behaviours and decreased dendritic branching in the medial prefrontal cortex of mice with tumors: A novel validated model of cancer-induced depression. Behav. Brain Res. 2015, 294, 25–35. [Google Scholar] [CrossRef]
- Lamkin, D.M.; Lutgendorf, S.K.; Lubaroff, D.; Sood, A.K.; Beltz, T.G.; Johnson, A.K. Cancer induces inflammation and depressive-like behavior in the mouse: Modulation by social housing. Brain Behav. Immun. 2011, 25, 555–564. [Google Scholar] [CrossRef] [Green Version]
- Lebeña, A.; Vegas, O.; Gómez-Lázaro, E.; Arregi, A.; Garmendia, L.; Beitia, G.; Azpiroz, A. Melanoma tumors alter proinflammatory cytokine production and monoamine brain function, and induce depressive-like behavior in male mice. Behav. Brain Res. 2014, 272, 83–92. [Google Scholar] [CrossRef]
- Fang, C.K.; Chen, H.W.; Chiang, I.T.; Chen, C.C.; Liao, J.F.; Su, T.P.; Hwang, J.J. Mirtazapine inhibits tumor growth via immune response and serotonergic system. PLoS ONE 2012, 7, e38886. [Google Scholar] [CrossRef]
- Pyter, L.M.; Pineros, V.; Galang, J.A.; McClintock, M.K.; Prendergast, B.J. Peripheral tumors induce depressive-like behaviors and cytokine production and alter hypothalamic-pituitary-adrenal axis regulation. Proc. Natl. Acad. Sci. USA 2009, 106, 9069–9074. [Google Scholar] [CrossRef] [Green Version]
- Norden, D.M.; Bicer, S.; Clark, Y.; Jing, R.; Henry, C.J.; Wold, L.E.; McCarthy, D.O. Tumor growth increases neuroinflammation, fatigue and depressive-like behavior prior to alterations in muscle function. Brain Behav. Immun. 2014, 43, 76–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, W.; Zhu, H.; Zhang, P.; Sun, T.; Gong, S.; Sun, G.; Ren, Z. Heterotopic 4T1 breast cancer transplantation induces hippocampal inflammation and depressive-like behaviors in mice. Metab. Brain Dis. 2022, 37, 2955–2963. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Zhou, F.; Yuan, X.; Yang, T.; Liang, X.; Wang, Y.; Tu, H.; Chang, J.; Nan, K.; Wei, Y. Reactive oxygen species are involved in the development of gastric cancer and gastric cancer-related depression through ABL1-mediated inflammation signaling pathway. Oxidative Med. Cell Longev. 2019, 2019, 5813985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.P.; Zhang, C.; Zhang, Y.P.; Li, K.W.; Song, C. The combination of chronic stress and smoke exacerbated depression-like changes and lung cancer factor expression in A/J mice: Involve inflammation and BDNF dysfunction. PLoS ONE 2022, 17, e0277945. [Google Scholar] [CrossRef] [PubMed]
- Almutabagani, L.F.; Almanqour, R.A.; Alsabhan, J.F.; Alhossan, A.M.; Alamin, M.A.; Alrajeh, H.M.; Alonazi, A.S.; El-Malky, A.M.; Alrasheed, N.M. Inflammation and Treatment-Resistant Depression from Clinical to Animal Study: A Possible Link? Neurol. Int. 2023, 15, 100–120. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.K.; Kavelaars, A.; Heijnen, C.J.; Dantzer, R. Neuroinflammation and comorbidity of pain and depression. Pharmacol. Rev. 2013, 66, 80–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, B.L. Biological basis of the behavior of sick animals. Neurosci. Biobehav. Rev. 1988, 12, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.J.; Chan, M.; Bhatti, H.; Halton, M.; Grassi, L.; Johansen, C.; Meader, N. Prevalence of depression, anxiety, and adjustment disorder in oncological, haematological, and palliative-care settings: A meta-analysis of 94 interview-based studies. Lancet Oncol. 2011, 12, 160–174. [Google Scholar] [CrossRef]
- Sharma, N.; Hansen, C.H.; O’Connor, M.; Thekkumpurath, P.; Walker, J.; Kleiboer, A.; Murray, G.; Espie, C.; Storey, D.; Sharpe, M.; et al. Sleep problems in cancer patients: Prevalence and association with distress and pain. Psychooncology 2012, 21, 1003–1009. [Google Scholar] [CrossRef]
- Miaskowski, C.; Dodd, M.; Lee, K. Symptom clusters: The new frontier in symptom management research. J. Natl. Cancer Inst. Monogr. 2004, 32, 17–21. [Google Scholar] [CrossRef]
- Dodd, M.J.; Miaskowski, C.; Lee, K.A. Occurrence of symptom clusters. J. Natl. Cancer Inst. Monogr. 2004, 32, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Dodd, M.J.; Miaskowski, C.; Paul, S.M. Symptom clusters and their effect on the functional status of patients with cancer. Oncol. Nurs. Forum. 2001, 28, 465–470. [Google Scholar] [PubMed]
