
Effects of IL-11/IL-11 Receptor Alpha on Proliferation and Steroidogenesis in Ovarian Granulosa Cells of Dairy Cows
 and
and 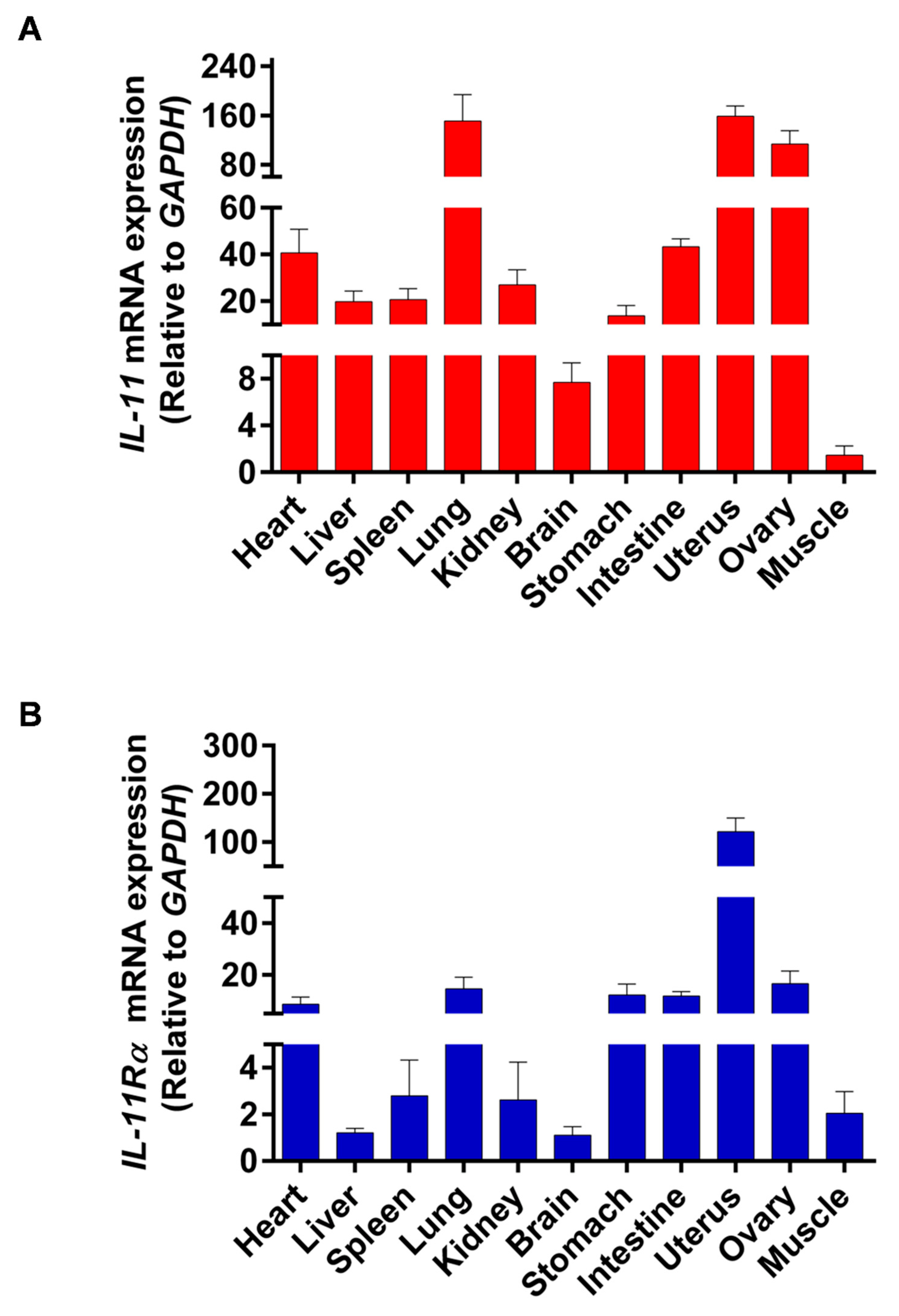{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Ethics Statement
2.2. Tissue Collection
2.3. Cell Culture
2.4. Interference Fragment Synthesis and Transfection
2.5. Cell Viability Assay
2.6. Cell Apoptosis Analysis
2.7. Cell Cycle Assay
2.8. Immunofluorescence Staining
2.9. Immunohistochemistry Staining
2.10. Hormone Detection
2.11. Quantitative Real-Time PCR (qRT-PCR) Assay
2.12. Western Blotting Analysis
2.13. Statistical Analysis
3. Results
3.1. Tissue Differential Expression of IL-11 and IL-11Rα in Bovine Organs
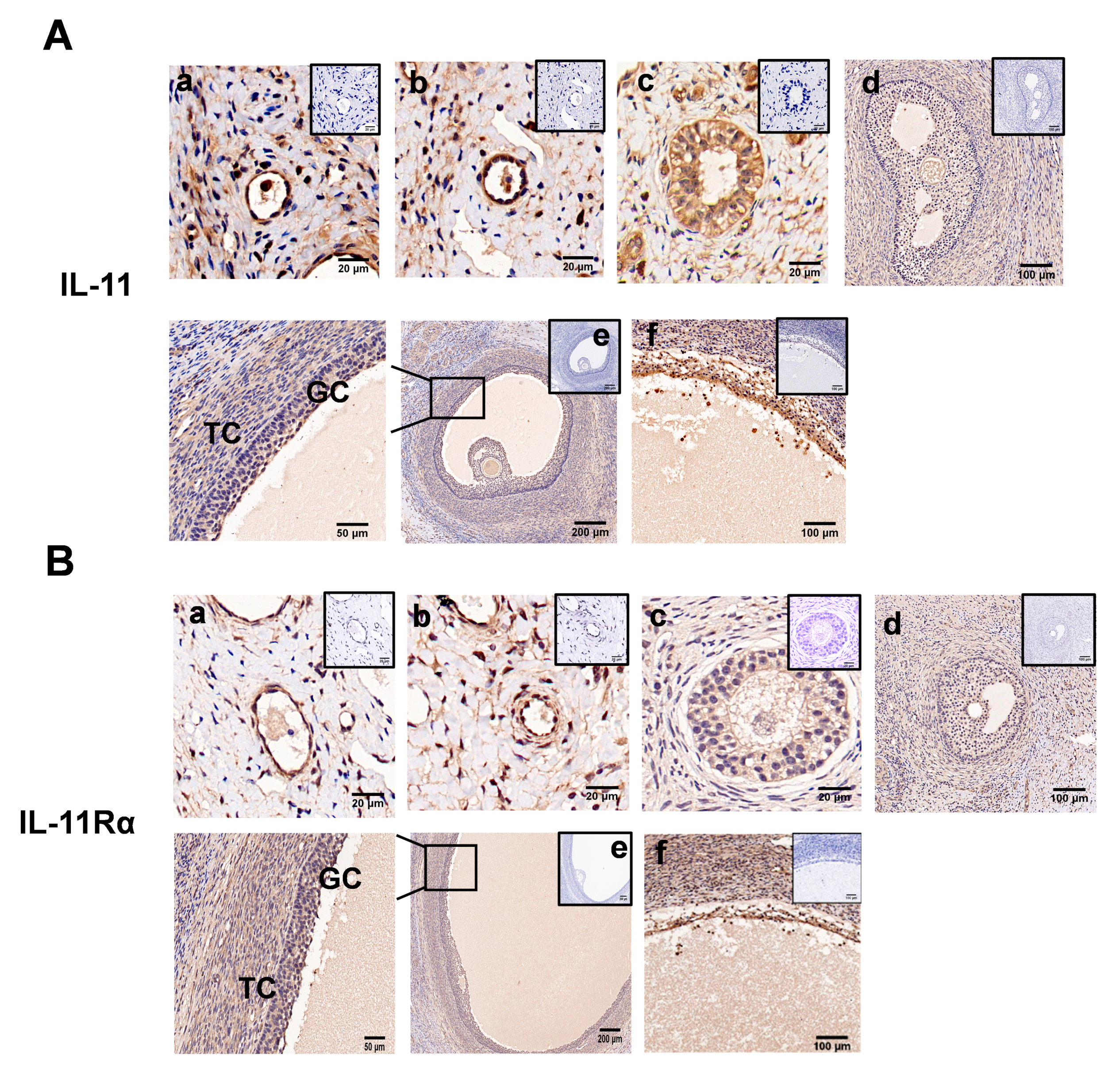
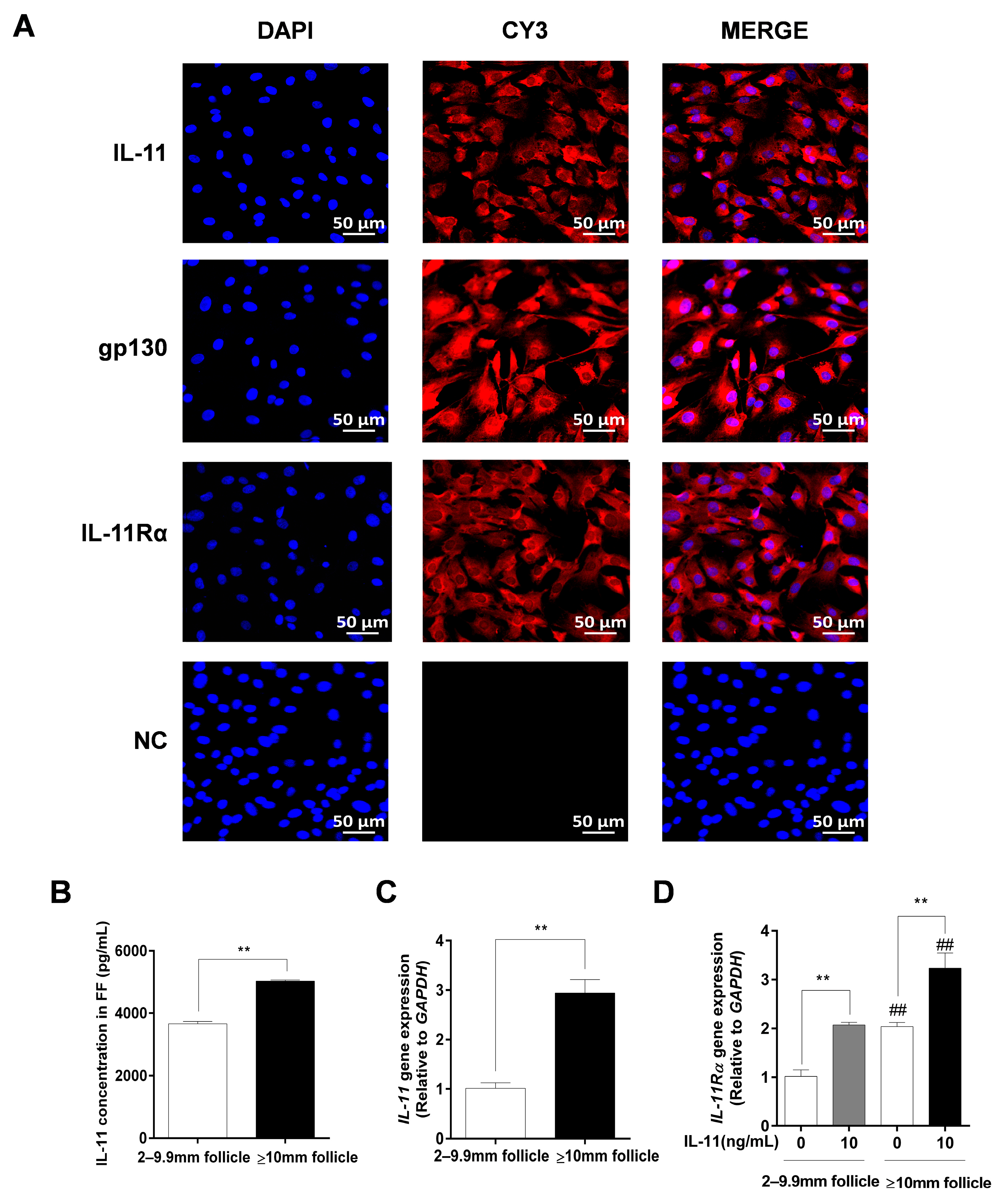
3.2. Localization of IL-11 System in Ovary and IL-11 Concentration in FF
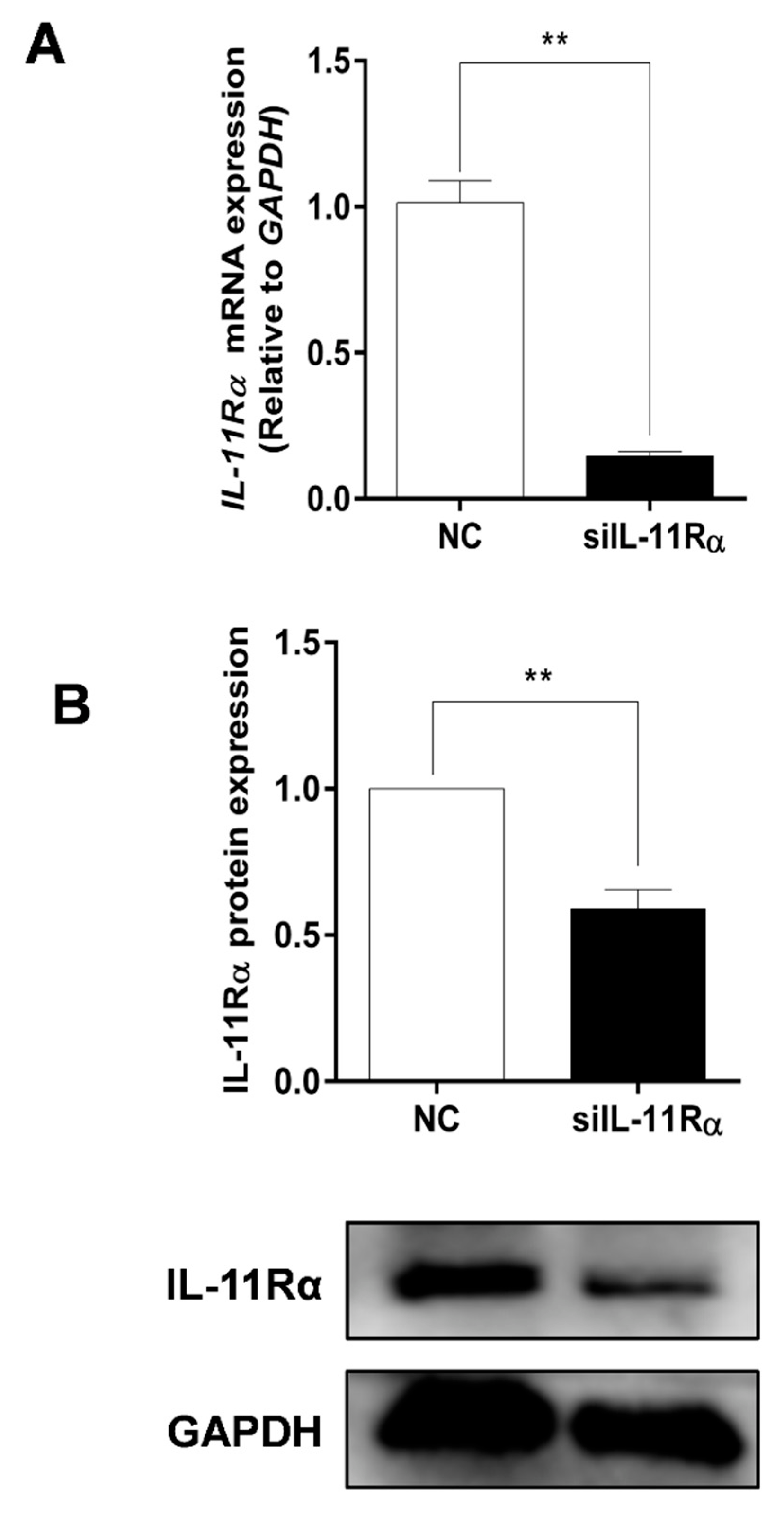
3.3. Detection of Knockdown Efficiency of IL-11Rα in Bovine GCs
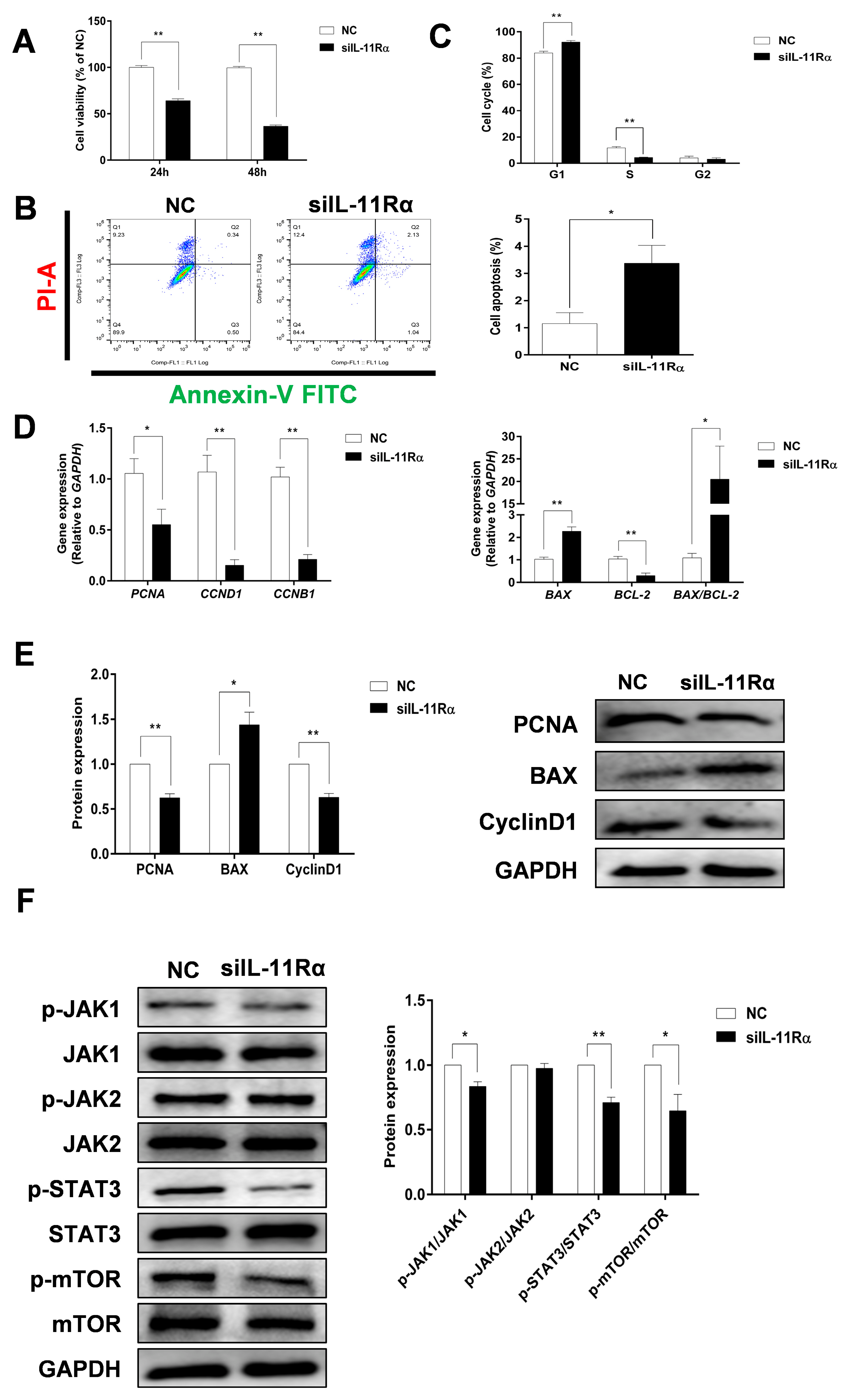
3.4. Effect of IL-11/IL-11Rα on Cell Proliferation and Apoptosis in Bovine GCs
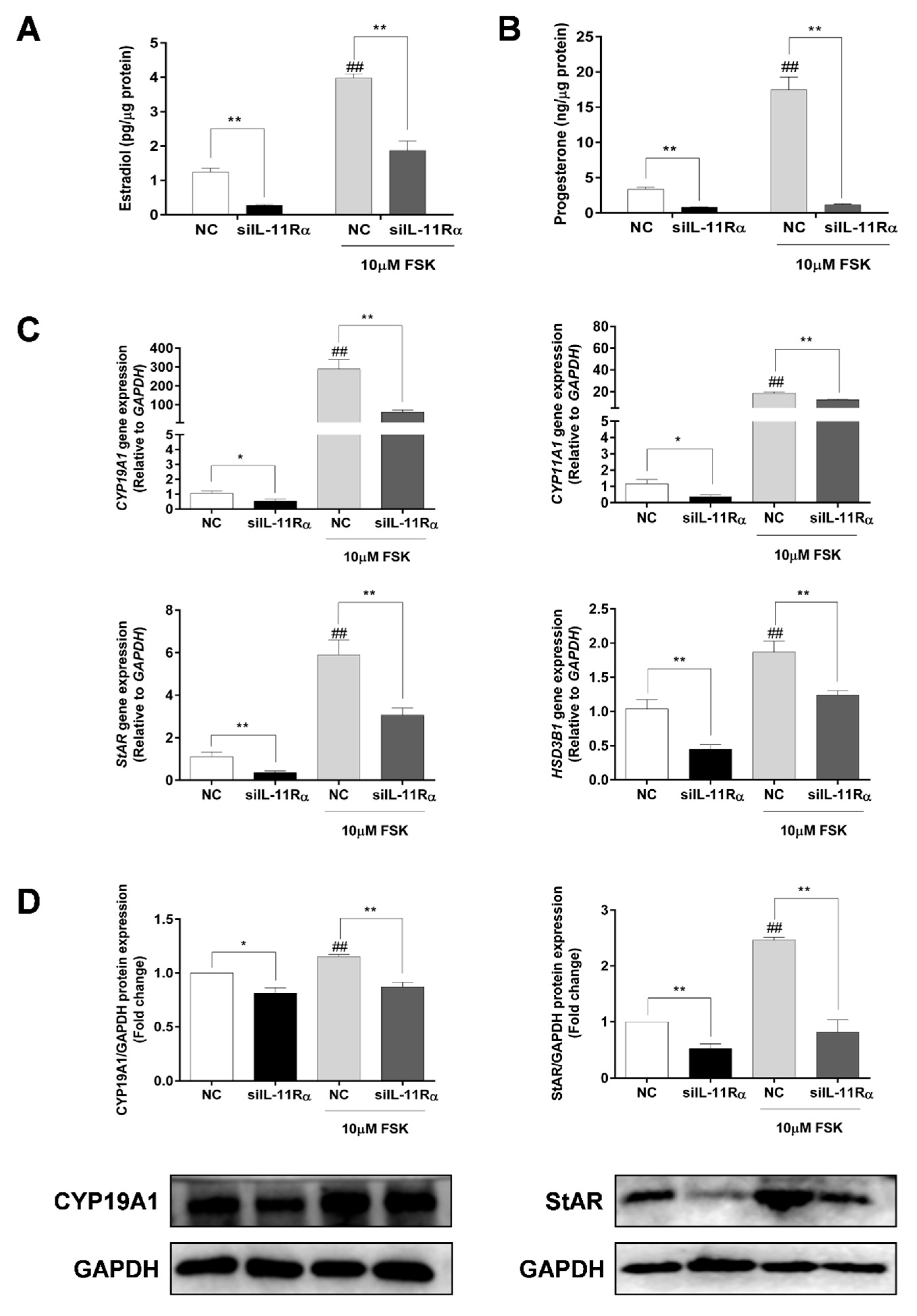
3.5. Effect of IL-11Rα Knockdown on Steroidogenesis in Bovine GCs
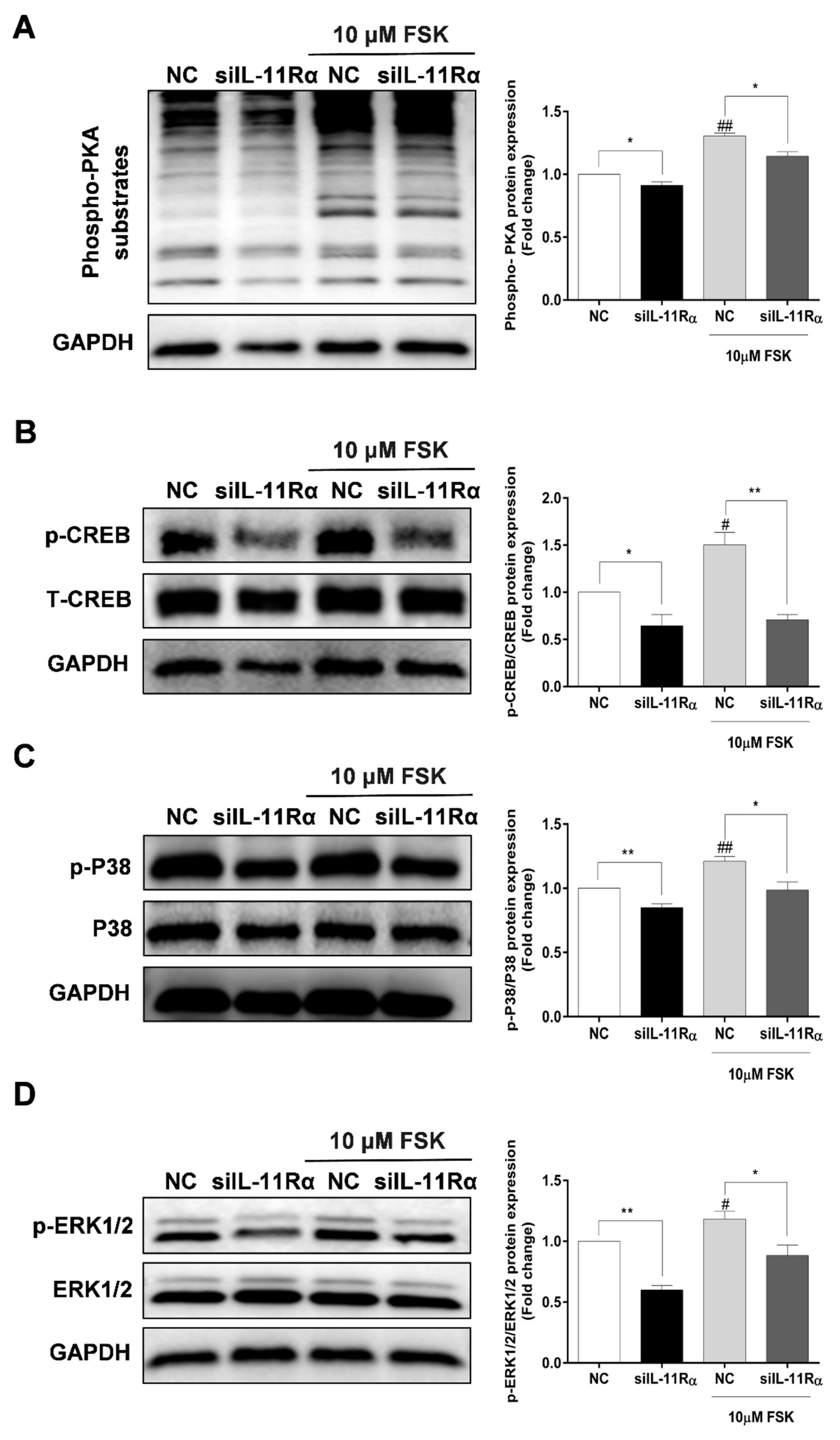
3.6. Involvement of PKA Signaling in Steroidogenesis after IL-11Rα Knockdown
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buyalos, R.P.; Watson, J.M.; Martinez-Maza, O. Detection of interleukin-6 in human follicular fluid. Fertil. Steril. 1992, 57, 1230–1234. [Google Scholar] [CrossRef] [PubMed]
- Gorospe, W.C.; Hughes, F.M., Jr.; Spangelo, B.L. Interleukin-6: Effects on and production by rat granulosa cells in vitro. Endocrinology 1992, 130, 1750–1752. [Google Scholar] [CrossRef] [PubMed]
