
Risk Assessment of Transgender People: Development of Rodent Models Mimicking Gender-Affirming Hormone Therapies and Identification of Sex-Dimorphic Liver Genes as Novel Biomarkers of Sex Transition
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Experimental Design of the Animal Study
- -
- for the dFM model, n = 4 based on T serum levels reported in Kinnear H.M. et al. [control group: 0.2 ± 0.3 and 0.45 mg of T group: 16 ± 5 ng/mL—mean ± standard deviation (SD)] [11];
- -
- for the dMF model, n = 3 according to Gomez A. et at. that report T serum levels of 1.901 ± 0.413 and 0.043 ± 0.023 ng/mL [mean ± standard error (SE)] in control and E2 plus CPA group (0.2 + 0.8 mg/kw bw day), respectively [12].
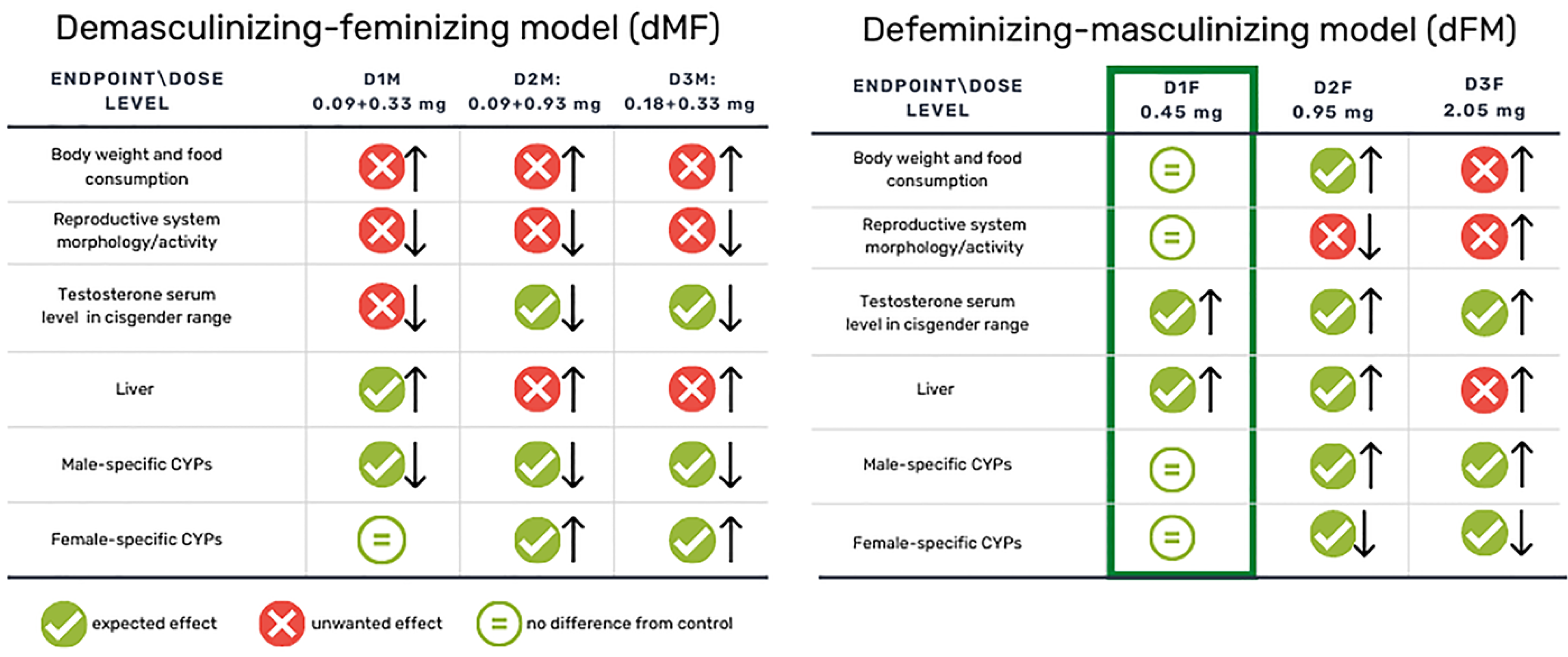
2.2.1. Demasculinizing-Feminizing Model (dMF)
- Control group (CM): sesame oil (vehicle);
- Dose 1 (D1M): 0.09 + 0.33 mg per dose;
- Dose 2 (D2M): 0.09 + 0.93 mg per dose;
- Dose 3 (D3M): 0.18 + 0.33 mg per dose.
2.2.2. Defeminizing-Masculinizing Model (dFM)
- Control group (CF): sesame oil (vehicle);
- Dose 1 (D1F): 0.45 mg per dose;
- Dose 2 (D2F): 0.95 mg per dose;
- Dose 3 (D3F): 2.05 mg per dose.