- Charalambous, A.; Giannakopoulou, M.; Bozas, E.; Paikousis, L. Parallel and serial mediation analysis between pain, anxiety, depression, fatigue and nausea, vomiting and retching within a randomised controlled trial in patients with breast and prostate cancer. BMJ Open 2019, 9, e026809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holliday, E.B.; Dieckmann, N.F.; McDonald, T.L.; Hung, A.Y.; Thomas, C.R., Jr.; Wood, L.J. Relationship between fatigue, sleep quality and inflammatory cytokines during external beam radiation therapy for prostate cancer: A prospective study. Radiother Oncol. 2016, 118, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Giuliani, F. The Role of Inflammation in Depression and Fatigue. Front. Immunol. 2019, 10, 1696. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, M.; Huh, Y.; Ji, R.R. Roles of inflammation, neurogenic inflammation, and neuroinflammation in pain. J. Anesth. 2019, 33, 131–139. [Google Scholar] [CrossRef]
- Katarina, V.; Gordana, T.; Svetlana, M.D.; Milica, B. Oxidative stress and neuroinflammation should be both considered in the occurrence of fatigue and depression in multiple sclerosis. Acta Neurol. Belg. 2020, 120, 853–861. [Google Scholar] [CrossRef]
- Lynn, M.; Maclachlan, L.; Finkelmeyer, A.; Clark, J.; Locke, J.; Todryk, S.; Ng, W.F.; Newton, J.L.; Watson, S. Reduction of Glucocorticoid Receptor Function in Chronic Fatigue Syndrome. Mediators Inflamm. 2018, 2018, 3972104. [Google Scholar] [CrossRef]
- Thornton, L.M.; Andersen, B.L.; Blakely, W.P. The pain, depression, and fatigue symptom cluster in advanced breast cancer: Covariation with the hypothalamic-pituitary-adrenal axis and the sympathetic nervous system. Health Psychol. 2010, 29, 333–337. [Google Scholar] [CrossRef] [Green Version]
- Lanser, L.; Kink, P.; Egger, E.M.; Willenbacher, W.; Fuchs, D.; Weiss, G.; Kurz, K. Inflammation-Induced Tryptophan Breakdown is Related With Anemia, Fatigue, and Depression in Cancer. Front. Immunol. 2020, 11, 249. [Google Scholar] [CrossRef] [Green Version]
- Sheng, J.; Liu, S.; Wang, Y.; Cui, R.; Zhang, X. The Link between Depression and Chronic Pain: Neural Mechanisms in the Brain. Neural Plast. 2017, 2017, 9724371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwekkeboom, K.L.; Tostrud, L.; Costanzo, E.; Coe, C.L.; Serlin, R.C.; Ward, S.E.; Zhang, Y. The Role of Inflammation in the Pain, Fatigue, and Sleep Disturbance Symptom Cluster in Advanced Cancer. J. Pain Symptom Manage. 2018, 55, 1286–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bamonti, P.M.; Moye, J.; Naik, A.D. Pain is associated with continuing depression in cancer survivors. Psychol. Health Med. 2018, 23, 1182–1195. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Dantzer, R.; Budac, D.P.; Walker, A.K.; Mao-Ying, Q.L.; Lee, A.W.; Heijnen, C.J.; Kavelaars, A. Peripheral indoleamine 2,3-dioxygenase 1 is required for comorbid depression-like behavior but does not contribute to neuropathic pain in mice. Brain Behav. Immun. 2015, 46, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Jovanovic, F.; Candido, K.D.; Knezevic, N.N. The Role of the Kynurenine Signaling Pathway in Different Chronic Pain Conditions and Potential Use of Therapeutic Agents. Int. J. Mol. Sci. 2020, 21, 6045. [Google Scholar] [CrossRef]
- Berger, A.M.; Mooney, K.; Alvarez-Perez, A.; Breitbart, W.S.; Carpenter, K.M.; Cella, D.; Cleeland, C.; Dotan, E.; Eisenberger, M.A.; Escalante, C.P.; et al. National comprehensive cancer network. Cancer-Related Fatigue, Version 2.2015. J. Natl. Compr. Canc. Netw. 2015, 13, 1012–1039. [Google Scholar] [CrossRef]
- Addington, A.M.; Gallo, J.J.; Ford, D.E.; Eaton, W.W. Epidemiology of unexplained fatigue and major depression in the community: The Baltimore ECA follow-up, 1981–1994. Psychol. Med. 2001, 31, 1037–1044. [Google Scholar] [CrossRef]
- Fava, M. Symptoms of fatigue and cognitive/executive dysfunction in major depressive disorder before and after antidepressant treatment. J. Clin. Psychiatry 2003, 64, 30–34. [Google Scholar]
- Endres, D.; Pollak, T.A.; Bechter, K.; Prüss, H.; Tebartz van Elst, L. Cerebrospinal Fluid Biomarkers for the Detection of Autoimmune Depression. Biol. Psychiatry 2022, 92, 530–532. [Google Scholar] [CrossRef]
