
Impact of Chronic Multi-Generational Exposure to an Environmentally Relevant Atrazine Concentration on Testicular Development and Function in Mice
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Puberty Determination
2.3. Post-Mortem Tissue Collection and DEXA Analysis
2.4. Sperm Analysis
2.5. RNA Isolation and cDNA Synthesis
2.6. Quantitative RT-PCR
2.7. Histology of Testis Sections
2.8. Immunohistochemistry of Testis Sections
2.9. Statistical Analyses
3. Results
3.1. ATZ Exposure Did Not Modify Litter Characteristics or the Timing of Puberty Initiation
3.2. ATZ Exposure Did Not Affect Body and Organ Weighs
3.3. ATZ Exposure Did Not Affect DEXA Parameters
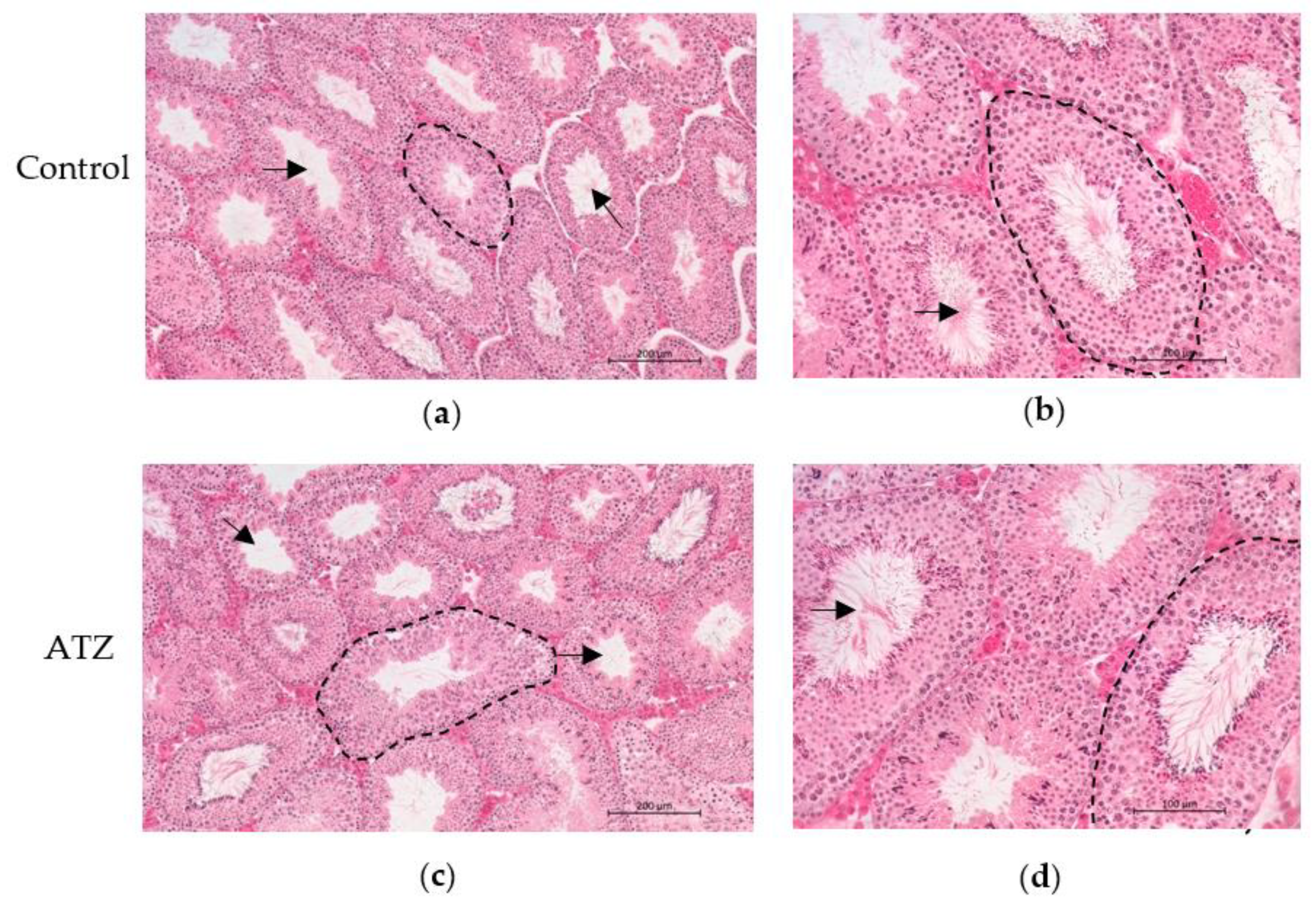
3.4. ATZ Exposure Did Not Result in Gross Morphological Changes in the Testis
3.5. ATZ Exposure Did Not Affect Sperm Characteristics
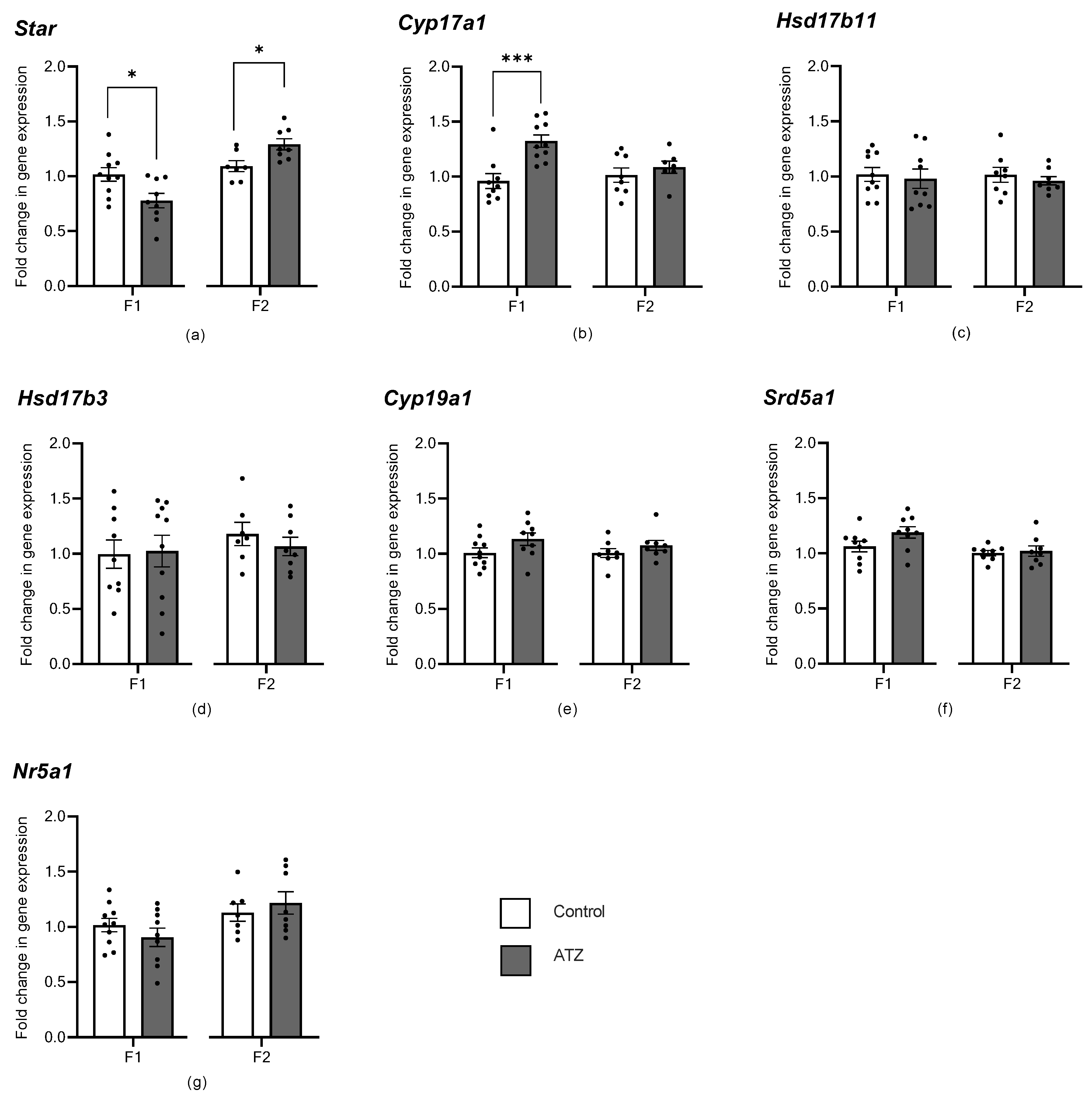
3.6. ATZ Exposure Altered Early Steroidogenic Gene Expression
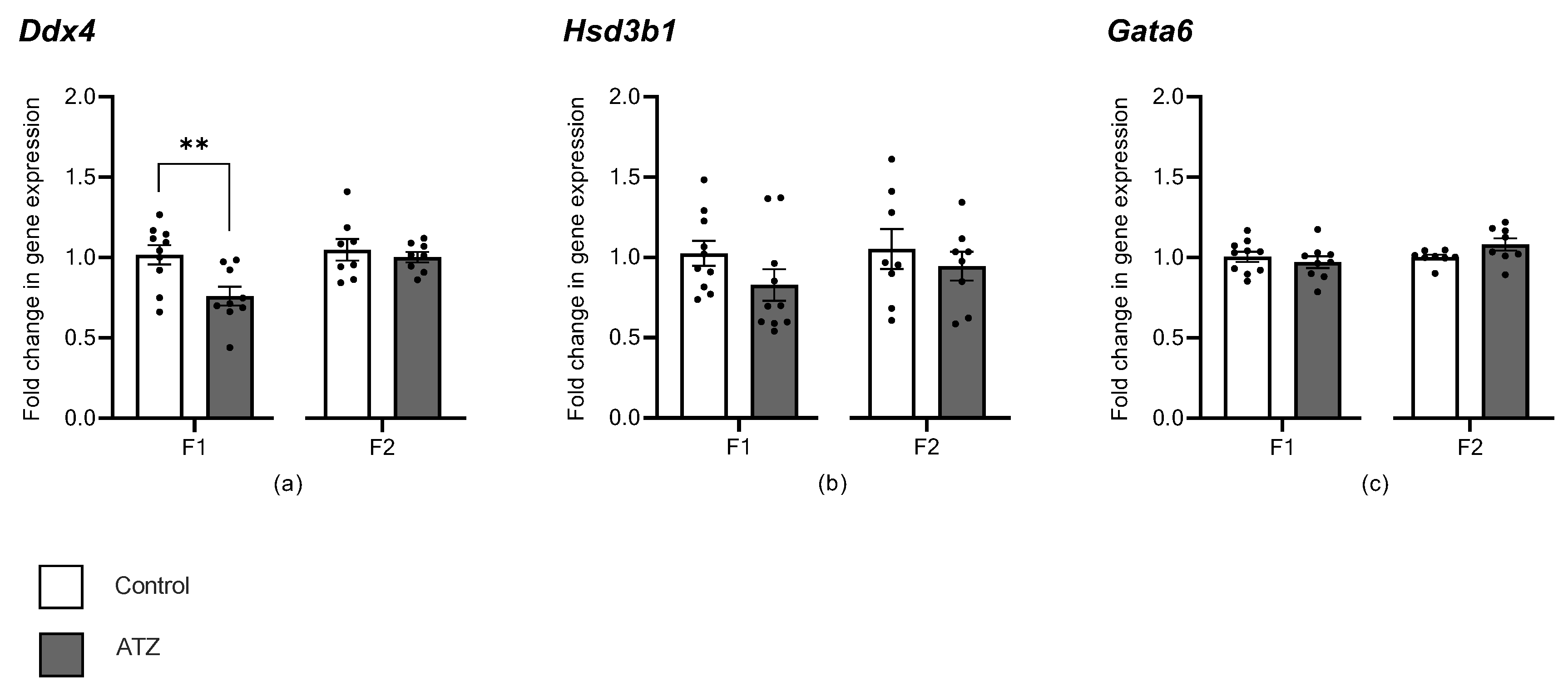