2.3. Sperm Count
2.4. Blood Collection and Biochemical Evaluation of Hormones
- -
- E2 Rat kit (RTC009R—BioVendor Brno, Czech Republic), LOD 2.5 pg/mL
- -
- T Mouse/Rat kit (RTC001R—BioVendor Brno, Czech Republic), LOD 2.5 pg/mL;
- -
- LH Rat Kit (ELK2367—ELK Biotechnology, Whuan, China), LOD 37.59 pg/mL
2.5. Histological and Histomorphometrical Analysis
2.6. Gene Expression Analysis
2.7. Data Analysis
3. Results
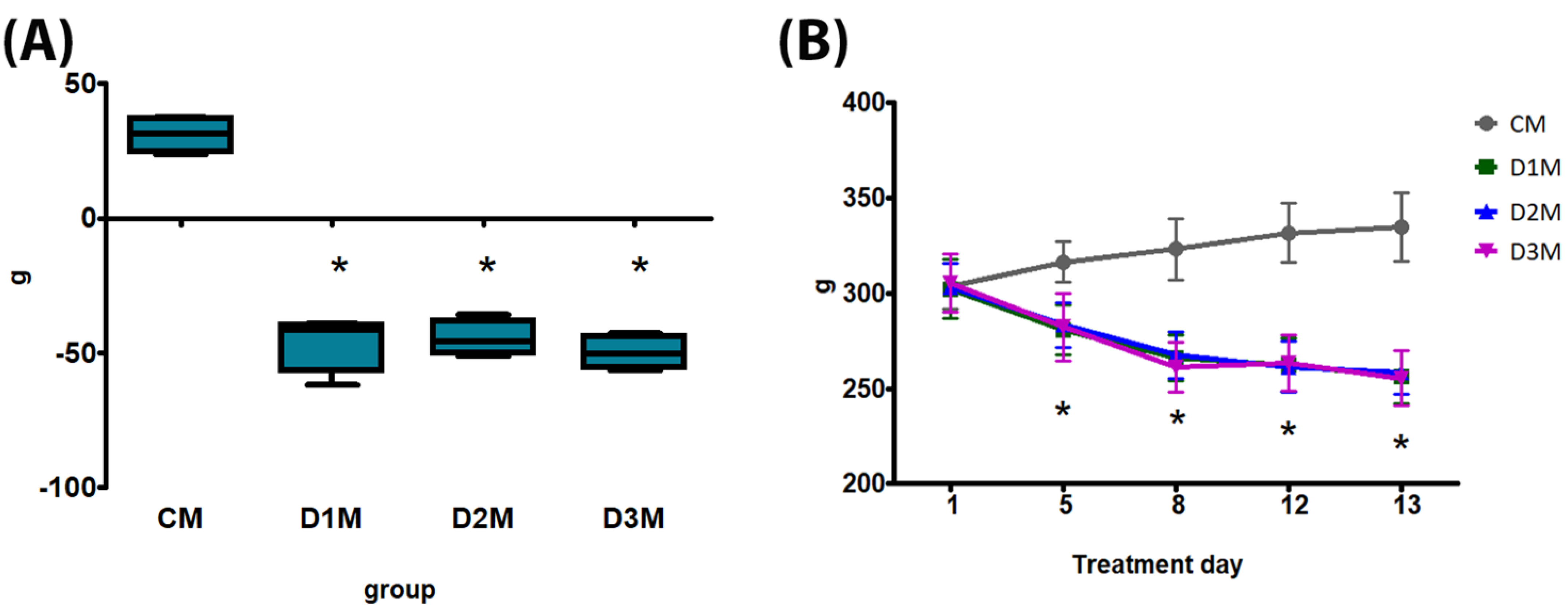
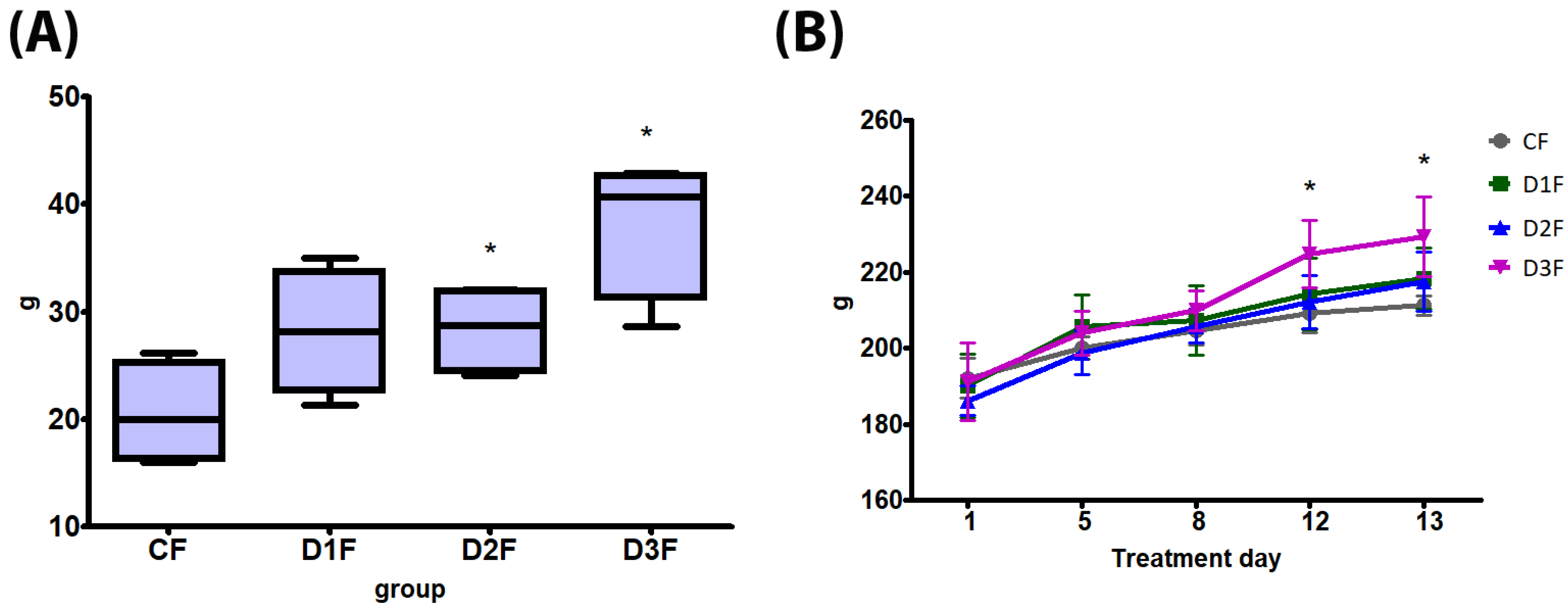
3.1. General Toxicity, Sperm Count and Clitoral Gain
3.1.1. Demasculinizing-Feminizing Model
3.1.2. Defeminizing-Masculinizing Model
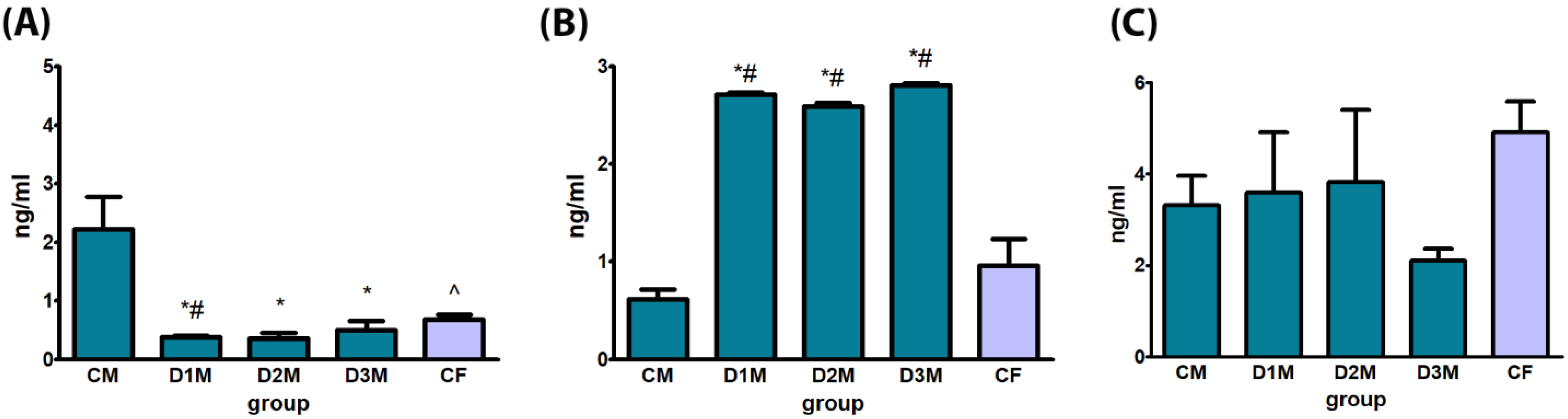
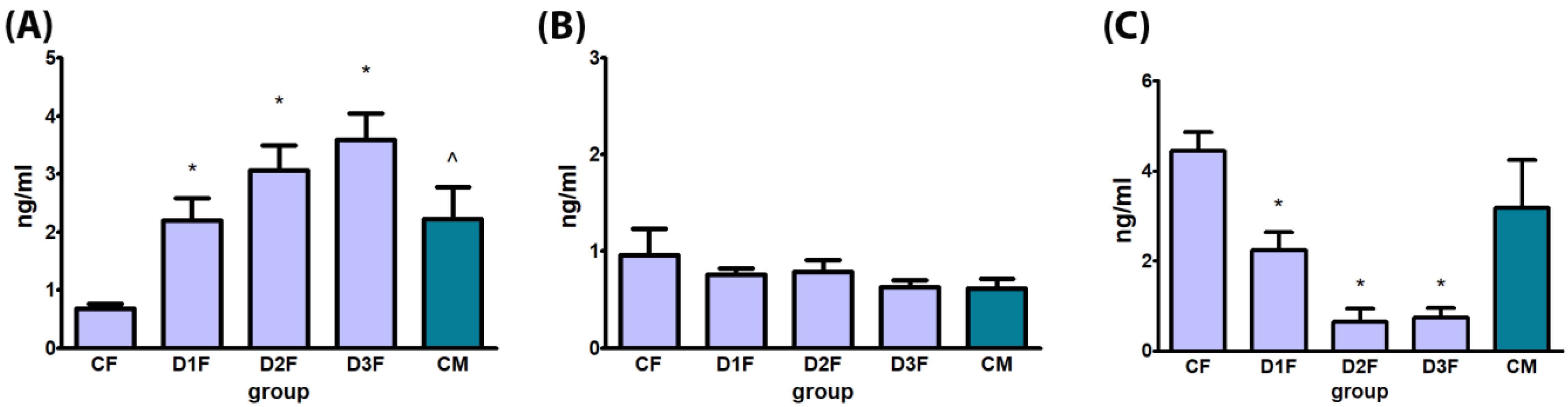
3.2. Biochemical Evaluation of Hormones
3.2.1. Demasculinizing-Feminizing Model
3.2.2. Defeminizing-Masculinizing Model
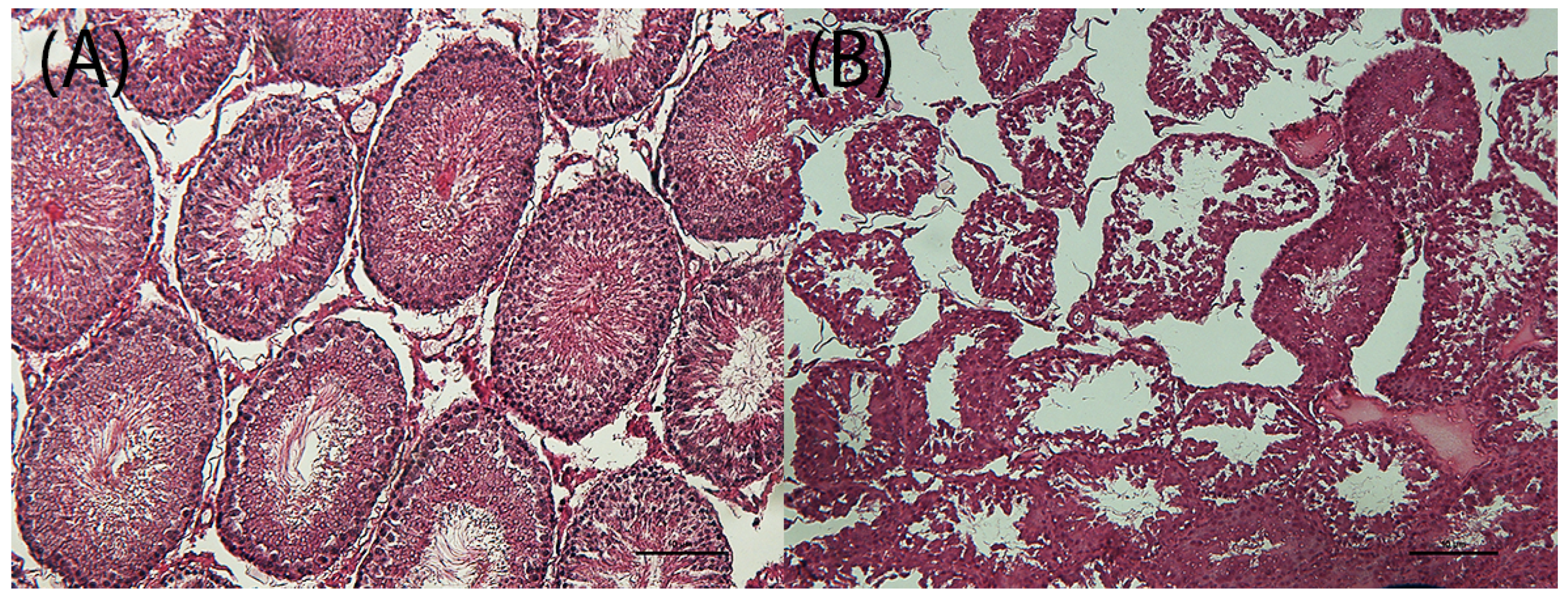
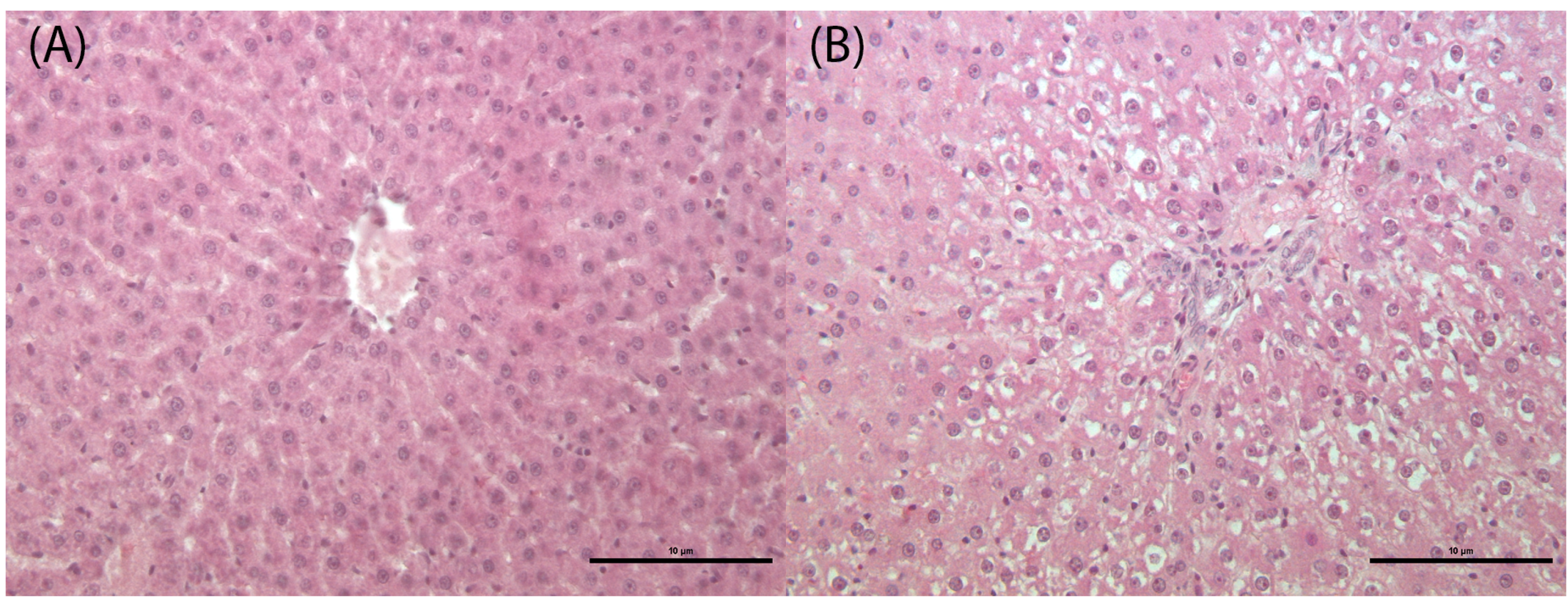
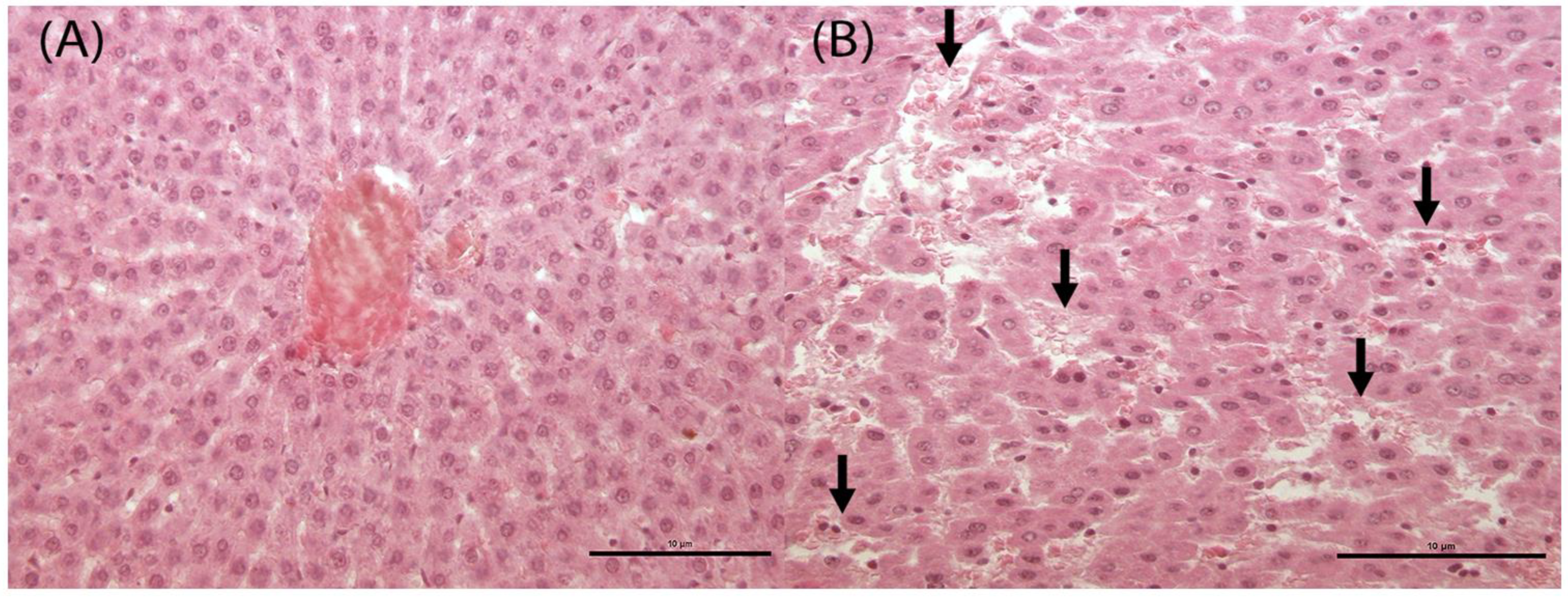
3.3. Histological and Histomorphometrical Analysis
3.3.1. Demasculinizing-Feminizing Model
3.3.2. Defeminizing-Masculinizing Model
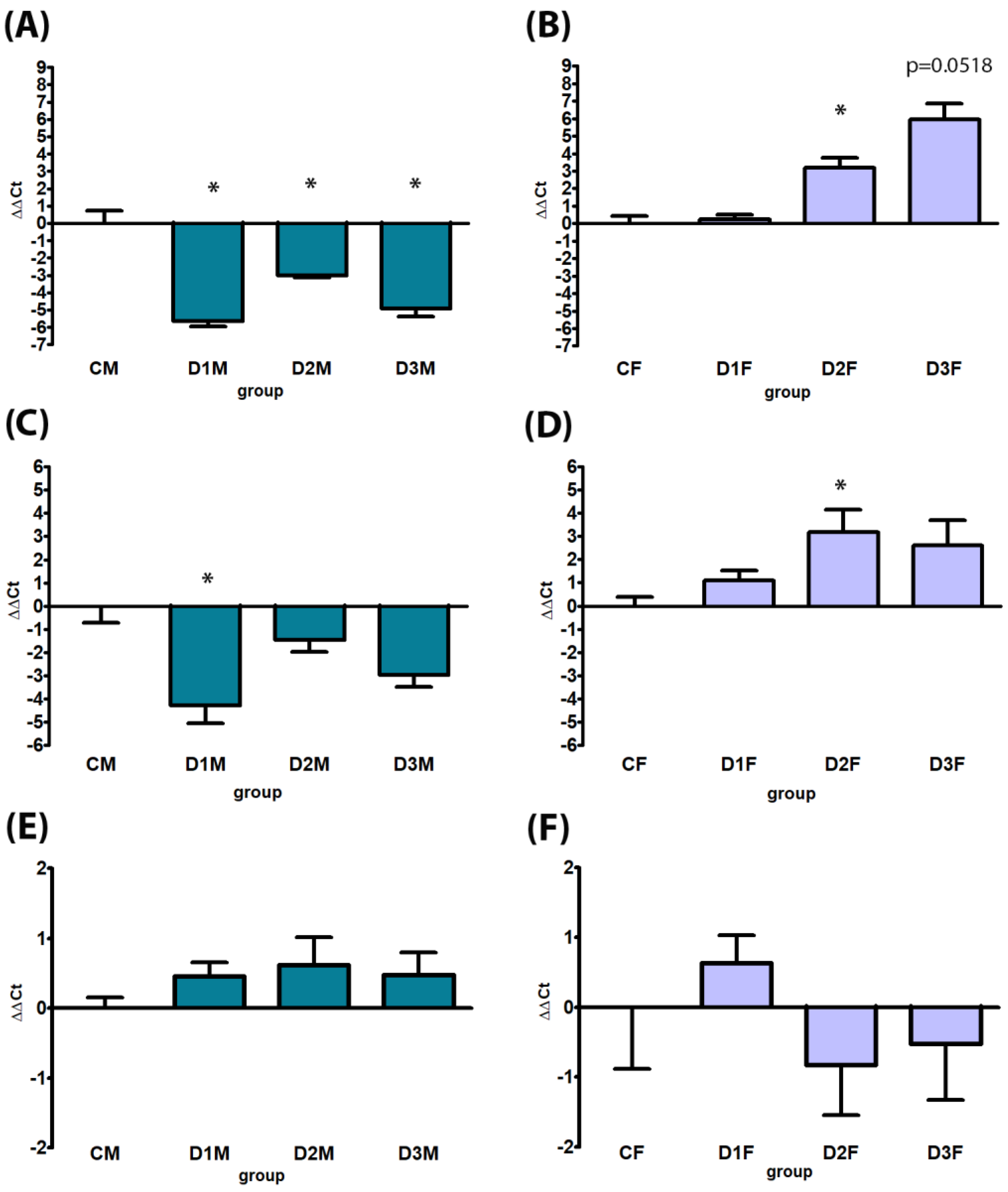
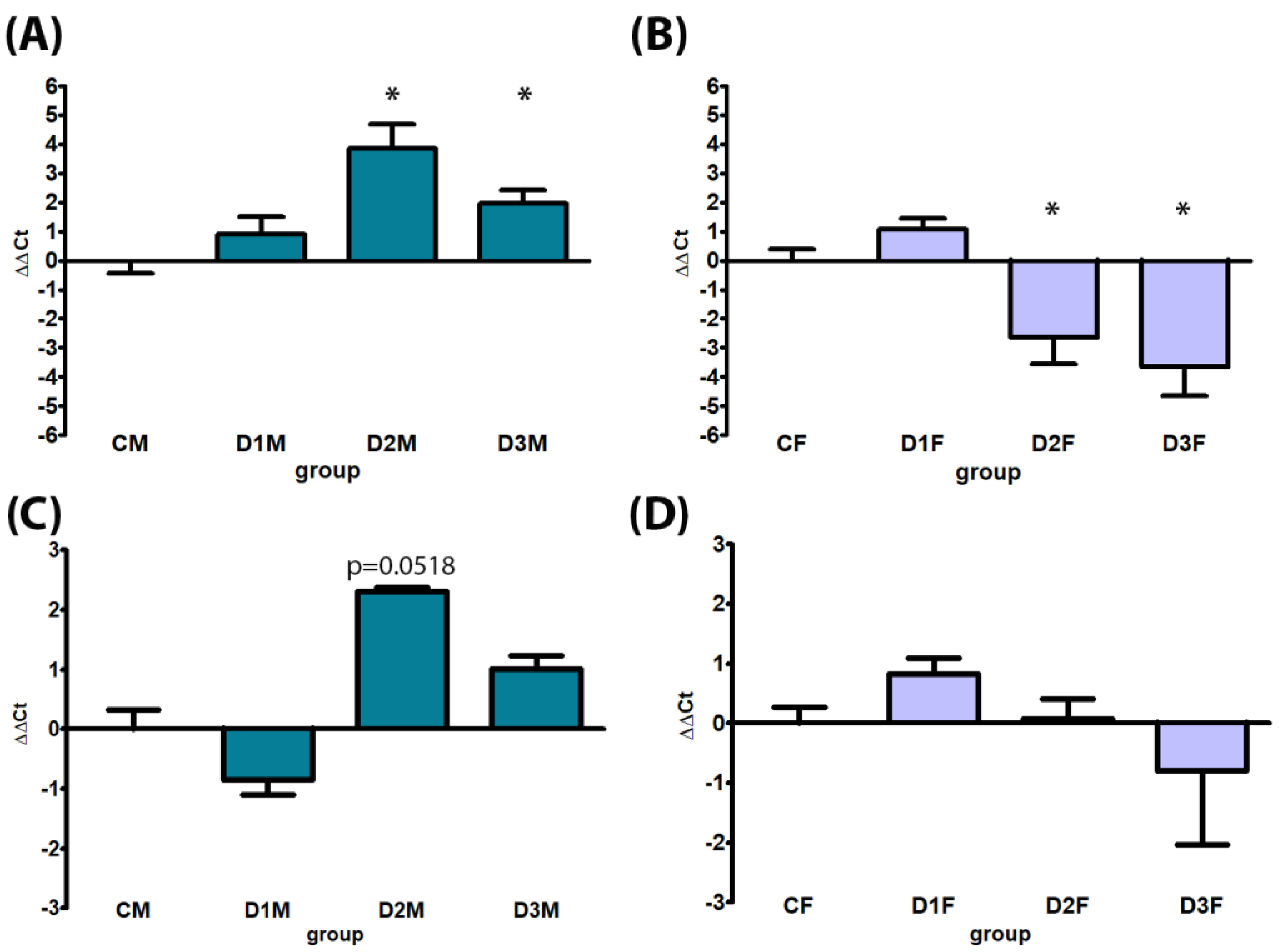
3.4. Gene Expression
3.4.1. Demasculinizing-Feminizing Model
3.4.2. Defeminizing-Masculinizing Model
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cirrincione, L.R.; Huang, K.J. Sex and Gender Differences in Clinical Pharmacology: Implications for Transgender Medicine. Clin. Pharmacol. Ther. 2021, 110, 897–908. [Google Scholar] [CrossRef] [PubMed]
- Mirabella, M.; Piras, I.; Fortunato, A.; Fisher, A.D.; Lingiardi, V.; Mosconi, M.; Ristori, J.; Speranza, A.M.; Giovanardi, G. Gender Identity and Non-Binary Presentations in Adolescents Attending Two Specialized Services in Italy. J. Sex. Med. 2022, 19, 1035–1048. [Google Scholar] [CrossRef] [PubMed]
