

Inflammatory Cell Dynamics after Murine Femoral Artery Wire Injury: A Multi-Parameter Flow Cytometry-Based Analysis
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Unilateral Femoral Artery Wire Injury
2.2. Immunofluorescence and Quantification of Staining
2.3. Preparation of Single-Cell Suspensions
2.4. Immunostaining and Flow Cytometry
2.5. Statistical Analysis
3. Results
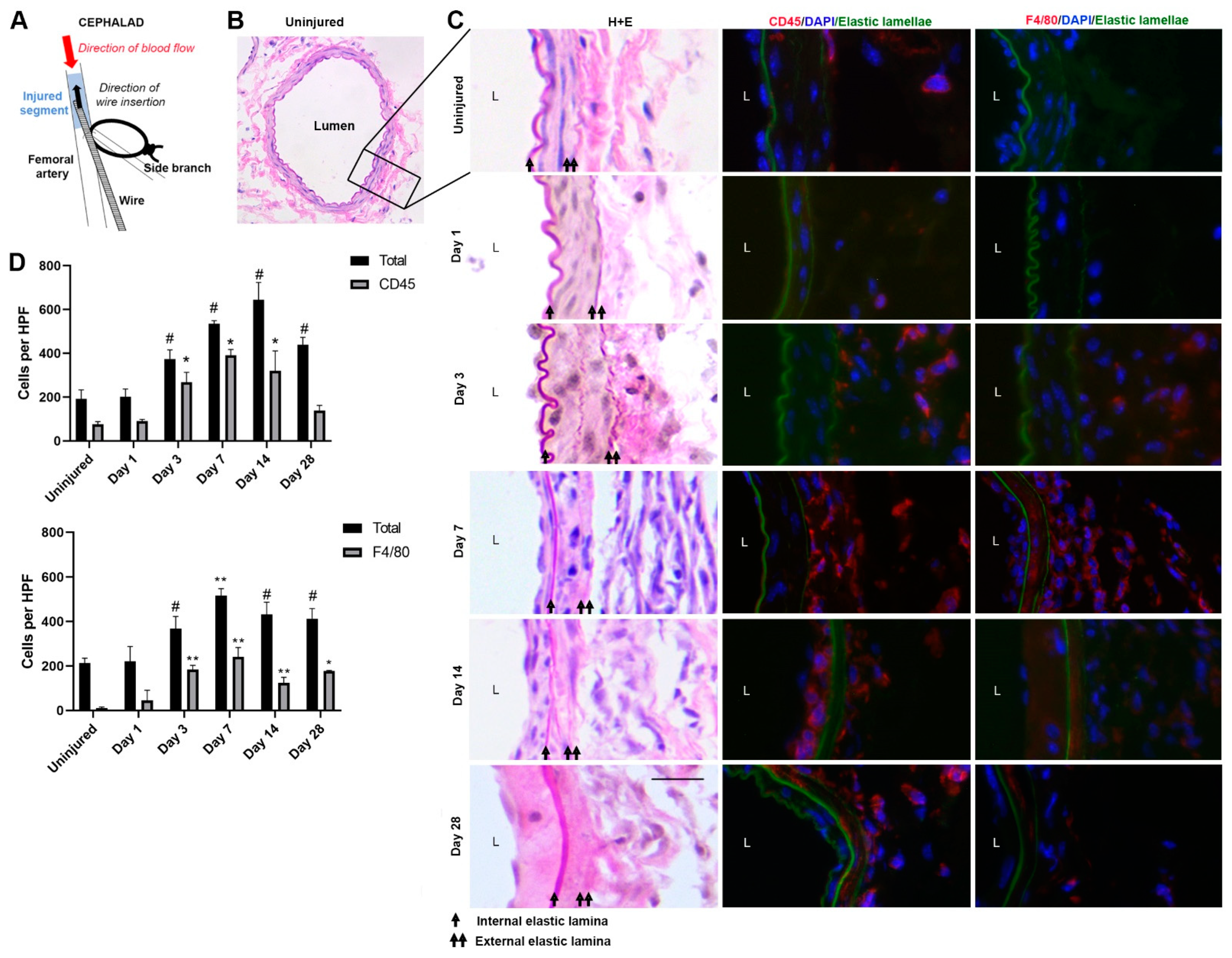
3.1. Immunofluorescence Staining of Femoral Artery Sections
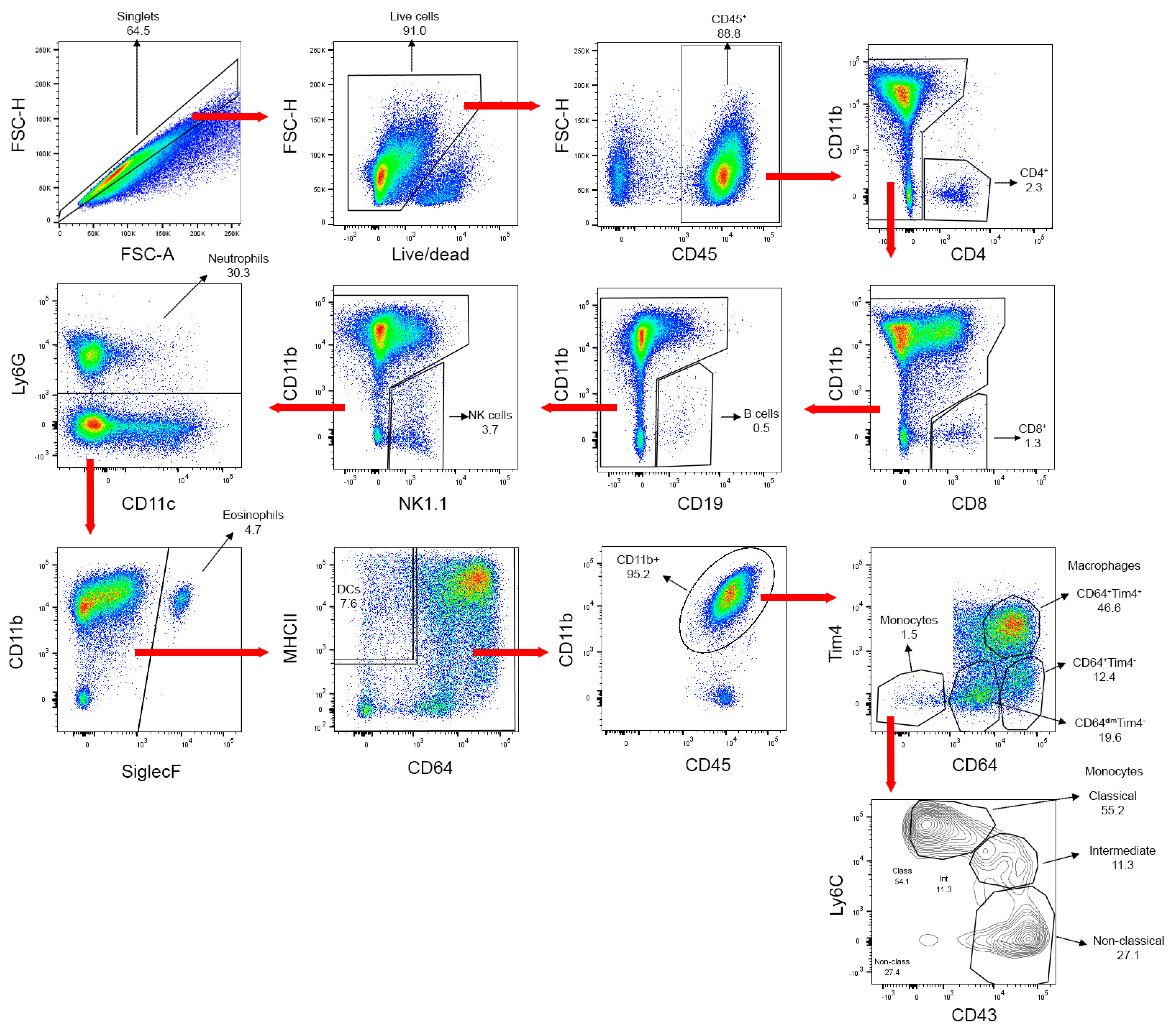
3.2. Flow Cytometry Gating Strategy for Leukocyte Sub-Populations
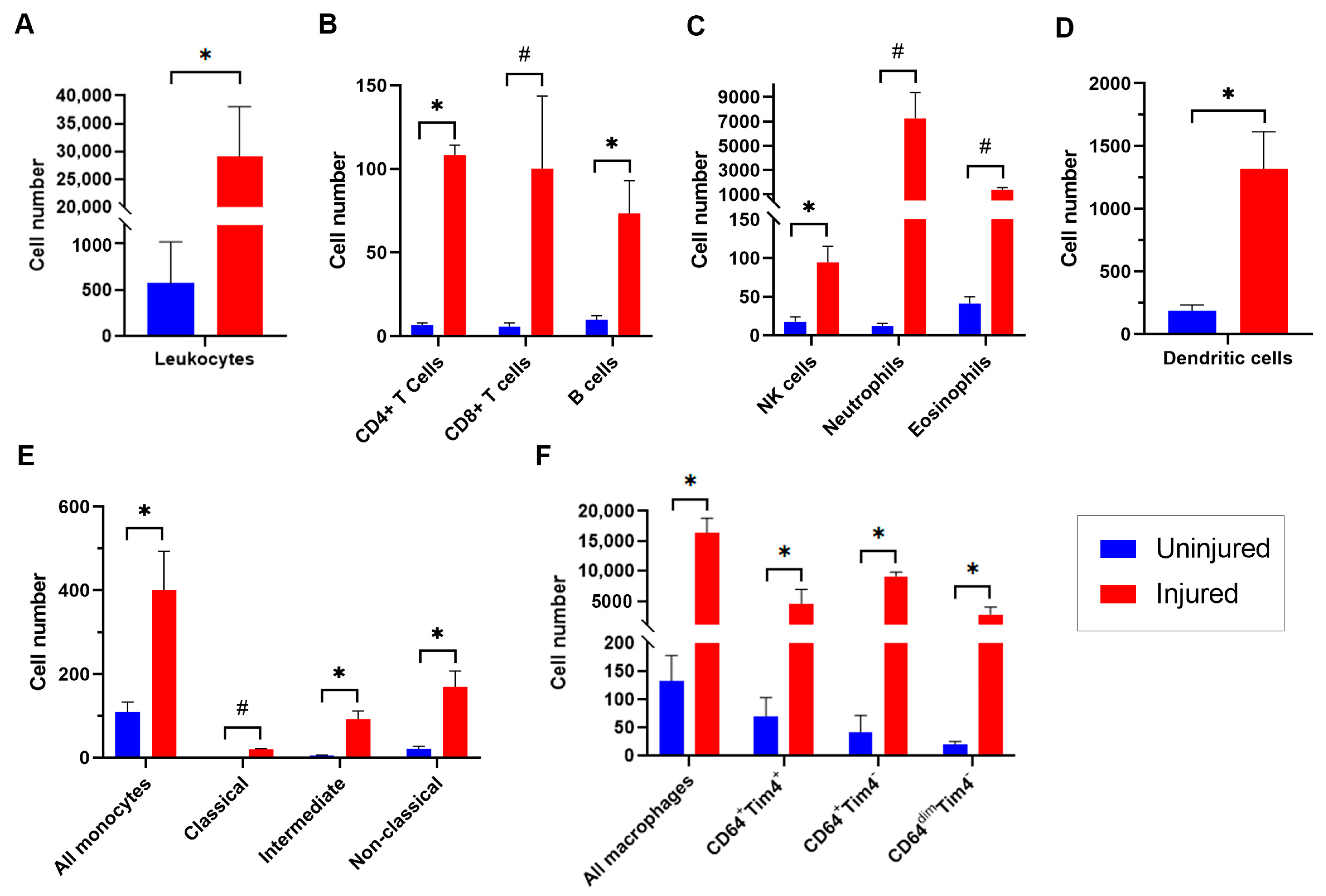
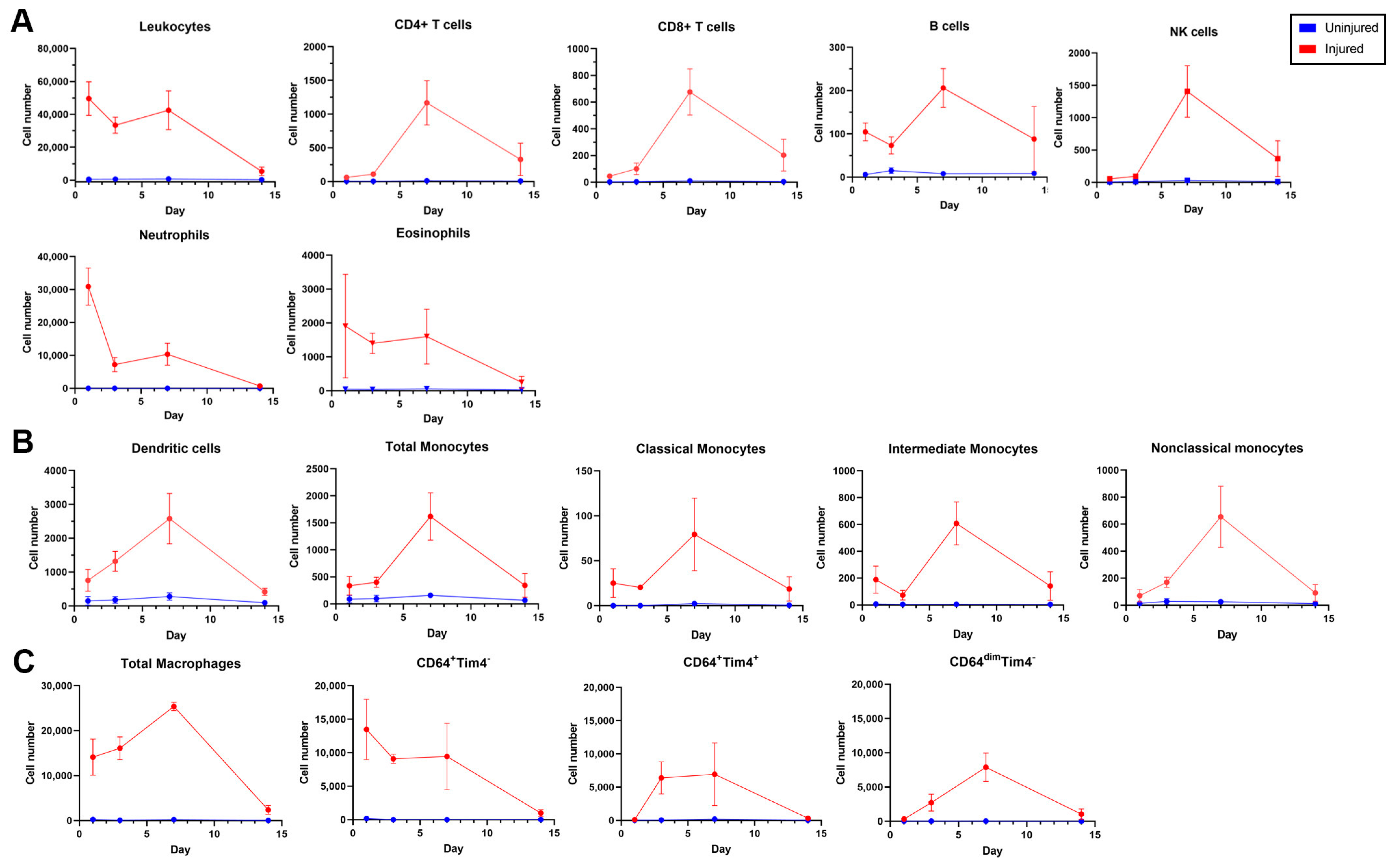
3.3. Dynamics of Leukocyte Accumulation in Injured Femoral Arteries
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kapadia, M.R.; Chow, L.W.; Tsihlis, N.D.; Ahanchi, S.S.; Eng, J.W.; Murar, J.; Martinez, J.; Popowich, D.A.; Jiang, Q.; Hrabie, J.A.; et al. Nitric oxide and nanotechnology: A novel approach to inhibit neointimal hyperplasia. J. Vasc. Surg. 2008, 47, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Ramos, J.R.; Berger, K.; Mansfield, P.B.; Sauvage, L.R. Histologic fate and endothelial changes of distended and nondistended vein grafts. Ann. Surg. 1976, 183, 205–228. [Google Scholar] [CrossRef]
- Owens, C.D.; Ridker, P.M.; Belkin, M.; Hamdan, A.D.; Pomposelli, F.; Logerfo, F.; Creager, M.A.; Conte, M.S. Elevated C-reactive protein levels are associated with postoperative events in patients undergoing lower extremity vein bypass surgery. J. Vasc. Surg. 2007, 45, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Stark, V.K.; Hoch, J.R.; Warner, T.F.; Hullett, D.A. Monocyte chemotactic protein-1 expression is associated with the development of vein graft intimal hyperplasia. Arter. Thromb. Vasc. Biol. 1997, 17, 1614–1621. [Google Scholar] [CrossRef]