- Höftberger, R.; Lassmann, H. Immune-mediated disorders. Handb. Clin. Neurol. 2018, 145, 285–299. [Google Scholar] [CrossRef]
- Grativvol, R.S.; Cavalcante, W.C.P.; Castro, L.H.M.; Nitrini, R.; Simabukuro, M.M. Updates in the Diagnosis and Treatment of Paraneoplastic Neurologic Syndromes. Curr. Oncol. Rep. 2018, 20, 92. [Google Scholar] [CrossRef] [PubMed]
- Iorio, R.; Spagni, G.; Masi, G. Paraneoplastic neurological syndromes. Semin. Diagn. Pathol. 2019, 36, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Dropcho, E.J. Neurologic paraneoplastic syndromes. Curr. Oncol. Rep. 2004, 6, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, E. Paraneoplastic disorders. Continuum (Minneap Minn). Neuro-Oncol. 2015, 21, 452–475. [Google Scholar] [CrossRef] [Green Version]
- Senties-Madrid, H.; Vega-Boada, F. Paraneoplastic syndromes associated with anti-Hu antibodies. Isr. Med. Assoc. J. 2001, 3, 94–103. [Google Scholar] [PubMed]
- Venkatraman, A.; Opal, P. Paraneoplastic cerebellar degeneration with anti-Yo antibodies—A review. Ann. Clin. Transl. Neurol. 2016, 3, 655–663. [Google Scholar] [CrossRef]
- Ortega Suero, G.; Sola-Valls, N.; Escudero, D.; Saiz, A.; Graus, F. Anti-Ma and anti-Ma2-associated paraneoplastic neurological syndromes. Neurologia 2018, 33, 18–27. [Google Scholar] [CrossRef]
- Olmez, O.F.; Kinikoglu, O.; Yilmaz, N.H.; Bilici, A.; Cubukcu, E.; Seker, M.; Cakir, T.; Yildiz, O.; Hamdard, J. Anti-Ri-associated paraneoplastic neurological syndrome: Initial symptom of breast cancer with HER2 overexpression and treatment by dual HER2 blockade. J. Oncol. Pharm. Pract. 2019, 25, 1526–1530. [Google Scholar] [CrossRef]
- Dalmau, J.; Tüzün, E.; Wu, H.Y.; Masjuan, J.; Rossi, J.E.; Voloschin, A.; Baehring, J.M.; Shimazaki, H.; Koide, R.; King, D.; et al. Paraneoplastic anti-N-methyl-D-aspartate receptor encephalitis associated with ovarian teratoma. Ann. Neurol. 2007, 61, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Laurido-Soto, O.; Brier, M.R.; Simon, L.E.; McCullough, A.; Bucelli, R.C.; Day, G.S. Patient characteristics and outcome associations in AMPA receptor encephalitis. J. Neurol. 2019, 266, 450–460. [Google Scholar] [CrossRef]
- Lancaster, E.; Lai, M.; Peng, X.; Hughes, E.; Constantinescu, R.; Raizer, J.; Friedman, D.; Skeen, M.B.; Grisold, W.; Kimura, A.; et al. Antibodies to the GABA(B) receptor in limbic encephalitis with seizures: Case series and characterisation of the antigen. Lancet Neurol. 2010, 9, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Lai, M.; Huijbers, M.G.; Lancaster, E.; Graus, F.; Bataller, L.; Balice-Gordon, R.; Cowel, J.K.; Dalmau, J. Investigation of LGI1 as the antigen in limbic encephalitis previously attributed to potassium channels: A case series. Lancet Neurol. 2010, 9, 776–785. [Google Scholar] [CrossRef] [Green Version]
- Bien, C.G.; Vincent, A.; Barnett, M.H.; Becker, A.J.; Blümcke, I.; Graus, F.; Jellinger, K.A.; Reuss, D.E.; Ribalta, T.; Schlegel, J.; et al. Immunopathology of autoantibody-associated encephalitides: Clues for pathogenesis. Brain 2012, 135, 1622–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Psimaras, D.; Carpentier, A.F.; Rossi, C. PNS Euronetwork. Cerebrospinal fluid study in paraneoplastic syndromes. J. Neurol. Neurosurg. Psychiatry 2010, 81, 42–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binks, S.; Uy, C.; Honnorat, J.; Irani, S.R. Paraneoplastic neurological syndromes: A practical approach to diagnosis and management. Pract. Neurol. 2022, 22, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Borovcanin, M.M.; Vesic, K. Breast cancer in schizophrenia could be interleukin-33-mediated. World J. Psychiatry 2021, 11, 1065–1074. [Google Scholar] [CrossRef] [PubMed]
- Vyas, D.; Laput, G.; Vyas, A.K. Chemotherapy-enhanced inflammation may lead to the failure of therapy and metastasis. OncoTargets Ther. 2014, 7, 1015–1023. [Google Scholar] [CrossRef] [Green Version]
- Proskuryakov, S.Y.; Gabai, V.L. Mechanisms of Tumor Cell Necrosis. Curr. Pharm. Des. 2010, 16, 56–68. [Google Scholar] [CrossRef]
- Jin, S.; DiPaola, R.S.; Mathew, R.; White, E. Metabolic catastrophe as a means to cancer cell death. J. Cell Sci. 2007, 120, 379–383. [Google Scholar] [CrossRef] [Green Version]