- Defreyne, J.; Vander Stichele, C.; Iwamoto, S.J.; T’Sjoen, G. Gender-affirming hormonal therapy for transgender and gender-diverse people-A narrative review. Best Pract. Res. Clin. Obstet. Gynaecol. 2022, 102296. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Goodman, M.; Adams, N.; Corneil, T.; Hashemi, L.; Kreukels, B.; Motmans, J.; Snyder, R.; Coleman, E. Epidemiological considerations in transgender health: A systematic review with focus on higher quality data. Int. J. Transgend. Health 2020, 21, 125–137. [Google Scholar] [CrossRef] [PubMed]
- T’Sjoen, G.; Arcelus, J.; Gooren, L.; Klink, D.T.; Tangpricha, V. Endocrinology of Transgender Medicine. Endocr. Rev. 2019, 40, 97–117. [Google Scholar] [CrossRef]
- Pettit, S.D. ToxPoint: Health Disparities, COVID-19, and Owning Our Share. Toxicol. Sci. 2021, 179, 147–148. [Google Scholar] [CrossRef]
- Maggi, A.; Della Torre, S. Sex, metabolism and health. Mol. Metab. 2018, 15, 3–7. [Google Scholar] [CrossRef]
- Robertson, G.R.; Farrell, G.C.; Liddle, C. Sexually Dimorphic Expression of Rat CYP3A9 and CYP3A18 Genes Is Regulated by Growth Hormone. Biochem. Biophys. Res. Commun. 1998, 242, 57–60. [Google Scholar] [CrossRef]
- Waxman, D.J.; Chang, T.K.H. Hormonal Regulation of Liver Cytochrome P450 Enzymes. In Cytochrome P450; De Montellano, P.R.O., Ed.; Springer: Boston, MA, USA, 1995. [Google Scholar]
- Wauthier, V.; Dubois, P.; Verbeeck, R.K.; Calderon, P.B. Induction of CYP2C12 expression in senescent male rats is well correlated to an increase of HNF3beta expression, while the decline of CYP2C11 expression is unlikely due to a decrease of STAT5 activation. Biochem. Pharmacol. 2007, 73, 923–933. [Google Scholar] [CrossRef]
- Kinnear, H.M.; Constance, E.S.; David, A.; Marsh, E.E.; Padmanabhan, V.; Shikanov, A.; Moravek, M.B. A mouse model to investigate the impact of testosterone therapy on reproduction in transgender men. Hum. Reprod. 2019, 34, 2009–2017. [Google Scholar] [CrossRef]