- Machelon, V.; Emilie, D.; Lefevre, A.; Nome, F.; Durand-Gasselin, I.; Testart, J. Interleukin-6 biosynthesis in human preovulatory follicles: Some of its potential roles at ovulation. J. Clin. Endocrinol. Metab. 1994, 79, 633–642. [Google Scholar] [CrossRef]
- Kawasaki, F.; Kawano, Y.; Kosay Hasan, Z.; Narahara, H.; Miyakawa, I. The clinical role of interleukin-6 and interleukin-6 soluble receptor in human follicular fluids. Clin. Exp. Med. 2003, 3, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Wang, L.; Wang, X.; Wang, X.; Yang, Z.; Li, J. IL-6 Promotes FSH-Induced VEGF Expression Through JAK/STAT3 Signaling Pathway in Bovine Granulosa Cells. Cell. Physiol. Biochem. 2017, 44, 293–302. [Google Scholar] [CrossRef]
- Suen, Y.; Chang, M.; Lee, S.M.; Buzby, J.S.; Cairo, M.S. Regulation of interleukin-11 protein and mRNA expression in neonatal and adult fibroblasts and endothelial cells. Blood 1994, 84, 4125–4134. [Google Scholar] [CrossRef] [Green Version]
- Kiessling, S.; Muller-Newen, G.; Leeb, S.N.; Hausmann, M.; Rath, H.C.; Strater, J.; Spottl, T.; Schlottmann, K.; Grossmann, J.; Montero-Julian, F.A.; et al. Functional expression of the interleukin-11 receptor alpha-chain and evidence of antiapoptotic effects in human colonic epithelial cells. J. Biol. Chem. 2004, 279, 10304–10315. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.R.; Bennett, F.; Calvetti, J.A.; Kelleher, K.; Wood, C.R.; O’Hara, R.M., Jr.; Leary, A.C.; Sibley, B.; Clark, S.C.; Williams, D.A.; et al. Molecular cloning of a cDNA encoding interleukin 11, a stromal cell-derived lymphopoietic and hematopoietic cytokine. Proc. Natl. Acad. Sci. USA 1990, 87, 7512–7516. [Google Scholar] [CrossRef] [Green Version]
- Robb, L.; Li, R.; Hartley, L.; Nandurkar, H.H.; Koentgen, F.; Begley, C.G. Infertility in female mice lacking the receptor for interleukin 11 is due to a defective uterine response to implantation. Nat. Med. 1998, 4, 303–308. [Google Scholar] [CrossRef]