3.7. ATZ Exposure Altered Expression of Germ Cell Marker, Ddx4
3.8. ATZ Exposure Did Not Affect Seminiferous Tubule Measurements
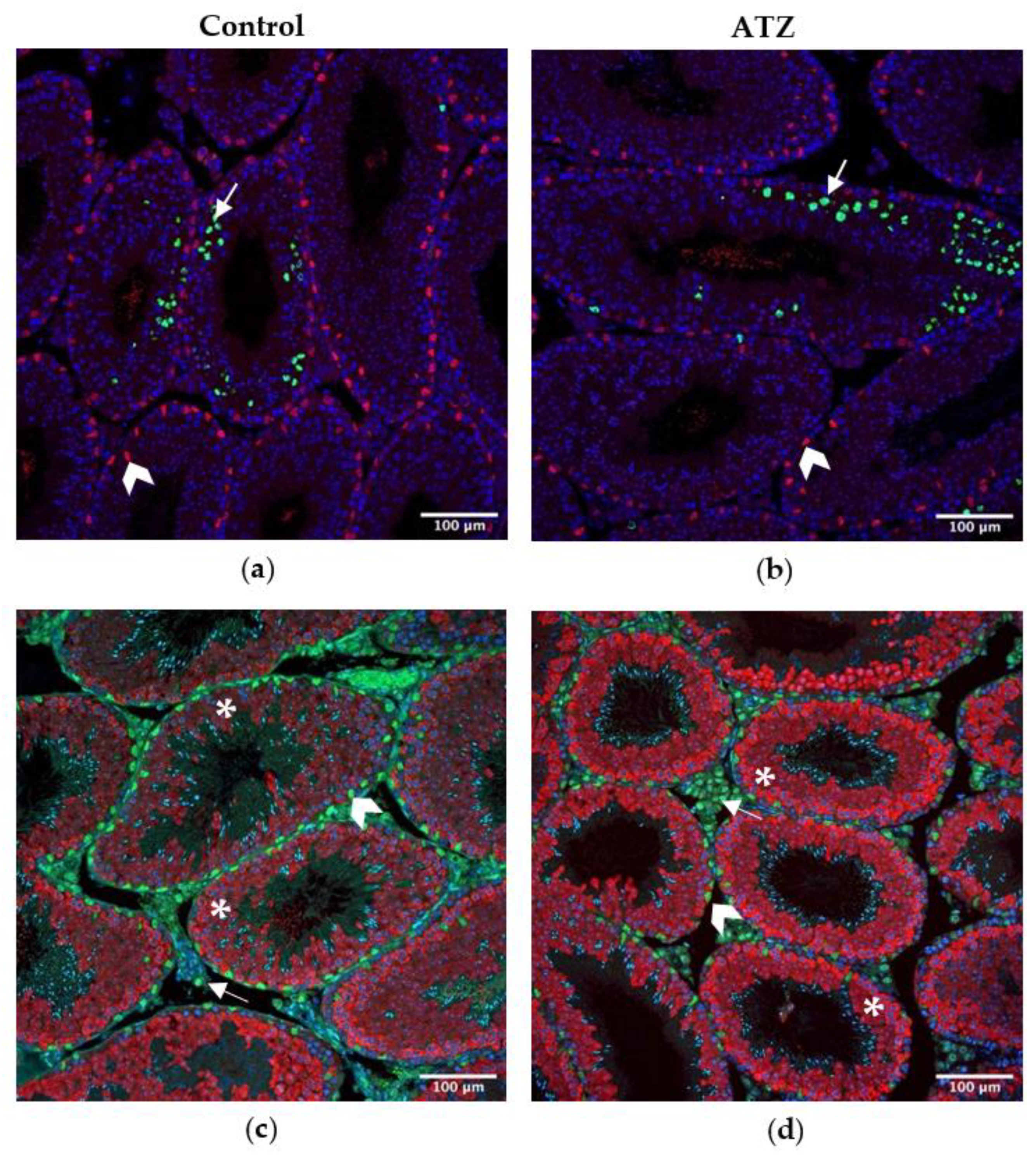
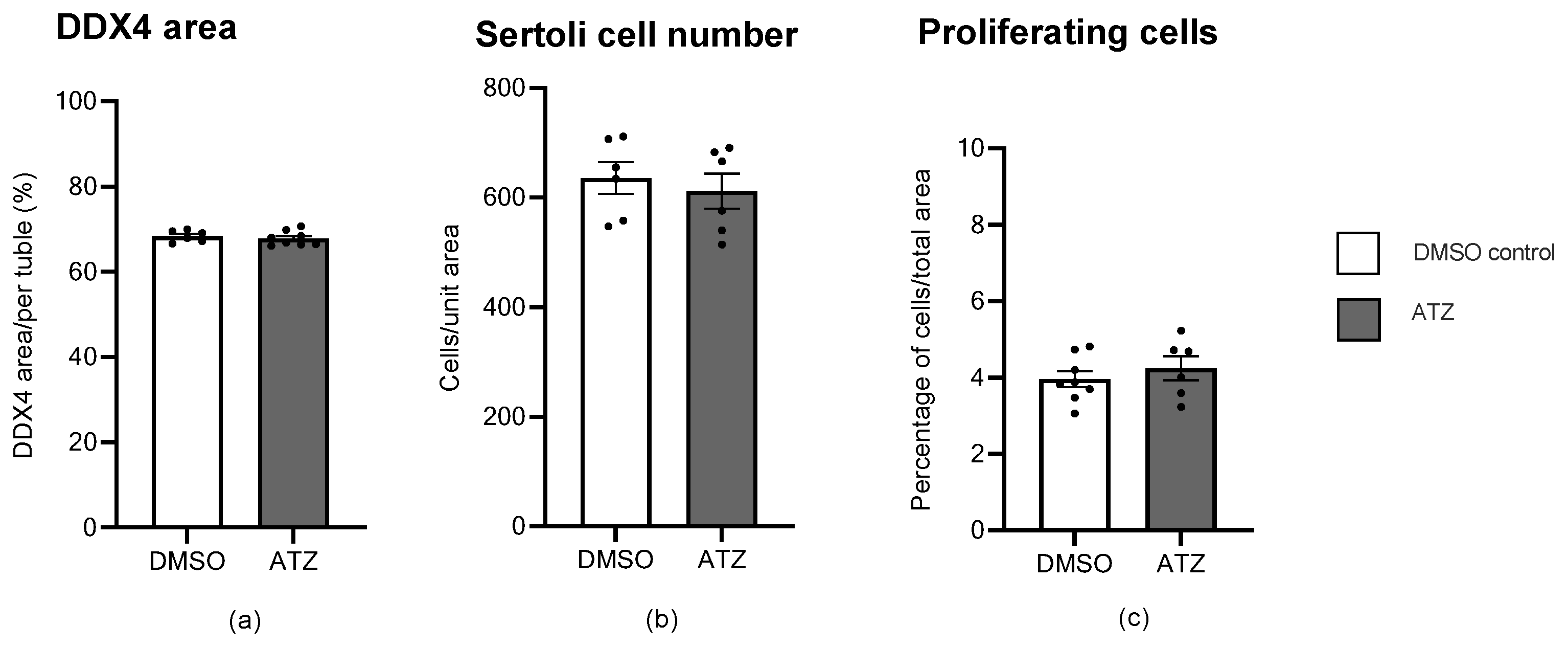
3.9. ATZ Exposure Did Not Affect the Area of Ddx4 Positive Cells, Sertoli Cell Number or the Number of Proliferating Cells
4. Discussion
4.1. Atrazine Affects Steroidogenesis
4.2. Atrazine’s Impact on Testis Cell Sub-Types
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Primer Sequence | Product Length | Reference |
|---|---|---|---|---|
| Tbp | NM_013684.3 | Forward—ACGGACAACTGCGTTGATTTT Reverse—ACTTAGCTGGGAAGCCCAAC | 128 | [19] |
| β-actin | NM_007393.5 | Forward—CCACTGTCGAGTCGCGT Reverse—GTCATCCATGGCGAACTGGT | 91 | [19] |
| Rpl19 | NM_001159483.1 | Forward—AGGCTACAGAAGAGGCTTGC Reverse—GGCATTGGCGATTTCATTGGT | 93 | N/A |
| Ddx4 | NM_010029.2 | Forward—CCAAGATCAGGGGACACAGC Reverse—CTTTGGTAAGTGTCACCATTGC | 90 | N/A |
| Cyp17a1 | NM_007809.3 | Forward—CAATGACCGGACTCACCTCC Reverse—GGCAAACTCTCCAATGCTGG | 124 | N/A |