- Gomez, A.; Cerdan, S.; Perez-Laso, C.; Ortega, E.; Pasaro, E.; Fernandez, R.; Gomez-Gil, E.; Mora, M.; Marcos, A.; Del Cerro, M.C.R.; et al. Effects of adult male rat feminization treatments on brain morphology and metabolomic profile. Horm. Behav. 2020, 125, 104839. [Google Scholar] [CrossRef]
- T’Sjoen, G.; Arcelus, J.; De Vries, A.L.C.; Fisher, A.D.; Nieder, T.O.; Özer, M.; Motmans, J. European Society for Sexual Medicine Position Statement “Assessment and Hormonal Management in Adolescent and Adult Trans People, With Attention for Sexual Function and Satisfaction”. J. Sex. Med. 2020, 17, 570–584. [Google Scholar] [CrossRef]
- Tassinari, R.; Cordelli, E.; Eleuteri, P.; Villani, P.; Pacchierotti, F.; Narciso, L.; Tait, S.; Valeri, M.; Martinelli, A.; Di Felice, G.; et al. Effects of sub-chronic oral exposure to pyrogenic synthetic amorphous silica (NM-203) in male and female Sprague-Dawley rats: Focus on reproductive systems. Reprod. Toxicol. 2021, 105, 17–24. [Google Scholar] [CrossRef]
- Shackelford, C.; Long, G.; Wolf, J.; Okerberg, C.; Herbert, R. Qualitative and Quantitative Analysis of Nonneoplastic Lesions in Toxicology Studies. Toxicol. Pathol. 2002, 30, 93–96. [Google Scholar] [CrossRef]
- Maranghi, F.; Tassinari, R.; Lagatta, V.; Moracci, G.; Macrì, C.; Eusepi, A.; Di Virgilio, A.; Scattoni, M.L.; Calamandrei, G. Effects of the food contaminant semicarbazide following oral administration in juvenile Sprague–Dawley rats. Food Chem. Toxicol. 2009, 47, 472–479. [Google Scholar] [CrossRef]
- Fortune, J.E. The early stages of follicular development: Activation of primordial follicles and growth of preantral follicles. Anim. Reprod. Sci. 2003, 78, 135–163. [Google Scholar] [CrossRef]
- Rusyn, I.; Chiu, W.A.; Wright, F.A. Model systems and organisms for addressing inter- and intra-species variability in risk assessment. Regul. Toxicol. Pharmacol. 2022, 132, 105197. [Google Scholar] [CrossRef]