- Huang, B.; Dreyer, T.; Heidt, M.; Yu, J.C.; Philipp, M.; Hehrlein, F.W.; Katz, N.; Al-Fakhri, N. Insulin and local growth factor PDGF induce intimal hyperplasia in bypass graft culture models of saphenous vein and internal mammary artery. Eur. J. Cardiothorac. Surg. 2002, 21, 1002–1008. [Google Scholar] [CrossRef] [Green Version]
- Guerri-Guttenberg, R.A.; Castilla, R.; Francos, G.C.; Muller, A.; Ambrosio, G.; Milei, J. Transforming growth factor beta1 and coronary intimal hyperplasia in pediatric patients with congenital heart disease. Can. J. Cardiol. 2013, 29, 849–857. [Google Scholar] [CrossRef]
- Wang, C.H.; Li, S.H.; Weisel, R.D.; Fedak, P.W.; Hung, A.; Li, R.K.; Rao, V.; Hyland, K.; Cherng, W.J.; Errett, L.; et al. Tetrahydrobiopterin deficiency exaggerates intimal hyperplasia after vascular injury. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R299–R304. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Gong, Y.; Zhang, R.; Piao, L.; Li, X.; Liu, Q.; Yan, S.; Shen, Y.; Guo, S.; Zhu, M.; et al. Resolvin E1 attenuates injury-induced vascular neointimal formation by inhibition of inflammatory responses and vascular smooth muscle cell migration. FASEB J. 2018, 32, 5413–5425. [Google Scholar] [CrossRef]
- Jukema, J.W.; Verschuren, J.J.; Ahmed, T.A.; Quax, P.H. Restenosis after PCI. Part 1: Pathophysiology and risk factors. Nat. Rev. Cardiol. 2011, 9, 53–62. [Google Scholar] [CrossRef]
- Grewe, P.H.; Deneke, T.; Machraoui, A.; Barmeyer, J.; Muller, K.M. Acute and chronic tissue response to coronary stent implantation: Pathologic findings in human specimen. J. Am. Coll. Cardiol. 2000, 35, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Burke, A.P.; Kolodgie, F.D.; Farb, A.; Weber, D.; Virmani, R. Morphological predictors of arterial remodeling in coronary atherosclerosis. Circulation 2002, 105, 297–303. [Google Scholar] [CrossRef]