| Cyp19a1 | NM_001348171.1 | Forward—CGGGCTACGTGGATGTGTT Reverse—GAGCTTGCCAGGCGTTAAAG | 135 | N/A |
| Gata6 | NM_010258.3 | Forward—TTGCTCCGGTAACAGCAGTG Reverse—GTGGTCGCTTGTGTAGAAGGA | 105 | [59] |
| Hsd17β3 | NM_008291.3 | Forward—ACAAGATGACCAAGACCGCC Reverse—CCACAGGATTCAGCTCCGAT | 76 | N/A |
| Hsd17β11 | NM_053262.3 | Forward—CCTTGGGACGAACAGGAGTG Reverse—CCCGTGCATGAGATGTTCCA | 134 | [19] |
| Hsd3β1 | NM_001304800.1 | Forward—CCTCCTAAGGGTTACCCTATATCATACCAGCT Reverse—GTCTCCTTCCAACACTGTCACCTTGG | 247 | [60] |
| Nr5a1 | NM_139051.3 | Forward—CAGGAGCCCTGTCACTAAGC Reverse—ATGGTTAGAAGCCCCCTTTGG | 184 | [19] |
| Srd5a1 | NM_175283.3 | Forward—GTTTGCTCTGTTCACCCTGTG Reverse—TGGACAGCACACTAAAGCAGG | 132 | [19] |
| Star | NM_011485.5 | Forward—TAAGGCAGCGCACTTGATCT Reverse—ACACAGCTTGAACGTAGCGA | 119 | N/A |
References
- United Nations Environmental Programme & World Health Organization. State of the Science of Endocrine Disrupting Chemicals- 2012; Bergman, A., Heindel, J.J., Jobling, S., Kidd, K.A., Zoeller, R.A., Eds.; United Nations Environmental Programme & World Health Organization: Geneva, Switzerland, 2013; pp. 1–289. [Google Scholar]
- Radcliffe, J.C. Pesticide Use in Australia: A Review Undertaken by the Australian Academy of Technological Sciences and Engineering; Australian Academy of Technological Sciences and Engineering: Melbourne, Australia, 2002; pp. 1–319. [Google Scholar]
- Wang, M. Visualizing pesticide usage in the United States from 1992 to 2009. Environ. Plan. A 2016, 48, 455–457. [Google Scholar] [CrossRef] [Green Version]
- Woodruff, T.J. Bridging epidemiology and model organisms to increase understanding of endocrine disrupting chemicals and human health effects. J. Steroid Biochem. Mol. Biol. 2011, 12, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Australian Pesticides and Veterinary Medicines Authority. Atrazine Final Review Report and Regulatory Decision; Australian Government: Canberra, Australia, 2004; pp. 1–47.