- Narciso, L.; Catone, T.; Aquilina, G.; Attias, L.; De Angelis, I.; Iuliano, M.G.; Tassinari, R.; Mantovani, A.; Maranghi, F. The juvenile toxicity study as a tool for a science-based risk assessment in the children population group. Reprod. Toxicol. 2017, 72, 136–141. [Google Scholar] [CrossRef]
- Pandey, G.; Pandey, S.P.; Sharma, M. Experimental Hepatotoxicity Produced by Ethinyl estradiol. Toxicol. Int. 2011, 18, 160–162. [Google Scholar] [CrossRef]
- Chen, C.S.; Gao, G.L.; Ho, D.R.; Lin, C.Y.; Chou, Y.T.; Chen, S.C.; Huang, M.C.; Kao, W.Y.; Su, J.J. Cyproterone acetate acts as a disruptor of the aryl hydrocarbon receptor. Sci. Rep. 2021, 11, 5457. [Google Scholar] [CrossRef]
- Tucker, M.J.; Jones, D.V. Effects of cyproterone acetate in C57B1/10J mice. Hum. Exp. Toxicol. 1996, 15, 64–66. [Google Scholar] [CrossRef] [PubMed]
- Kuijpers, S.M.E.; Wiepjes, C.M.; Conemans, E.B.; Fisher, A.D.; T’Sjoen, G.; den Heijer, M. Toward a Lowest Effective Dose of Cyproterone Acetate in Trans Women: Results from the ENIGI Study. J. Clin. Endocrinol. Metab. 2021, 106, e3936–e3945. [Google Scholar] [CrossRef] [PubMed]
- Gusmão-Silva, J.V.; Lichtenecker, D.C.K.; Ferreira, L.G.A.; Gois, I.; Argeri, R.; Gomes, G.N.; Dias-da-Silva, M.R. Body, metabolic and renal changes following cross-sex estrogen/progestogen therapy in a rodent model simulating its use by transwomen. J. Endocrinol. Investig. 2022, 45, 1875–1885. [Google Scholar] [CrossRef] [PubMed]
- Iwasa, T.; Matsuzaki, T.; Yano, K.; Yiliyasi, M.; Kuwahara, A.; Matsui, S.; Irahara, M. Effects of chronic testosterone administration on the degree of preference for a high-fat diet and body weight in gonadal-intact and ovariectomized female rats. Behav. Brain Res. 2018, 349, 102–108. [Google Scholar] [CrossRef]
- Camargo, I.C.; Gaspar, A.L.; Frei, F.; Mesquita Sde, F. Effects of androgenic anabolic steroids on the uterus and reproductive parameters of adult female rats. Rev. Bras. Ginecol. Obstet. 2009, 31, 453–460. [Google Scholar] [CrossRef]