- Putoczki, T.; Ernst, M. More than a sidekick: The IL-6 family cytokine IL-11 links inflammation to cancer. J. Leukoc. Biol. 2010, 88, 1109–1117. [Google Scholar] [CrossRef]
- Yeh, C.C.; Chao, K.C.; Huang, S.J. Innate immunity, decidual cells, and preeclampsia. Reprod. Sci. 2013, 20, 339–353. [Google Scholar] [CrossRef] [Green Version]
- Barton, V.A.; Hall, M.A.; Hudson, K.R.; Heath, J.K. Interleukin-11 signals through the formation of a hexameric receptor complex. J. Biol. Chem. 2000, 275, 36197–36203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinrich, P.C.; Behrmann, I.; Müller-Newen, G.; Schaper, F.; Graeve, L. Interleukin-6-type cytokine signalling through the gp130/Jak/STAT pathway. Biochem. J. 1998, 334 Pt 2, 297–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbers, C.; Hermanns, H.M.; Schaper, F.; Müller-Newen, G.; Grötzinger, J.; Rose-John, S.; Scheller, J. Plasticity and cross-talk of interleukin 6-type cytokines. Cytokine Growth Factor Rev. 2012, 23, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Matadeen, R.; Hon, W.C.; Heath, J.K.; Jones, E.Y.; Fuller, S. The dynamics of signal triggering in a gp130-receptor complex. Structure 2007, 15, 441–448. [Google Scholar] [CrossRef] [Green Version]
- Bollrath, J.; Phesse, T.J.; von Burstin, V.A.; Putoczki, T.; Bennecke, M.; Bateman, T.; Nebelsiek, T.; Lundgren-May, T.; Canli, O.; Schwitalla, S.; et al. gp130-mediated Stat3 activation in enterocytes regulates cell survival and cell-cycle progression during colitis-associated tumorigenesis. Cancer Cell 2009, 15, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Gibson, D.L.; Montero, M.; Ropeleski, M.J.; Bergstrom, K.S.; Ma, C.; Ghosh, S.; Merkens, H.; Huang, J.; Månsson, L.E.; Sham, H.P.; et al. Interleukin-11 reduces TLR4-induced colitis in TLR2-deficient mice and restores intestinal STAT3 signaling. Gastroenterology 2010, 139, 1277–1288. [Google Scholar] [CrossRef]