- Farb, A.; Weber, D.K.; Kolodgie, F.D.; Burke, A.P.; Virmani, R. Morphological predictors of restenosis after coronary stenting in humans. Circulation 2002, 105, 2974–2980. [Google Scholar] [CrossRef] [Green Version]
- Owens, C.D.; Rybicki, F.J.; Wake, N.; Schanzer, A.; Mitsouras, D.; Gerhard-Herman, M.D.; Conte, M.S. Early remodeling of lower extremity vein grafts: Inflammation influences biomechanical adaptation. J. Vasc. Surg. 2008, 47, 1235–1242. [Google Scholar] [CrossRef] [Green Version]
- Miyahara, T.; Runge, S.; Chatterjee, A.; Chen, M.; Mottola, G.; Fitzgerald, J.M.; Serhan, C.N.; Conte, M.S. D-series resolvin attenuates vascular smooth muscle cell activation and neointimal hyperplasia following vascular injury. FASEB J. 2013, 27, 2220–2232. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Mottola, G.; Chatterjee, A.; Lance, K.D.; Chen, M.; Siguenza, I.O.; Desai, T.A.; Conte, M.S. Perivascular delivery of resolvin D1 inhibits neointimal hyperplasia in a rat model of arterial injury. J. Vasc. Surg. 2017, 65, 207–217.e3. [Google Scholar] [CrossRef] [Green Version]
- Akagi, D.; Chen, M.; Toy, R.; Chatterjee, A.; Conte, M.S. Systemic delivery of proresolving lipid mediators resolvin D2 and maresin 1 attenuates intimal hyperplasia in mice. FASEB J. 2015, 29, 2504–2513. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Menocal, L.; Faridi, M.H.; Martinez, L.; Shehadeh, L.A.; Duque, J.C.; Wei, Y.; Mesa, A.; Pena, A.; Gupta, V.; Pham, S.M.; et al. Macrophage-derived IL-18 and increased fibrinogen deposition are age-related inflammatory signatures of vascular remodeling. Am. J. Physiol. Heart Circ. Physiol. 2014, 306, H641–H653. [Google Scholar] [CrossRef]
- Vavra, A.K.; Havelka, G.E.; Martinez, J.; Lee, V.R.; Fu, B.; Jiang, Q.; Keefer, L.K.; Kibbe, M.R. Insights into the effect of nitric oxide and its metabolites nitrite and nitrate at inhibiting neointimal hyperplasia. Nitric. Oxide. 2011, 25, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Grassia, G.; Maddaluno, M.; Guglielmotti, A.; Mangano, G.; Biondi, G.; Maffia, P.; Ialenti, A. The anti-inflammatory agent bindarit inhibits neointima formation in both rats and hyperlipidaemic mice. Cardiovasc. Res. 2009, 84, 485–493. [Google Scholar] [CrossRef]