- Yang, M.; Li, J.; An, Y.; Zhang, S. Effects of androgen on immunohistochemical localization of androgen receptor and Connexin 43 in mouse ovary. Tissue Cell 2015, 47, 526–532. [Google Scholar] [CrossRef]
- Uchida, K.; Inoue, K.; Hasegawa, Y.; Hakuno, F.; Takahashi, S.I.; Takenaka, A. Endogenous testosterone reduces hepatic lipid accumulation in protein-restricted male rats. Nutrition 2021, 85, 111130. [Google Scholar] [CrossRef]
- Bartels, C.B.; Uliasz, T.F.; Lestz, L.; Mehlmann, L.M. Short-term testosterone use in female mice does not impair fertilizability of eggs: Implications for the fertility care of transgender males. Hum. Reprod. 2021, 36, 189–198. [Google Scholar] [CrossRef]
- Dkhil, M.A.; Al-Quraishy, S.; Abdel-Baki, A.; Ghanjati, F.; Arauzo-Bravo, M.J.; Delic, D.; Wunderlich, F. Epigenetic modifications of gene promoter DNA in the liver of adult female mice masculinized by testosterone. J. Steroid Biochem. Mol. Biol. 2015, 145, 121–130. [Google Scholar] [CrossRef]
- Le, A.; Huang, K.J.; Cirrincione, L.R. Regulation of drug-metabolizing enzymes by sex-related hormones: Clinical implications for transgender medicine. Trends Pharmacol. Sci. 2022, 43, 582–592. [Google Scholar] [CrossRef]
- Gerges, S.H.; El-Kadi, A.O.S. Sexual Dimorphism in the Expression of Cytochrome P450 Enzymes in Rat Heart, Liver, Kidney, Lung, Brain, and Small Intestine. Drug Metab. Dispos. 2022, 51, 81–94. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CM | D1M | D2M | D3M | |
|---|---|---|---|---|
| n | 4 | 4 | 4 | 4 |
| Feed consumption (g, mean ± SD) | 21.04 ± 0.08 | 13.48 ± 0.40 * | 15.69 ± 0.77 * | 14.97 ± 1.22 * |
| Testis absolute weight (g, mean ± SD) | 3.49 ± 0.15 | 2.20 ± 0.50 * | 2.16 ± 0.16 * | 2.21 ± 0.21 * |
| Testis relative weight (mean ± SD) | 1.05 ± 0.06 | 0.86 ± 0.22 | 0.83 ± 0.07 * | 0.86 ± 0.07 * |
| Liver absolute weight (g, mean ± SD) | 12.23 ± 1.17 | 10.03 ± 1.50 | 11.20 ± 0.96 | 10.14 ± 0.66 * |
| Liver relative weight (mean ± SD) | 3.65 ± 0.26 | 3.91 ± 0.37 | 4.33 ± 0.24 * | 3.97 ± 0.24 |