- Nishina, T.; Deguchi, Y.; Ohshima, D.; Takeda, W.; Ohtsuka, M.; Shichino, S.; Ueha, S.; Yamazaki, S.; Kawauchi, M.; Nakamura, E.; et al. Interleukin-11-expressing fibroblasts have a unique gene signature correlated with poor prognosis of colorectal cancer. Nat. Commun. 2021, 12, 2281. [Google Scholar] [CrossRef]
- Yin, T.; Yang, Y.C. Mitogen-activated protein kinases and ribosomal S6 protein kinases are involved in signaling pathways shared by interleukin-11, interleukin-6, leukemia inhibitory factor, and oncostatin M in mouse 3T3-L1 cells. J. Biol. Chem. 1994, 269, 3731–3738. [Google Scholar] [CrossRef]
- Sun, R.; Chen, C.; Deng, X.; Wang, F.; Song, S.; Cai, Q.; Wang, J.; Zhang, T.; Shi, M.; Ke, Q.; et al. IL-11 mediates the Radioresistance of Cervical Cancer Cells via the PI3K/Akt Signaling Pathway. J. Cancer 2021, 12, 4638–4647. [Google Scholar] [CrossRef]
- Fuhrer, D.K.; Yang, Y.C. Activation of Src-family protein tyrosine kinases and phosphatidylinositol 3-kinase in 3T3-L1 mouse preadipocytes by interleukin-11. Exp. Hematol. 1996, 24, 195–203. [Google Scholar] [PubMed]
- Branisteanu, I.; Pijnenborg, R.; Spiessens, C.; Van der Auwera, I.; Keith, J.C., Jr.; Van Assche, F.A. Detection of immunoreactive interleukin-11 in human follicular fluid: Correlations with ovarian steroid, insulin-like growth factor I levels, and follicular maturity. Fertil. Steril. 1997, 67, 1054–1058. [Google Scholar] [CrossRef] [Green Version]