- Xing, D.; Feng, W.; Not, L.G.; Miller, A.P.; Zhang, Y.; Chen, Y.F.; Majid-Hassan, E.; Chatham, J.C.; Oparil, S. Increased protein O-GlcNAc modification inhibits inflammatory and neointimal responses to acute endoluminal arterial injury. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H335–H342. [Google Scholar] [CrossRef] [Green Version]
- Religa, P.; Bojakowski, K.; Bojakowska, M.; Gaciong, Z.; Thyberg, J.; Hedin, U. Allogenic immune response promotes the accumulation of host-derived smooth muscle cells in transplant arteriosclerosis. Cardiovasc. Res. 2005, 65, 535–545. [Google Scholar] [CrossRef]
- Hay, C.; Micko, C.; Prescott, M.F.; Liau, G.; Robinson, K.; De Leon, H. Differential cell cycle progression patterns of infiltrating leukocytes and resident cells after balloon injury of the rat carotid artery. Arter. Thromb. Vasc. Biol. 2001, 21, 1948–1954. [Google Scholar] [CrossRef] [Green Version]
- Simon, D.I.; Dhen, Z.; Seifert, P.; Edelman, E.R.; Ballantyne, C.M.; Rogers, C. Decreased neointimal formation in Mac-1(-/-) mice reveals a role for inflammation in vascular repair after angioplasty. J. Clin. Investig. 2000, 105, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Bahnson, E.S.; Vavra, A.K.; Flynn, M.E.; Vercammen, J.M.; Jiang, Q.; Schwartz, A.R.; Kibbe, M.R. Long-term effect of PROLI/NO on cellular proliferation and phenotype after arterial injury. Free Radic. Biol. Med. 2016, 90, 272–286. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Hoover, J.L.; Simmons, C.A.; Lindner, V.; Shebuski, R.J. Remodeling and neointimal formation in the carotid artery of normal and P-selectin-deficient mice. Circulation 1997, 96, 4333–4342. [Google Scholar] [CrossRef]