| CF | D1F | D2F | D3F | |
|---|---|---|---|---|
| n | 4 | 4 | 4 | 4 |
| Feed consumption (g, mean ± SD) | 15.8 ± 0.2 | 16.8 ± 1.0 | 15.4 ± 0.5 | 17.9 ± 1.4 * |
| Haemorrhagic ovaries and uteri | 0/4 (0%) # | 0/4 (0%) | 1/4 (20%) | 4/4 (100%) § |
| Clitoral gain (mm, mean ± SD) | −0.12 ± 0.47 | 0.48 ± 0.43 | 0.24 ± 0.20 | 0.39 ± 0.36 |
| Ovary absolute weight (g, mean ± SD) | 0.17 ± 0.02 | 0.14 ± 0.12 | 0.12 ± 0.02 * | 0.10 ± 0.01 * |
| Ovary relative weight (mean ± SD) | 0.08 ± 0.01 | 0.07 ± 0.01 | 0.06 ± 0.01 * | 0.05 ± 0.01 * |
| Uterus absolute weight (g, mean ± SD) | 0.52 ± 0.07 | 0.36 ± 0.02 * | 0.37 ± 0.03 * | 0.41 ± 0.10 |
| Uterus relative weight (mean ± SD) | 0.21 ± 0.04 | 0.17 ± 0.01 | 0.17 ± 0.02 | 0.18 ± 0.05 |
| Liver absolute weight (g, mean ± SD) | 6.94 ± 0.14 | 7.36 ± 0.59 | 7.47 ±0.61 | 7.25 ± 1.45 |
| Liver relative weight (mean ± SD) | 3.25 ± 0.07 | 3.37 ± 0.25 | 3.48 ± 0.20 | 3.16 ± 0.63 |
| ORGAN/Observation | CM | D1M | D2M | D3M | |
|---|---|---|---|---|---|
| N | 4 | 4 | 4 | 4 | |
| TESTIS: tubule degeneration with germinal epithelium degeneration | |||||
| 0 | 4 | ||||
| 2 | 4 | ||||
| 3 | 4 | ||||
| 4 | 4 | ||||
| Total Finding Incidence | 0 | 4 § | 4 § | 4 § | |
| TESTIS: tuble area (μm2; mean ± SD) | 422.3 ± 74.3 | 271.9 ± 147.1 | 264.8 ± 85.8 | 223.0 ± 30.7 * | |
| LIVER: sinusoidal dilatation | |||||
| 0 | 4 | ||||
| 1 | 2 | ||||
| 2 | 2 | 4 | 4 | ||
| Total Finding Incidence | 0 | 4 § | 4 § | 4 § | |
| LIVER: hepatocyte vacuolation | |||||
| 0 | 4 | ||||
| 1 | 4 | ||||
| 2 | 2 | ||||
| 3 | 2 | 4 | |||
| Total Finding Incidence | 0 | 4 § | 4 § | 4 § | |
| ORGAN/Observation | CF | D1F | D2F | D3F | |
|---|---|---|---|---|---|
| N | 4 | 4 | 4 | 4 | |
| UTERUS: endometrial and/or myometrial hyperaemic vessels | |||||
| 0 | 4 | 4 | 3 | ||
| 3 | 1 | 4 | |||
| Total Finding Incidence | 0 ## | 0 | 1 | 4 § | |
| UTERUS: total horn areas (μm2; mean ± SD) | 60,651 ± 12,149 | 33,878 ± 3137 * | 36,023 ± 8422 * | 37,795 ± 11,903 | |