- Leek, R.D.; Landers, R.J.; Harris, A.L.; Lewis, C.E. Necrosis correlates with high vascular density and focal macrophage infltration in invasive carcinoma of the breast. Br. J. Cancer 1999, 79, 991–995. [Google Scholar] [CrossRef] [Green Version]
- Sauter, B.; Albert, M.L.; Francisco, L.; Larsson, M.; Somersan, S.; Bhardwaj, N. Consequences of cell death: Exposure to necrotic tumor cells, but not primary tissue cells or apoptotic cells, induces the maturation of immunostimulatory dendritic cells. J. Exp. Med. 2000, 191, 423–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francesco Colotta, F.; Allavena, P.; Sica, A.; Garlanda, C.; Mantovani, A. Cancer-related inflammation, the seventh hallmark of cancer: Links to genetic instability. Carcinogenesis 2009, 30, 1073–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolan, R.D.; McMillan, D.C. The prevalence of cancer associated systemic inflammation: Implications of prognostic studies using the Glasgow Prognostic Score. Crit. Rev. Oncol. Hematol. 2020, 150, 102962. [Google Scholar] [CrossRef] [PubMed]
- Ernst, B.; Anderson, K.S. Immunotherapy for the treatment of breast cancer. Curr. Oncol. Rep. 2015, 17, 5. [Google Scholar] [CrossRef] [PubMed]
- Dhupkar, P.; Gordon, N.; Stewart, J.; Kleinerman, E.S. Anti-PD-1 therapy redirects macrophages from an M2 to an M1 phenotype inducing regression of OS lung metastases. Cancer Med. 2018, 7, 2654–2664. [Google Scholar] [CrossRef]
- Parekh, V.V.; Lalani, S.; Kim, S.; Halder, R.; Azuma, M.; Yagita, H.; Kumar, V.; Wu, L.; Kaer, L.V. PD-1/PD-L blockade prevents anergy induction and enhances the anti-tumor activities of glycolipid-activated invariant NKT cells. J. Immunol. 2009, 182, 2816–2826. [Google Scholar] [CrossRef] [Green Version]
- Rotte, A.; D’Orazi, G.; Bhandaru, M. Nobel committee honors tumor immunologists. J. Exp. Clin. Cancer Res. 2018, 37, 262. [Google Scholar] [CrossRef] [Green Version]
- Helmy, K.Y.; Patel, S.A.; Nahas, G.R.; Rameshwar, P. Cancer immunotherapy: Accomplishments to date and future promise. Ther. Deliv. 2013, 4, 1307–1320. [Google Scholar] [CrossRef]
- Solomon, B.L.; Garrido-Laguna, I. TIGIT: A novel immunotherapy target moving from bench to bedside. Cancer Immunol. Immunother. 2018, 67, 1659–1667. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhong, H.; Fan, Y.; Liu, Q.; Song, J.; Yao, S.; Cao, F. Immune and Clinical Features of CD96 Expression in Glioma by in silico Analysis. Front. Bioeng. Biotechnol. 2020, 8, 592. [Google Scholar] [CrossRef]
- van Hall, T.; André, P.; Horowitz, A.; Ruan, D.F.; Borst, L.; Zerbib, R.; Narni-Mancinelli, E.; van der Burg, S.H.; Vivier, E. Monalizumab: Inhibiting the novel immune checkpoint NKG2A. J. Immunother. Cancer 2019, 7, 263. [Google Scholar] [CrossRef] [PubMed]
- Burugu, S.; Dancsok, A.R.; Nielsen, T.O. Emerging targets in cancer immunotherapy. Semin. Cancer Biol. 2018, 52, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Raison, C.L.; Rutherford, R.E.; Woolwine, B.J.; Shuo, C.; Schettler, P.; Drake, D.F.; Haroon, E.; Miller, A.H. A Randomized Controlled Trial of the Tumor Necrosis Factor Antagonist Infliximab for Treatment-Resistant Depression: The Role of Baseline Inflammatory Biomarkers. JAMA Psychiatry. 2013, 70, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Tookman, A.J.; Jones, C.L.; DeWitte, M.; Fatigu Lodge, P.J. Fatigue in patients with advanced cancer: A pilot study of an intervention with infliximab. Support Care Cancer 2008, 16, 1131–1140. [Google Scholar] [CrossRef]
- Loftus, E.V.; Feagan, B.G.; Colombel, J.F.; Rubin, D.T.; Wu, E.Q.; Yu, A.P.; Pollack, P.F.; Chao, J.; Mulani, P. Effects of adalimumab maintenance therapy on health-related quality of life of patients with Crohn’s disease: Patient-reported outcomes of the CHARM trial. Am. J. Gastroenterol. 2008, 103, 3132–3141. [Google Scholar] [CrossRef]
- Leman, J.; Walton, S.; Layton, A.M.; Ward, K.A.; McBride, S.; Cliff, S.; Downs, A.; Landeira, M.; Bewley, A. The real world impact of adalimumab on quality of life and the physical and psychological effects of moderate-to-severe psoriasis: A UK prospective, multicenter, observational study. J Dermatolog Treat. 2020, 31, 213–221. [Google Scholar] [CrossRef]