- Australian Pesticides and Veterinary Medicines Authority. Atrazine Final Review Report and Regulatory Decision; Australian Government: Canberra, Australia, 2008; pp. 1–49.
- European Commission. Commission decision of 10 March 2004 concerning the non- inclusion of atrazine in Annex 1 to council directive 91/414/EEC and the withdrawal of authorisations for plant protection products containing this active substance. Off. J. Eur. Union. 2004, 248, 1–3. [Google Scholar]
- Allinson, G.; Zhang, P.; Bui, A.; Allinson, M.; Rose, G.; Marshall, S.; Pettigrove, V. Pesticide and trace metal occurrence and aquatic benchmark exceedances in surface waters and sediments of urban wetlands and retention ponds in Melbourne, Australia. Environ. Sci. Pollut. Res. Int. 2015, 22, 10214–10226. [Google Scholar] [CrossRef]
- Swan, S.H.; Kruse, R.L.; Liu, F.; Barr, D.B.; Drobnis, E.Z.; Redmon, J.B.; Wang, C.; Brazil, C.; Overstreet, J.W. Semen quality in relation to biomarkers of pesticide exposure. Environ. Health Perspect. 2003, 111, 1478–1484. [Google Scholar] [CrossRef] [Green Version]
- Kniewald, J.; Jakominic, M.; Tomljenovic, A.; Simic, B.; Romac, P.; Vranesic, D.; Kniewald, Z. Disorders of male rat reproductive tract under the influence of atrazine. J. Appl. Toxicol. 2000, 20, 61–68. [Google Scholar] [CrossRef]
- Abarikwu, S.O.; Adesiyan, A.C.; Oyeloja, T.O.; Oyeyemi, M.O.; Farombi, E.O. Changes in sperm characteristics and induction of oxidative stress in the testis and epididymis of experimental rats by a herbicide, atrazine. Arch. Environ. Contam. Toxicol. 2010, 58, 874–882. [Google Scholar] [CrossRef]
- Pogrmic, K.; Fa, S.; Dakic, V.; Kaisarevic, S.; Kovacevic, R. Atrazine oral exposure of peripubertal male rats downregulates steroidogenesis gene expression in Leydig cells. Toxicol. Sci. 2009, 111, 189–197. [Google Scholar] [CrossRef]
- Trentacoste, S.V.; Friedmann, A.S.; Youker, R.T.; Breckenridge, C.B.; Zirkin, B.R. Atrazine effects on testosterone levels and androgen-dependent reproductive organs in peripubertal male rats. J. Androl. 2001, 22, 142–148. [Google Scholar]
- Stoker, T.E.; Laws, S.C.; Guidici, D.L.; Cooper, R.L. The effect of atrazine on puberty in male wistar rats: An evaluation in the protocol for the assessment of pubertal development and thyroid function. Toxicol. Sci. 2000, 58, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Hayes, T.B.; Anderson, L.L.; Beasley, V.R.; de Solla, S.R.; Iguchi, T.; Ingraham, H.; Kestemont, P.; Kniewald, J.; Kniewald, Z.; Langlois, V.S.; et al. Demasculinization and feminization of male gonads by atrazine: Consistent effects across vertebrate classes. J. Steroid Biochem. Mol. Biol. 2011, 127, 64–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedmann, A.S. Atrazine inhibition of testosterone production in rat males following peripubertal exposure. Reprod. Toxicol. 2002, 16, 275–279. [Google Scholar] [CrossRef] [PubMed]
- McBirney, M.; King, S.E.; Pappalardo, M.; Houser, E.; Unkefer, M.; Nilsson, E.; Sadler-Riggleman, I.; Beck, D.; Winchester, P.; Skinner, M.K. Atrazine induced epigenetic transgenerational inheritance of disease, lean phenotype and sperm epimutation pathology biomarkers. PLoS ONE 2017, 12, e0184306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kniewald, J.; Osredecki, V.; Gojmerac, T.; Zechner, V.; Kniewald, Z. Effect of s-triazine compounds on testosterone metabolism in the rat prostate. J. Appl. Toxicol. 1995, 15, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Harper, A.P.; Finger, B.J.; Green, M.P. Chronic atrazine exposure beginning prenatally impacts liver function and sperm concentration with multi-generational consequences in mice. Front. Endocrinol. 2020, 11, 580124. [Google Scholar] [CrossRef]
- Cook, L.E.; Finger, B.J.; Green, M.P.; Pask, A.J. Exposure to atrazine during puberty reduces sperm viability, increases weight gain and alters the expression of key metabolic genes in the liver of male mice. Reprod. Fertil. Dev. 2019, 31, 920–931. [Google Scholar] [CrossRef]
- Fang, Y.; Ni, C.B.; Dong, Y.Y.; Li, H.T.; Wu, S.W.; Li, X.H.; Lv, Y.; Huang, T.L.; Lian, Q.Q.; Ge, R.S. In utero exposure to atrazine disrupts rat fetal testis development. Front Pharmacol. 2018, 9, 1391. [Google Scholar] [CrossRef]
- Tan, H.; Wu, G.; Wang, S.; Lawless, J.; Sim, A.; Chen, D.; Zheng, Z. Prenatal exposure to atrazine induces cryptorchidism and hypospadias in F1 male mouse offspring. Birth Defects Res. 2021, 113, 469–484. [Google Scholar] [CrossRef]
- Pandey, N.; Maske, P.; Mote, C.; Dighe, V. Exposure to Atrazine through gestation and lactation period led to impaired sexual maturation and subfertility in F1 male rats with congenital deformities in F2 progeny. Food Chem. Toxicol. 2021, 157, 112586. [Google Scholar] [CrossRef]