- Sata, M.; Maejima, Y.; Adachi, F.; Fukino, K.; Saiura, A.; Sugiura, S.; Aoyagi, T.; Imai, Y.; Kurihara, H.; Kimura, K.; et al. A mouse model of vascular injury that induces rapid onset of medial cell apoptosis followed by reproducible neointimal hyperplasia. J. Mol. Cell Cardiol. 2000, 32, 2097–2104. [Google Scholar] [CrossRef]
- Lindner, V.; Fingerle, J.; Reidy, M.A. Mouse model of arterial injury. Circ. Res. 1993, 73, 792–796. [Google Scholar] [CrossRef] [Green Version]
- Moroi, M.; Zhang, L.; Yasuda, T.; Virmani, R.; Gold, H.K.; Fishman, M.C.; Huang, P.L. Interaction of genetic deficiency of endothelial nitric oxide, gender, and pregnancy in vascular response to injury in mice. J. Clin. Investig. 1998, 101, 1225–1232. [Google Scholar] [CrossRef] [Green Version]
- Sata, M.; Tanaka, K.; Fukuda, D. Wire-Mediated Endovascular Injury that Induces Rapid Onset of Medial Cell Apoptosis Followed by Reproducible Neointimal Hyperplasia. In Mouse Models of Vascular Diseases; Sata, M., Ed.; Springer: Tokyo, Japan, 2016; pp. 3–20. [Google Scholar]
- McKinnon, K.M. Multiparameter Conventional Flow Cytometry. Methods Mol. Biol. 2018, 1678, 139–150. [Google Scholar]
- Nooromid, M.; Chen, E.B.; Xiong, L.; Shapiro, K.; Jiang, Q.; Demsas, F.; Eskandari, M.; Priyadarshini, M.; Chang, E.B.; Layden, B.T.; et al. Microbe-Derived Butyrate and Its Receptor, Free Fatty Acid Receptor 3, but Not Free Fatty Acid Receptor 2, Mitigate Neointimal Hyperplasia Susceptibility After Arterial Injury. J. Am. Heart Assoc. 2020, 9, e05247. [Google Scholar] [CrossRef]
- Wun, K.; Theriault, B.R.; Pierre, J.F.; Chen, E.B.; Leone, V.A.; Harris, K.G.; Xiong, L.; Jiang, Q.; Spedale, M.; Eskandari, O.M.; et al. Microbiota control acute arterial inflammation and neointimal hyperplasia development after arterial injury. PLoS ONE 2018, 13, e0208426. [Google Scholar] [CrossRef]