- Jang, Y.J.; Park, J.I.; Jeong, S.E.; Seo, Y.M.; Dam, P.T.M.; Seo, Y.W.; Choi, B.C.; Song, S.J.; Chun, S.Y.; Cho, M.K. Regulation of interleukin-11 expression in ovulatory follicles of the rat ovary. Reprod. Fertil. Dev. 2017, 29, 2437–2445. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.; Pan, X.; Zhao, K.; Gao, W.; Liu, J.; Deng, T.; Qin, W. The Effect of Interleukin-6 (IL-6), Interleukin-11 (IL-11), Signal Transducer and Activator of Transcription 3 (STAT3), and AKT Signaling on Adipocyte Proliferation in a Rat Model of Polycystic Ovary Syndrome. Med. Sci. Monit. 2019, 25, 7218–7227. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhang, C.; Wang, L.; Huang, X.; Zhang, J.; He, Y.; Chen, L.; Li, J. Successful reversal of ovarian hyperstimulation syndrome in a mouse model by rapamycin, an mTOR pathway inhibitor. Mol. Hum. Reprod. 2019, 25, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Winship, A.; Menkhorst, E.; Van Sinderen, M.; Dimitriadis, E. Interleukin 11: Similar or opposite roles in female reproduction and reproductive cancer? Reprod. Fertil. Dev. 2016, 28, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Lokau, J.; Kespohl, B.; Kirschke, S.; Garbers, C. The role of proteolysis in interleukin-11 signaling. Biochim. Biophys. Acta Mol. Cell Res. 2022, 1869, 119135. [Google Scholar] [CrossRef] [PubMed]
- Kuenzler, K.A.; Pearson, P.Y.; Schwartz, M.Z. IL-11 pretreatment reduces cell death after intestinal ischemia-reperfusion. J. Surg. Res. 2002, 108, 268–272. [Google Scholar] [CrossRef]
- Yu, J.; Feng, Z.; Tan, L.; Pu, L.; Kong, L. Interleukin-11 protects mouse liver from warm ischemia/reperfusion (WI/Rp) injury. Clin. Res. Hepatol. Gastroenterol. 2016, 40, 562–570. [Google Scholar] [CrossRef]
- Totty, M.L.; Morrell, B.C.; Spicer, L.J. Fibroblast growth factor 9 (FGF9) regulation of cyclin D1 and cyclin-dependent kinase-4 in ovarian granulosa and theca cells of cattle. Mol. Cell. Endocrinol. 