- Hayes, T.B.; Collins, A.; Lee, M.; Mendoza, M.; Noriega, N.; Stuart, A.A.; Vonk, A. Hermaphroditic, demasculinized frogs after exposure to the herbicide atrazine at low ecologically relevant doses. Proc. Natl. Acad. Sci. USA 2002, 99, 5476–5480. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Wang, L.; Fu, Z. Oral exposure to atrazine modulates hormone synthesis and the transcription of steroidogenic genes in male peripubertal mice. Gen. Comp. Endocrinol. 2013, 184, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Holloway, A.C.; Anger, D.A.; Crankshaw, D.J.; Wu, M.; Foster, W.G. Atrazine-induced changes in aromatase activity in estrogen sensitive target tissues. J. Appl. Toxicol. 2008, 28, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Roberge, M.; Hakk, H.; Larsen, G. Atrazine is a competitive inhibitor of phosphodiesterase but does not affect the estrogen receptor. Toxicol. Lett. 2004, 154, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Kucka, M.; Pogrmic-Majkic, K.; Fa, S.; Stojilkovic, S.S.; Kovacevic, R. Atrazine acts as an endocrine disrupter by inhibiting cAMP-specific phosphodiesterase-4. Toxicol. Appl. Pharmacol. 2012, 265, 19–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hess, R.A.; Bunick, D.; Lee, K.-H.; Bahr, J.; Taylor, J.A.; Korach, K.S.; Lubahn, D.B. A role for oestrogens in the male reproductive system. Nature 1997, 390, 509–512. [Google Scholar] [CrossRef] [Green Version]
- Kinter, K.J.; Anekar, A.A. Biochemistry, Dihydrotestosterone. In StatPearls; StatPearl Publishing: Treasure Island, CA, USA, 2022. [Google Scholar]
- Hao, C.; Gely-Pernot, A.; Kervarrec, C.; Boudjema, M.; Becker, E.; Khil, P.; Tevosian, S.; Jegou, B.; Smagulova, F. Exposure to the widely used herbicide atrazine results in deregulation of global tissue-specific RNA transcription in the third generation and is associated with a global decrease of histone trimethylation in mice. Nucleic Acids Res. 2016, 44, 9784–9802. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, B.G.; Chen, H.; Folmer, J.; Liu, J.; Papadopoulos, V.; Zirkin, B.R. Gestational exposure to atrazine: Effects on the postnatal development of male offspring. J. Androl. 2008, 29, 304–311. [Google Scholar] [CrossRef]
- Govers, L.C.; Harper, A.P.; Finger, B.J.; Mattiske, D.M.; Pask, A.J.; Green, M.P. Atrazine induces penis abnormalities including hypospadias in mice. J. Dev. Orig. Health Dis. 2020, 11, 246–249. [Google Scholar] [CrossRef]
- Victor-Costa, A.B.; Bandeira, S.M.C.; Oliveira, A.G.; Mahecha, G.A.B.; Oliveira, C.A. Changes in testicular morphology and steroidogenesis in adult rats exposed to Atrazine. Reprod. Toxicol. 2010, 29, 323–331. [Google Scholar] [CrossRef]
- National Health and Medical Research Council (NHMRC) and National Resource Management Ministerial Council (NRMMC). Australian Drinking Water Guidlines 6 National Water Quality Management Strategy; Australian Government: Canberra, Australia, 2017; pp. 1–1163.
- Environmental Protection Agency. National primary drinking water regulations. United States Environmental Protection Agency. Available online: https://www.epa.gov/ground-water-and-drinking-water/national-primary-drinking-water-regulations (accessed on 12 December 2022).
- Pinter, A.; Torok, G.; Borzsonyi, M.; Surjan, A.; Csik, M.; Kelecsenyi, Z.; Kocsis, Z. Long-term carcinogenicity bioassay of the herbicide atrazine in F344 rats. Neoplasma 1990, 37, 533–544. [Google Scholar] [CrossRef]
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; Flaws, J.A.; Nadal, A.; Prins, G.S.; Toppari, J.; Zoeller, R.T. EDC-2: The Endocrine Society’s second scientific statement on endocrine-disrupting chemicals. Endocr. Rev. 2015, 36, E1–E150. [Google Scholar] [PubMed]
- Xin, F.; Susiarjo, M.; Bartolomei, M.S. Multigenerational and transgenerational effects of endocrine disrupting chemicals: A role for altered epigenetic regulation? Semin. Cell Dev. Biol. 2015, 43, 66–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, C.E.; Myers, J.P.; Vandenberg, L.N. Nonmonotonic dose-response curves occur in dose ranges that are relevant to regulatory decision-making. Dose Response 2018, 16, 1559325818798282. [Google Scholar] [CrossRef] [PubMed]
- Fraites, M.J.; Narotsky, M.G.; Best, D.S.; Stoker, T.E.; Davis, L.K.; Goldman, J.M.; Hotchkiss, M.G.; Klinefelter, G.R.; Karenl, A.; Qian, Y.R.; et al. Gestational atrazine exposure: Effects on male reproductive development and metabolite distribution in the dam, fetus, and neonate. Reprod. Toxicol. 2011, 32, 52–63. [Google Scholar] [CrossRef]
- National Health and Medical Research Council (NHMRC). Australian Code for the Care and Use of Animals for Scientific Purposes, 3rd ed.; Australian Government: Canbera, Australia, 2013; pp. 1–90.