- Scheinfeld, N.; Sundaram, M.; Teixeira, H.; Gu, Y.; Okun, M. Reduction in pain scores and improvement in depressive symptoms in patients with hidradenitis suppurativa treated with adalimumab in a phase 2, randomized, placebo-controlled trial. Dermatol. Online J. 2016, 22, 13030/qt38x5922j. [Google Scholar] [CrossRef]
- Bayramgürler, D.; Karson, A.; Özer, C.; Utkan, T. Effects of long-term etanercept treatment on anxiety- and depression-like neurobehaviors in rats. Physiol Behav. 2013, 119, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Brymer, K.J.; Fenton, E.Y.; Kalynchuk, L.E.; Caruncho, H.J. Peripheral etanercept administration normalizes behavior, hippocampal neurogenesis, and hippocampal reelin and GABAA receptor expression in a preclinical model of depression. Front. Pharmacol. 2018, 9, 121. [Google Scholar] [CrossRef]
- Alshammari, M.A.; Khan, M.R.; Majid Mahmood, H.; Alshehri, A.O.; Alasmari, F.F.; Alqahtani, F.M.; Alasmari, A.F.; Alsharari, S.D.; Alhossan, A.; Ahmad, S.F. Systemic TNF-α blockade attenuates anxiety and depressive-like behaviors in db/db mice through downregulation of inflammatory signaling in peripheral immune cells. Saudi Pharm. J. 2020, 28, 621–629. [Google Scholar] [CrossRef]
- Tyring, S.; Gottlieb, A.; Papp, K.; Gordon, K.; Leonardi, C.; Wang, A.; Lalla, D.; Woolley, M.; Jahreis, A.; Zitnik, R.; et al. Etanercept and clinical outcomes, fatigue, and depression in psoriasis: Double-blind placebo-controlled randomised phase III trial. Lancet 2006, 367, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.C.; Gun, S.C.; Mok, C.C.; Khandker, R.; Nab, H.W.; Koenig, A.S.; Vlahos, B.; Pedersen, R.; Singh, A. Improved health outcomes with Etanercept versus usual DMARD therapy in an Asian population with established rheumatoid arthritis. BMC Musculoskelet. Disord. 2013, 14, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, B.; Aboul-Fotouh, S.; Ibrahim, E.A.; Shehata, H.; Mansour, A.A.; Az Yassin, N.; El-Eraky, W.; Abdel-Twab, A.M. Effects of pentoxifylline, 7-nitroindazole, and imipramine on tumor necrosis factor-α and indoleamine 2,3-dioxygenase enzyme activity in the hippocampus and frontal cortex of chronic mild-stress-exposed rats. Neuropsychiatr. Dis. Treat. 2013, 9, 697–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elgarf, A.S.A.; Aboul-Fotouh, S.; Abd-Alkhalek, H.A.; El Tabbal, M.; Hassan, A.N.; Kassim, S.K.; Hammouda, G.A.; Farrag, K.A.; Abdel-Tawab, A.M. Lipopolysaccharide repeated challenge followed by chronic mild stress protocol introduces a combined model of depression in rats: Reversibility by imipramine and pentoxifylline. Pharmacol. Biochem. Behav. 2014, 126, 152–162. [Google Scholar] [CrossRef]
- El-Haggar, S.M.; Eissa, M.A.; Mostafa, T.M.; El-Attar, K.S.; Abdallah, M.S. The phosphodiesterase inhibitor pentoxifylline as a novel adjunct to antidepressants in major depressive disorder patients: A proof-of-concept, randomized, double-blind, placebo-controlled trial. Psychother. Psychosom. 2018, 87, 331–339. [Google Scholar] [CrossRef]
- Langley, R.G.; Feldman, S.R.; Han, C.; Schenkel, B.; Szapary, P.; Hsu, M.C.; Ortonne, J.P.; Gordon, K.B.; Kimball, A.B. Ustekinumab significantly improves symptoms of anxiety, depression, and skin-related quality of life in patients with moderate-to-severe psoriasis: Results from a randomized, double-blind, placebo-controlled phase III trial. J. Am Acad Dermatol. 2010, 63, 457–465. [Google Scholar] [CrossRef]
- Cork, M.J.; Eckert, L.; Simpson, E.L.; Armstrong, A.; Barbarot, S.; Puig, L.; Girolomoni, G.; de Bruin-Weller, M.; Wollenberg, A.; Kataoka, Y.; et al. Dupilumab Improves Patient-Reported Symptoms of Atopic Dermatitis, Symptoms of Anxiety and Depression, and Health-Related Quality of Life in Moderate-to-Severe Atopic Dermatitis: Analysis of Pooled Data from the Randomized Trials SOLO 1 and SOLO 2. J. Dermatolog. Treat. 2020, 31, 606–614. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, C.E.M.; Fava, M.; Miller, A.H.; Russell, J.; Ball, S.G.; Xu, W.; Acharya, N.; Rapaport, M.H. Impact of Ixekizumab Treatment on Depressive Symptoms and Systemic Inflammation in Patients with Moderate-to-Severe Psoriasis: An Integrated Analysis of Three Phase 3 Clinical Studies. Psychother. Psychosom. 2017, 86, 260–267. [Google Scholar] [CrossRef]