2017, 440, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Li, C.; Chen, S.; Sun, L.; Li, H.; Chen, L.; Zhou, X. Effect of inhibin A on proliferation of porcine granulosa cells in vitro. Theriogenology 2018, 114, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.H.; Zhu, Z.; Wakefield, M.R.; Xiao, H.; Bai, Q.; Fang, Y. The role of IL-11 in immunity and cancer. Cancer Lett. 2016, 373, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Ishihara, K.; Hibi, M. Roles of STAT3 in mediating the cell growth, differentiation and survival signals relayed through the IL-6 family of cytokine receptors. Oncogene 2000, 19, 2548–2556. [Google Scholar] [CrossRef] [PubMed]
- Siveen, K.S.; Sikka, S.; Surana, R.; Dai, X.; Zhang, J.; Kumar, A.P.; Tan, B.K.; Sethi, G.; Bishayee, A. Targeting the STAT3 signaling pathway in cancer: Role of synthetic and natural inhibitors. Biochim. Biophys. Acta 2014, 1845, 136–154. [Google Scholar] [CrossRef] [Green Version]
- Shien, K.; Papadimitrakopoulou, V.A.; Ruder, D.; Behrens, C.; Shen, L.; Kalhor, N.; Song, J.; Lee, J.J.; Wang, J.; Tang, X.; et al. JAK1/STAT3 Activation through a Proinflammatory Cytokine Pathway Leads to Resistance to Molecularly Targeted Therapy in Non-Small Cell Lung Cancer. Mol. Cancer Ther. 2017, 16, 2234–2245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keck, C.; Rajabi, Z.; Pfeifer, K.; Bettendorf, H.; Brandstetter, T.; Breckwoldt, M. Expression of interleukin-6 and interleukin-6 receptors in human granulosa lutein cells. Mol. Hum. Reprod. 1998, 4, 1071–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmassi, A.; Lü, S.; Hedderich, J.; Oettinghaus, C.; Jonat, W.; Mettler, L. Interaction of interleukin-6 on human granulosa cell steroid secretion. J. Endocrinol. 2001, 170, 471–478. [Google Scholar] [CrossRef] [Green Version]
- Van der Hoek, K.H.; Woodhouse, C.M.; Brännström, M.; Norman, R.J. Effects of interleukin (IL)-6 on luteinizing hormone- and IL-1beta-induced ovulation and steroidogenesis in the rat ovary. Biol. Reprod. 1998, 58, 1266–1271. [Google Scholar] [CrossRef] [Green Version]