- Deboer, M.D.; Li, Y. Puberty is delayed in male mice with dextran sodium sulfate colitis out of proportion to changes in food intake, body weight, and serum levels of leptin. Pediatr. Res. 2011, 69, 34–39. [Google Scholar] [CrossRef] [Green Version]
- Houston, B.J.; Nagirnaja, N.; Merriner, D.J.; O’Connor, A.E.; Okuda, H.; Omurtag, K.; Smith, S.; Aston, K.I.; Conrad, D.F.; O’Bryan, M.K. The Sertoli cell expressed gene secernin-1 (Scrn1) is dispensable for male fertility in the mouse. Dev. Dyn. 2021, 250, 922–931. [Google Scholar] [CrossRef]
- Evans, J.; Kaitu’u-Lino, T.; Salamonsen, L.A. Extracellular matrix dynamics in scar-free endometrial repair: Perspectives from mouse in vivo and human in vitro studies. Biol. Reprod. 2011, 85, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Tarulli, G.A.; Meachem, S.J.; Schlatt, S.; Stanton, P.G. Regulation of testicular tight junctions by gonadotrophins in the adult Djungarian hamster in vivo. Reproduction 2008, 135, 867–877. [Google Scholar] [CrossRef] [Green Version]
- Tarulli, G.A.; Laven-Law, G.; Shehata, M.; Walters, K.A.; Denis, I.M.; Rahmen, M.M.; Handelsman, D.J.; Dean, N.R.; Tilley, W.D.; Hickey, T.E. Androgen receptor signalling promotes a luminal phenotype in mammary epithelial cells. J. Mammary Gland Biol. Neoplasia 2019, 24, 99–108. [Google Scholar] [CrossRef]
- Tarulli, G.A.; Pask, A.J.; Renfree, M.B. Discrete hedgehog factor expression and action in the developing phallus. Int. J. Mol. Sci. 2020, 21, 1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manna, P.R.; Dyson, M.T.; Stocco, D.M. Regulation of the steroidogenic acute regulatory protein gene expression: Present and future perspectives. Mol. Hum. Reprod. 2009, 15, 321–333. [Google Scholar] [CrossRef] [Green Version]
- Yadav, R.; Petrunak, E.M.; Estrada, D.F.; Scott, E.E. Structural insights into the function of steroidogenic cytochrome P450 17A1. Mol. Cell. Endocrinol. 2017, 441, 68–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, W.L.; Auchus, R.J. The molecular biology, biochemistry, and physiology of human steroidogenesis and its disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manna, P.R.; Stetson, C.L.; Slominski, A.T.; Pruitt, K. Role of the steroidogenic acute regulatory protein in health and disease. Endocrine 2016, 51, 7–21. [Google Scholar] [CrossRef] [Green Version]
- Papoulias, D.M.; Tillitt, D.E.; Talykina, M.G.; Whyte, J.J.; Richter, C.A. Atrazine reduces reproduction in Japanese medaka (Oryzias latipes). Aquat. Toxicol. 2014, 154, 230–239. [Google Scholar] [CrossRef]
- Leet, J.K.; Richter, C.A.; Conman, R.S.; Berninger, J.P.; Bhandari, R.K.; Nicks, D.K.; Zajicek, J.L.; Blazer, V.S.; Tillitt, D.E. Effects of early life stage exposure of largemouth bass to atrazine or a model estrogen (17alpha-ethinylestradiol). PeerJ 2020, 8, e9614. [Google Scholar] [CrossRef]
- Azizi, H.; NiaziTabar, A.; Mohammadi, A.; Skutella, T. Characterization of DDX4 gene expression in human cases with non-obstructive azoospermia and in sterile and fertile mice. J. Reprod. Infertil. 2021, 22, 85–91. [Google Scholar] [CrossRef]
- Tarulli, G.A.; Stanton, P.G.; Meachem, S.J. Is the adult Sertoli cell terminally differentiated? Biol. Reprod. 2012, 87, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Martins-Santos, E.; Pimenta, C.G.; Campos, P.R.N.; Franco, M.B.; Gomes, D.A.; Mahecha, G.A.B.; Oliveira, C.A. Persistent testicular structural and functional alterations after exposure of adult rats to atrazine. Reprod. Toxicol. 2017, 73, 201–213. [Google Scholar] [CrossRef]
- Nadworny, A.S.; Guruju, M.R.; Poor, D.; Doran, R.M.; Sharma, R.V.; Kotlikoff, M.I.; Davisson, R.L. Nox2 and Nox4 influence neonatal c-kit(+) cardiac precursor cell status and differentiation. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H829–H842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebourcet, D.; Monteiro, A.; Cruickshanks, L.; Jeffery, N.; Smith, S.; Milne, L.; O’Shaughnessy, P.J.; Smith, L.B. Relationship of transcriptional markers to Leydig cell number in the mouse testis. PLoS ONE 2019, 14, e0219524. [Google Scholar] [CrossRef] [PubMed] [Green Version]