- Gordon, K.B.; Armstrong, A.W.; Han, C.; Foley, P.; Song, M.; Wasfi, Y.; You, Y.; Shen, Y.K.; Reich, K. Anxiety and Depression in Patients with Moderate-to-Severe Psoriasis and Comparison of Change from Baseline after Treatment with Guselkumab vs. Adalimumab: Results from the Phase 3 VOYAGE 2 Study. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 1940–1949. [Google Scholar] [CrossRef]
- Leonard, B.E. Inflammation and depression: A causal or coincidental link to the pathophysiology? Acta Neuropsychiatr. 2018, 30, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Dionisie, V.; Filip, G.A.; Manea, M.C.; Manea, M.; Riga, S. The anti-inflammatory role of SSRI and SNRI in the treatment of depression: A review of human and rodent research studies. Inflammopharmacology 2021, 29, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.J.; Wei, Y.B.; Strawbridge, R.; Bao, Y.; Chang, S.; Shi, L.; Que, J.; Gadad, B.S.; Trivedi, M.H.; Kelsoe, J.R.; et al. Peripheral cytokine levels and response to antidepressant treatment in depression: A systematic review and meta-analysis. Mol. Psychiatry 2020, 25, 339–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levite, M. Neurotransmitters activate T-cells and elicit crucial functions via neurotransmitter receptors. Curr. Opin. Pharmacol. 2008, 8, 460–471. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, S.H.; Hosseini, F.; Modabbernia, A.; Ashrafi, M.; Akhondzadeh, S. Effect of celecoxib add-on treatment on symptoms and serum IL-6 concentrations in patients with major depressive disorder: Randomized double-blind placebo-controlled study. J. Affect Disord. 2012, 141, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.S.; Kaneko, N.; Sawamoto, K. Minocycline treatment ameliorates interferon-alpha- induced neurogenic defects and depression-like behaviors in mice. Front. Cell Neurosci. 2015, 9, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohler, O.; Krogh, J.; Mors, O.; Benros, M.E. Inflammation in Depression and the Potential for Anti-Inflammatory Treatment. Curr. Neuropharmacol 2016, 14, 732–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, M.; Shi, Z.; Sha, N.N.; Chen, N.; Peng, S.Y.; Liao, D.F.; Wong, M.S.; Dong, X.L.; Wang, Y.J.; Yuan, T.F.; et al. Paricalcitol alleviates lipopolysaccharide-induced depressive-like behavior by suppressing hypothalamic microglia activation and neuroinflammation. Biochem. Pharmacol. 2019, 163, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Elewa, H.F.; Hilali, H.; Hess, D.C.; Machado, L.S.; Fagan, S.C. Minocycline for short-term neuroprotection. Pharmacotherapy 2006, 26, 515–521. [Google Scholar] [CrossRef] [Green Version]
- Avan, R.; Sahebnasagh, A.; Hashemi, J.; Monajati, M.; Faramarzi, F.; Henney, N.C.; Montecucco, F.; Jamialahmadi, T.; Sahebkar, A. Update on Statin Treatment in Patients with Neuropsychiatric Disorders. Life 2021, 11, 1365. [Google Scholar] [CrossRef]
- Zhao, Q.; Wu, X.; Yan, S.; Xie, X.; Fan, Y.; Zhang, J.; Peng, C.; You, Z. The antidepressant-like effects of pioglitazone in a chronic mild stress mouse model are associated with PPARγ-mediated alteration of microglial activation phenotypes. J. Neuroinflammation. 2016, 13, 259. [Google Scholar] [CrossRef] [Green Version]
- Kemp, D.E.; Schinagle, M.; Gao, K.; Conroy, C.; Ganocy, S.J.; Ismail-Beigi, F.; Calabrese, J.R. PPAR-gamma agonism as a modulator of mood: Proof-of-concept for pioglitazone in bipolar depression. CNS Drugs. 2014, 28, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Zager, A. Modulating the immune response with the wake-promoting drug modafinil: A potential therapeutic approach for inflammatory disorders. Brain Behav. Immunity. 2020, 88, 878–886. [Google Scholar] [CrossRef]
- DeBattista, C.; Posener, J.A.; Kalehzan, B.M.; Schatzberg, A.F. Acute antidepressant effects of intravenous hydrocortisone and CRH in depressed patients: A double-blind, placebo-controlled study. Am. J. Psychiatry. 2000, 157, 1334–1337. [Google Scholar] [CrossRef]
- Rice, J.B.; White, A.G.; Scarpati, L.M.; Wan, G.; Nelson, W.W. Long-term Systemic Corticosteroid Exposure: A Systematic Literature Review. Clin. Ther. 2017, 39, 2216–2229. [Google Scholar] [CrossRef] [Green Version]