- Takayama, T.; Shi, X.; Wang, B.; Franco, S.; Zhou, Y.; DiRenzo, D.; Kent, A.; Hartig, P.; Zent, J.; Guo, L.W. A murine model of arterial restenosis: Technical aspects of femoral wire injury. J. Vis. Exp. 2015, 97, e52561. [Google Scholar]
- Roque, M.; Fallon, J.T.; Badimon, J.J.; Zhang, W.X.; Taubman, M.B.; Reis, E.D. Mouse model of femoral artery denudation injury associated with the rapid accumulation of adhesion molecules on the luminal surface and recruitment of neutrophils. Arter. Thromb. Vasc. Biol. 2000, 20, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Hagita, S.; Haraguchi, M.; Kajimura, M.; Suematsu, M.; Yoshida, M. Real-time imaging of mechanically injured femoral artery in mice reveals a biphasic pattern of leukocyte accumulation. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H1876–H1882. [Google Scholar]
- Zhang, Q.; Wang, H.; Wu, X.; Liu, B.; Liu, W.; Wang, R.; Liang, X.; Ma, C.; Gao, L. TIM-4 promotes the growth of non-small-cell lung cancer in a RGD motif-dependent manner. Br. J. Cancer 2015, 113, 1484–1492. [Google Scholar] [CrossRef] [Green Version]
- Meyers, J.H.; Chakravarti, S.; Schlesinger, D.; Illes, Z.; Waldner, H.; Umetsu, S.E.; Kenny, J.; Zheng, X.X.; Umetsu, D.T.; DeKruyff, R.H.; et al. TIM-4 is the ligand for TIM-1, and the TIM-1-TIM-4 interaction regulates T cell proliferation. Nat. Immunol. 2005, 6, 455–464. [Google Scholar] [CrossRef]
- Xu, L.; Qi, J.; Zhao, P.; Liang, X.; Ju, Y.; Liu, P.; Liu, B.; Guo, C.; Zhang, L.; Ma, C.; et al. T cell immunoglobulin- and mucin-domain-containing molecule-4 attenuates concanavalin A-induced hepatitis by regulating macrophage. J. Leukoc. Biol. 2010, 88, 329–336. [Google Scholar] [CrossRef]