- Pitzel, L.; Jarry, H.; Wuttke, W. Effects and interactions of prostaglandin F2 alpha, oxytocin, and cytokines on steroidogenesis of porcine luteal cells. Endocrinology 1993, 132, 751–756. [Google Scholar] [CrossRef]
- Alpizar, E.; Spicer, L.J. Effects of interleukin-6 on proliferation and follicle-stimulating hormone-induced estradiol production by bovine granulosa cells in vitro: Dependence on size of follicle. Biol. Reprod. 1994, 50, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Carlone, D.L.; Richards, J.S. Functional interactions, phosphorylation, and levels of 3′,5′-cyclic adenosine monophosphate-regulatory element binding protein and steroidogenic factor-1 mediate hormone-regulated and constitutive expression of aromatase in gonadal cells. Mol. Endocrinol. 1997, 11, 292–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunzicker-Dunn, M.; Maizels, E.T. FSH signaling pathways in immature granulosa cells that regulate target gene expression: Branching out from protein kinase A. Cell. Signal. 2006, 18, 1351–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, P.J.; Fujimoto, Y.; Yamauchi, N.; Hattori, M.A. Real-time monitoring of cAMP response element binding protein signaling in porcine granulosa cells modulated by ovarian factors. Mol. Cell. Biochem. 2006, 290, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Maizels, E.T.; Cottom, J.; Jones, J.C.; Hunzicker-Dunn, M. Follicle stimulating hormone (FSH) activates the p38 mitogen-activated protein kinase pathway, inducing small heat shock protein phosphorylation and cell rounding in immature rat ovarian granulosa cells. Endocrinology 1998, 139, 3353–3356. [Google Scholar] [CrossRef]