- Sforzini, L.; Nettis, M.A.; Mondelli, V.; Pariante, C.M. Inflammation in cancer and depression: A starring role for the kynurenine pathway. Psychopharmacology 2019, 236, 2997–3011. [Google Scholar] [CrossRef] [Green Version]
- Sohl, S.J.; Weaver, K.E.; Birdee, G.; Kent, E.E.; Danhauer, S.C.; Hamilton, A.S. Characteristics Associated with the Use of Complementary Health Approaches among Long-Term Cancer Survivors. Support. Care Cancer 2014, 22, 927–936. [Google Scholar] [CrossRef] [Green Version]
- Gansler, T.; Strollo, S.; Fallon, E.; Leach, C. Use of Complementary/Integrative Methods: Cancer Survivors’ Misconceptions about Recurrence Prevention. J. Cancer Surviv. 2019, 13, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Werneke, U. Risk Management of Nutritional Supplements in Chronic Illness: The Implications for the Care of Cancer and Depression. Proc. Nutr. Soc. 2007, 66, 483–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drake, E.N. Cancer Chemoprevention: Selenium as a Prooxidant, Not an Antioxidant. Med. Hypotheses 2006, 67, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Castaño, A. Low Selenium Diet Increases the Dopamine Turnover in Prefrontal Cortex of the Rat. Neurochem. Int. 1997, 30, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Young, S.N. Folate and Depression—A Neglected Problem. J. Psychiatry Neurosci. 2007, 32, 80–82. [Google Scholar] [PubMed]
- Grosso, G.; Galvano, F.; Marventano, S.; Malaguarnera, M.; Bucolo, C.; Drago, F.; Caraci, F. Omega-3 Fatty Acids and Depression: Scientific Evidence and Biological Mechanisms. Oxid. Med. Cell Longev. 2014, 2014, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, M.P.; Hibbeln, J.R.; Wisner, K.L.; Davis, J.M.; Mischoulon, D.; Peet, M.; Keck, P.E.; Marangell, L.B.; Richardson, A.J.; Lake, J. Omega-3 Fatty Acids: Evidence Basis for Treatment and Future Research in Psychiatry. J. Clin. Psychiatry 2006, 67, 1954–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardman, W.E. (N-3) Fatty Acids and Cancer Therapy. J. Nutr. 2004, 134, S3427–S3430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jevremović, A.; Božinović, N.; Arsenijević, D.; Marmakov, S.; Nedić Vasiljević, B.; Uskoković-Marković, S.; Bajuk-Bogdanović, D.; Milojević-Rakić, M. Modulation of cytotoxicity by consecutive adsorption of tannic acid and pesticides on surfactant functionalized zeolites, Environ Sci Process. Impacts 2020, 22, 2199–2211. [Google Scholar] [CrossRef]
- Janićijević, A.; Jevremović, A.; Janošević Ležaić, B.; Nedić Vasiljević, S.; Uskoković-Marković, D.; Bajuk-Bogdanović, M.; Milojević-Rakić, M. Comparative assessment of pesticide adsorption capacity and antioxidant activity of Silver Dodecatungstophosphate/HΒEA zeolite composites. J. Environ. Chem. Eng. 2021, 9, 106341. [Google Scholar] [CrossRef]
- Pavelić, S.K.; Medica, J.S.; Gumbarević, D.; Filošević, A.; Pržulj, N.; Pavelić, K. Critical review on zeolite clinoptilolite safety and medical applications in vivo. Front. Pharmacol. 2018, 9, 1350. [Google Scholar] [CrossRef]
- Mijailović, N.R.; Nedić Vasiljević, B.; Ranković, M.; Milanović, V.; Uskoković-Marković, S. Environmental and Pharmacokinetic Aspects of Zeolite/Pharmaceuticals Systems—Two Facets of Adsorption Ability. Catalysts 2022, 12, 837. [Google Scholar] [CrossRef]
- Krajišnik, D.; Daković, A.; Milojević, M.; Malenović, A.; Kragović, M.; Bajuk-Bogdanović, D.; Dondur, V.; Milić, J. Properties of diclofenac sodium sorption onto natural zeolite modified with cetylpyridinium chloride. Colloids Surf. B Biointerfaces 2011, 83, 165–172. [Google Scholar] [CrossRef]
- Krajišnik, D.; Daković, A.; Malenović, M.; Milojević-Rakić, M.; Dondur, V.; Radulović, Ž.; Milić, J.; Radulović, Z.; Milić, J. Investigation of adsorption and release of diclofenac sodium by modified zeolites composites. Appl. Clay Sci. 2013, 83–84, 322–326. [Google Scholar] [CrossRef]