- Xu, L.Y.; Qi, J.N.; Liu, X.; Ma, H.X.; Yuan, W.; Zhao, P.Q.; Liang, X.H.; Xu, Y.; Wang, H.X.; Xu, X.Y.; et al. Tim-4 Inhibits NO Generation by Murine Macrophages. PLoS ONE 2015, 10, e0124771. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, N.; Karisola, P.; Pena-Cruz, V.; Dorfman, D.M.; Jinushi, M.; Umetsu, S.E.; Butte, M.J.; Nagumo, H.; Chernova, I.; Zhu, B.; et al. TIM-1 and TIM-4 glycoproteins bind phosphatidylserine and mediate uptake of apoptotic cells. Immunity 2007, 27, 927–940. [Google Scholar] [CrossRef] [Green Version]
- Miyanishi, M.; Tada, K.; Koike, M.; Uchiyama, Y.; Kitamura, T.; Nagata, S. Identification of Tim4 as a phosphatidylserine receptor. Nature 2007, 450, 435–439. [Google Scholar] [CrossRef] [Green Version]
- Foks, A.C.; Engelbertsen, D.; Kuperwaser, F.; Alberts-Grill, N.; Gonen, A.; Witztum, J.L.; Lederer, J.; Jarolim, P.; DeKruyff, R.H.; Freeman, G.J.; et al. Blockade of Tim-1 and Tim-4 Enhances Atherosclerosis in Low-Density Lipoprotein Receptor-Deficient Mice. Arter. Thromb. Vasc. Biol. 2016, 36, 456–465. [Google Scholar] [CrossRef] [Green Version]
- Phillips, J.W.; Barringhaus, K.G.; Sanders, J.M.; Hesselbacher, S.E.; Czarnik, A.C.; Manka, D.; Vestweber, D.; Ley, K.; Sarembock, I.J. Single injection of P-selectin or P-selectin glycoprotein ligand-1 monoclonal antibody blocks neointima formation after arterial injury in apolipoprotein E-deficient mice. Circulation 2003, 107, 2244–2249. [Google Scholar] [CrossRef] [Green Version]
- Alberts-Grill, N.; Rezvan, A.; Son, D.J.; Qiu, H.; Kim, C.W.; Kemp, M.L.; Weyand, C.M.; Jo, H. Dynamic immune cell accumulation during flow-induced atherogenesis in mouse carotid artery: An expanded flow cytometry method. Arter. Thromb. Vasc. Biol. 2012, 32, 623–632. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fluorophore | Antibody | Marker | Clone | Staining Concentration * | Vendor |
|---|---|---|---|---|---|
| BB700 | CD8 | Cytotoxic T cells | 53–6.7 | 0.5 | BD Biosciences (Franklin Lakes, NJ, USA) |
| BB515 | CD43 | Monocyte subsets | S7 | 5 | BD Biosciences |