| Age = 3 Months | Age = 6 Months | |||
|---|---|---|---|---|
| Control (n = 19) | ATZ (n = 15) | Control (n = 17) | ATZ (n = 13) | |
| Body weight (g) | 27.85 ± 0.55 | 29.14 ± 0.55 | 35.17 ± 1.00 | 35.98 ± 1.35 |
| Testes (g) | 0.19 ± 0.01 | 0.19 ± 0.01 | 0.20 ± 0.01 | 0.20 ± 0.01 |
| Seminal vesicle (g) | 0.22 ± 0.01 | 0.25 ± 0.01 | 0.37 ± 0.02 | 0.40 ± 0.02 |
| Liver (g) | 1.18 ± 0.07 | 1.23 ± 0.07 | 1.48 ± 0.12 | 1.48 ± 0.13 |
| Gonadal fat (g) | 0.58 ± 0.05 | 0.68 ± 0.08 | 1.27 ± 0.14 | 1.43 ± 0.20 |
| Age = 3 Months | Age = 6 Months | |||
|---|---|---|---|---|
| Control (n = 12) | ATZ (n = 17) | Control (n = 12) | ATZ (n = 19) | |
| Body weight (g) | 29.09 ± 0.95 | 29.54 ± 0.62 | 34.74 ± 1.81 | 36.19 ± 0.71 |
| Testes (g) | 0.18 ± 0.01 | 0.19 ± 0.01 | 0.18 ± 0.01 | 0.20 ± 0.01 |
| Seminal vesicle (g) | 0.25 ± 0.01 | 0.27 ± 0.01 | 0.39 ± 0.02 | 0.42 ± 0.01 |
| Liver (g) | 1.08 ± 0.10 | 1.22 ± 0.06 | 1.37 ± 0.11 | 1.48 ± 0.06 |
| Gonadal fat (g) | 0.71 ± 0.12 | 0.76 ± 0.07 | 1.41 ± 0.19 | 1.45 ± 0.09 |
| Age = 3 Months | Age = 6 Months | |||
|---|---|---|---|---|
| Control (n = 19) | ATZ (n = 15) | Control (n = 17) | ATZ (n = 13) | |
| BMD (mg/cm2) | 53.15 ± 0.60 | 54.33 ± 0.50 | 58.37 ± 0.86 | 58.09 ± 1.17 |
| BMC (mg) | 480.11 ± 10.94 | 493.87 ± 10.20 | 524.06 ± 9.94 | 520.46 ± 11.03 |
| Fat (%) | 17.06 ± 1.02 | 16.89 ± 1.34 | 25.06 ± 1.95 | 24.06 ± 2.23 |
| Muscle (%) | 82.94 ± 1.02 | 83.11 ± 1.34 | 74.94 ± 1.95 | 75.94 ± 2.23 |
| Age = 3 Months | Age = 6 Months | |||
|---|---|---|---|---|
| Control (n = 14) | ATZ (n = 17) | Control (n = 10) | ATZ (n = 19) | |
| BMD (mg/cm2) | 55.90 ± 1.03 | 56.74 ± 0.63 | 58.84 ± 1.93 | 60.15 ± 0.92 |
| BMC (mg) | 475.43 ± 11.03 | 503.71 ± 9.31 | 507.88 ± 25.37 | 543.00 ± 7.55 |
| Fat (%) | 18.86 ± 1.45 | 18.34 ± 0.95 | 27.06 ± 2.65 | 26.01 ± 1.25 |
| Muscle (%) | 81.14 ± 1.45 | 81.67 ± 0.95 | 72.94 ± 2.65 | 73.99 ± 1.25 |
| F1 | F2 | |||
|---|---|---|---|---|
| Control (n = 7) | ATZ (n = 10) | Control (n = 10) | ATZ (n = 11) | |
| Total motile sperm (%) | 81.43 ± 3.34 | 85.60 ± 2.20 | 84.90 ± 2.09 | 83.18 ± 1.85 |
| Progressive motile sperm (%) | 50.43 ± 4.19 | 55.80 ± 2.80 | 54.70 ± 2.80 | 53.72 ± 2.49 |
| Rapid velocity distribution (%) | 54.86 ± 4.10 | 60.30 ± 2.70 | 59.30 ± 2.90 | 58.27 ± 2.53 |
| Control (n = 4) | ATZ (n = 5) | |
|---|---|---|
| Epithelium height (µm) | 65.85 ± 1.58 | 67.23 ± 1.15 |
| Epithelium diameter (µm) | 184.26 ± 4.82 | 189.30 ± 4.58 |
| Tubule (%) | 90.71 ± 0.74 | 91.34 ± 0.77 |
| Interstitial space (%) | 9.29 ± 0.74 | 8.66 ± 0.77 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolaitis, N.D.; Finger, B.J.; Merriner, D.J.; Nguyen, J.; Houston, B.J.; O’Bryan, M.K.; Stringer, J.M.; Zerafa, N.; Nguyen, N.; Hutt, K.J.; et al. Impact of Chronic Multi-Generational Exposure to an Environmentally Relevant Atrazine Concentration on Testicular Development and Function in Mice. Cells 2023, 12, 648. https://doi.org/10.3390/cells12040648
Kolaitis ND, Finger BJ, Merriner DJ, Nguyen J, Houston BJ, O’Bryan MK, Stringer JM, Zerafa N, Nguyen N, Hutt KJ, et al. Impact of Chronic Multi-Generational Exposure to an Environmentally Relevant Atrazine Concentration on Testicular Development and Function in Mice. Cells. 2023; 12(4):648. https://doi.org/10.3390/cells12040648
Chicago/Turabian StyleKolaitis, Nicola D., Bethany J. Finger, D. Jo Merriner, Joseph Nguyen, Brendan J. Houston, Moira K. O’Bryan, Jessica M. Stringer, Nadeen Zerafa, Ngoc Nguyen, Karla J. Hutt, and et al. 2023. "Impact of Chronic Multi-Generational Exposure to an Environmentally Relevant Atrazine Concentration on Testicular Development and Function in Mice" Cells 12, no. 4: 648. https://doi.org/10.3390/cells12040648