- Yu, F.Q.; Han, C.S.; Yang, W.; Jin, X.; Hu, Z.Y.; Liu, Y.X. Activation of the p38 MAPK pathway by follicle-stimulating hormone regulates steroidogenesis in granulosa cells differentially. J. Endocrinol. 2005, 186, 85–96. [Google Scholar] [CrossRef]
- Fan, H.Y.; Liu, Z.; Shimada, M.; Sterneck, E.; Johnson, P.F.; Hedrick, S.M.; Richards, J.S. MAPK3/1 (ERK1/2) in ovarian granulosa cells are essential for female fertility. Science 2009, 324, 938–941. [Google Scholar] [CrossRef] [Green Version]
- Bai, L.; Chang, H.M.; Cheng, J.C.; Chu, G.; Leung, P.C.K.; Yang, G. Lithium chloride inhibits StAR and progesterone production through GSK-3β and ERK1/2 signaling pathways in human granulosa-lutein cells. Mol. Cell. Endocrinol. 2018, 461, 89–99. [Google Scholar] [CrossRef]
- Hinshelwood, M.M.; Michael, M.D.; Simpson, E.R. The 5′-flanking region of the ovarian promoter of the bovine CYP19 gene contains a deletion in a cyclic adenosine 3′,5′-monophosphate-like responsive sequence. Endocrinology 1997, 138, 3704–3710. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Sun, P.; Lv, C.; Zhao, X.; Liu, M.; Zhou, Q.; Tang, J.; Yang, L.; Liang, A. Effects of IL-11/IL-11 Receptor Alpha on Proliferation and Steroidogenesis in Ovarian Granulosa Cells of Dairy Cows. Cells 2023, 12, 673. https://doi.org/10.3390/cells12040673
Wu H, Sun P, Lv C, Zhao X, Liu M, Zhou Q, Tang J, Yang L, Liang A. Effects of IL-11/IL-11 Receptor Alpha on Proliferation and Steroidogenesis in Ovarian Granulosa Cells of Dairy Cows. Cells. 2023; 12(4):673. https://doi.org/10.3390/cells12040673
Chicago/Turabian StyleWu, Hanxiao, Peihao Sun, Ce Lv, Xinzhe Zhao, Mingxiao Liu, Qunli Zhou, Jiaomei Tang, Liguo Yang, and Aixin Liang. 2023. "Effects of IL-11/IL-11 Receptor Alpha on Proliferation and Steroidogenesis in Ovarian Granulosa Cells of Dairy Cows" Cells 12, no. 4: 673. https://doi.org/10.3390/cells12040673