| AF700 | CD4 | Helper T cells | RM4-5 | 3 | BD Biosciences |
| A647 | CD64 | Monocytes and macrophages | X54-5/7.1 | 1.25 | BD Biosciences |
| PE/Cy7 | MHCII | Professional antigen-presenting cells | M5/114.15.2 | 3 | Invitrogen (Waltham, MA, USA) |
| PE/CF594 | SiglecF | Eosinophils | E50-2440 | 0.6 | BD Biosciences |
| PE | Tim4 | Macrophage subsets | RMT4-54 | 2.5 | BD Biosciences |
| BV786 | NK1.1 | Natural killer cells | PK136 | 1 | BD Biosciences |
| BV711 | Ly6G | Neutrophils | 1A8 | 1 | BD Biosciences |
| BV480 | Live/Dead | Viability dye | v500 | 1 μL/2 mL | eBioscience (San Diego, CA, USA) |
| BV421 | Ly6C | Monocytes and macrophages | HK1.4 | 2.5 | BioLegend (San Diego, CA, USA) |
| BUV805 | CD45 | Leukocytes | 30-F11 | 0.25 | BD Biosciences |
| BUV737 | CD11b | Innate immune cells | M1/70 | 3 | BD Biosciences |
| BUV563 | CD19 | B cells | 1D3 | 1.25 | BD Biosciences |
| BUV395 | CD11c | Dendritic cells, monocytes, macrophages, granulocytes, NK cells, and subsets of B and T cells | HL3 | 5 | BD Biosciences |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pamulapati, V.; Cuda, C.M.; Smith, T.L.; Jung, J.; Xiong, L.; Swaminathan, S.; Ho, K.J. Inflammatory Cell Dynamics after Murine Femoral Artery Wire Injury: A Multi-Parameter Flow Cytometry-Based Analysis. Cells 2023, 12, 689. https://doi.org/10.3390/cells12050689
Pamulapati V, Cuda CM, Smith TL, Jung J, Xiong L, Swaminathan S, Ho KJ. Inflammatory Cell Dynamics after Murine Femoral Artery Wire Injury: A Multi-Parameter Flow Cytometry-Based Analysis. Cells. 2023; 12(5):689. https://doi.org/10.3390/cells12050689
Chicago/Turabian StylePamulapati, Vivek, Carla M. Cuda, Tracy L. Smith, Jonathan Jung, Liqun Xiong, Suchitra Swaminathan, and Karen J. Ho. 2023. "Inflammatory Cell Dynamics after Murine Femoral Artery Wire Injury: A Multi-Parameter Flow Cytometry-Based Analysis" Cells 12, no. 5: 689. https://doi.org/10.3390/cells12050689