- Hassanvand, A.; Gharibzadeh, S. Is It Possible to Treat Some Brain Diseases by Drug-Substituted Zeolites? J. Neuropsychiatry Clin. Neurosci 2013, 25, E04. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Hong, Z.; Xue, J.; Luo, J.; Liu, Q.; Chen, X.; Pan, Z.; Zhou, J.; Liu, Z.; Chen, T. Near-Infrared Radiation-Assisted Drug Delivery Nanoplatform to Realize Blood–Brain Barrier Crossing and Protection for Parkinsonian Therapy. ACS Appl. Mater. Interfaces 2021, 13, 37746–37760. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Niu, M.; Chen, X.; Tan, L.; Fu, C.; Ren, X.; Ren, J.; Li, L.; Xu, K.; Zhong, H.; et al. Biocompatible and biodegradable zeolitic imidazolate framework/polydopamine nanocarriers for dual stimulus triggered tumor thermo-chemotherapy. Biomaterials 2018, 162, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Alchujyan, N.K.; Hovhannisyan, M.R.; Movsesyan, N.H.; Madoyan, R.A.; Sargsyan, H.H.; Aghababova, A.A.; Minasyan, G.H.; Hairapetyan, H.L.; Kevorkian, R.G.; Chailyan, S.G.; et al. Free and zeolite-immobilized mixture versus sodium valproate in prevention of oxidative stress and modulation of the larginine intracellular metabolic pathways in the rat brain and blood following dexamphetamine-induced bipolar disorder. Eur. Chem. Bull 2018, 7, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Eisenwagen, S.; Pavelic, K. Potential Role of Zeolites in Rehabilitation of Cancer Patients. Arch. Physiother. Rehabil 2022, 3, 29–40. Available online: http://www.fotunejournals.com/potential-role-of-zeolites-in-rehabilitation-of-cancer-patients.html (accessed on 21 August 2022).
- Johnson, H.H.; Microchemical, A. Method for the Determination of Histamine. AMA Arch. Derm. 1955, 72, 307. [Google Scholar] [CrossRef]
- Mück-Šeler, D.; Pivac, N. The effect of natural clinoptilolite on the serotonergic receptors in the brain of mice with mammary carcinoma. Life Sci. 2003, 73, 2059–2069. [Google Scholar] [CrossRef]
- Vitale, M.G.; Barbato, C.; Crispo, A.; Habetswallner, F.; De Martino, B.M.; Riccardi, F.; Maione, A.; Eisenwagen, S.; Vitale, G.; Cartenì, G.; et al. Multi Trial: A Randomized, Double-Blinded, Placebo-Controlled Trial of Oral PMA-Zeolite to Prevent Chemotherapy-Induced Side Effects, in Particular, Peripheral Neuropathy. Molecules 2020, 25, 2297. [Google Scholar] [CrossRef]
- Basha, M.P.; Begum, S.; Mir, B.A. Neuroprotective Actions of Clinoptilolite and Ethylenediaminetetraacetic Acid Against Lead-induced Toxicity in Mice Mus musculus. Toxicol. Int. 2013, 20, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Delavarian, M.; Hassanvand, A.; Gharibzadeh, S. Increasing performance in children with ADHD by trapping lead with a nano-zeolite. J. Neuropsychiatry Clin. Neurosci. 2013, 25, E23. [Google Scholar] [CrossRef]
- Teixeira, M.F.S.; Bergamini, M.F.; Marques, C.M.P.; Bocchi, N. Voltammetric determination of L-dopa using an electrode modified with trinuclear ruthenium ammine complex (Ru-red) supported on Y-type zeolite. Talanta 2004, 63, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Delavarian, M.; Hassanvand, A.; Gharibzadeh, S. Using natural zeolite as a transporter of dopamine. J. Neuropsychiatry Clin. Neurosci. 2013, 25, E21. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Drug | Class | Reference |
|---|---|---|
| Celecoxib | NSAIDs | [137] |
| Minocycline | tetracycline antibiotics | [138] |
| Statins | HMG-CoA reductase inhibitor | [139] |
| Pioglitazone | Antihyperglycemic | [139] |
| Modafinil | wakefulness promoting agents | [139] |
| Steroids | corticosteroids | [139] |
| Paricalcitol | vitamin D analog | [140] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borovcanin, M.M.; Vesić, K.; Arsenijević, D.; Milojević-Rakić, M.; Mijailović, N.R.; Jovanovic, I.P. Targeting Underlying Inflammation in Carcinoma Is Essential for the Resolution of Depressiveness. Cells 2023, 12, 710. https://doi.org/10.3390/cells12050710
Borovcanin MM, Vesić K, Arsenijević D, Milojević-Rakić M, Mijailović NR, Jovanovic IP. Targeting Underlying Inflammation in Carcinoma Is Essential for the Resolution of Depressiveness. Cells. 2023; 12(5):710. https://doi.org/10.3390/cells12050710
Chicago/Turabian StyleBorovcanin, Milica M., Katarina Vesić, Dragana Arsenijević, Maja Milojević-Rakić, Nataša R. Mijailović, and Ivan P. Jovanovic. 2023. "Targeting Underlying Inflammation in Carcinoma Is Essential for the Resolution of Depressiveness" Cells 12, no. 5: 710. https://doi.org/10.3390/cells12050710