| UTERUS: lumen areas (μm2; mean ± SD) | 3864 ± 3878 | 1355 ± 514 | 1683 ± 1364 | 1032 ± 248 * | |
| UTERUS: myometrium areas (μm2; mean ± SD) | 12,614 ± 1158 | 7660 ± 1396 * | 7269 ± 2350 * | 9464 ± 2730 | |
| UTERUS: endometrium areas (μm2; mean ± SD) | 26,246 ± 3253 | 17,261 ± 1939 * | 17,495 ± 3596 * | 13,519 ± 5230 * | |
| UTERUS: endometrium/myometrium areas (mean ± SD) | 2.09 ± 0.28 | 2.31 ± 0.52 * | 2.52 ± 0.60 * | 1.44 ± 0.33 * | |
| UTERUS: luminal epithelium height (μm; mean ± SD) | 23.03 ± 1.56 | 18.10 ± 3.80 | 17.14 ± 2.23 * | 17.60 ± 1.98 * | |
| OVARY: hyperaemic vessels | |||||
| 0 | 4 | 4 | 2 | ||
| 2 | 2 | 4 | |||
| Total Finding Incidence | 0 ## | 0 | 2 | 4§ | |
| OVARY: follicular density (mean ± SD) | 0.08 ± 0.0 | 0.11 ± 0.1 | 0.14 ± 0.0 | 0.15 ± 0.0 | |
| OVARY: primary + secondary follicles (n; mean ± SD) | 28.5 ± 11.7 | 21.25 ±10.5 | 47.3 ±14.2 | 65 ± 6.8 * | |
| OVARY: Graaf follicles (n; mean ± SD) | 3.5 ±1.3 | 2.5 ± 1.0 | 2.5 ± 1.0 | 1.25 ± 0.5 * | |
| LIVER: hepatocyte vacuolation | |||||
| 0 | 4 | 4 | |||
| 1 | 4 | 4 | |||
| Total Finding Incidence | 4 | 4 | 0 § | 0 § | |
| LIVER: sinusoidal dilatation | |||||
| 0 | 4 | ||||
| 1 | 4 | ||||
| 2 | 3 | ||||
| 3 | 1 | 4 | |||
| Total Finding Incidence | 0 | 4 § | 4 § | 4 § | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tassinari, R.; Tammaro, A.; Lori, G.; Tait, S.; Martinelli, A.; Cancemi, L.; Frassanito, P.; Maranghi, F. Risk Assessment of Transgender People: Development of Rodent Models Mimicking Gender-Affirming Hormone Therapies and Identification of Sex-Dimorphic Liver Genes as Novel Biomarkers of Sex Transition. Cells 2023, 12, 474. https://doi.org/10.3390/cells12030474
Tassinari R, Tammaro A, Lori G, Tait S, Martinelli A, Cancemi L, Frassanito P, Maranghi F. Risk Assessment of Transgender People: Development of Rodent Models Mimicking Gender-Affirming Hormone Therapies and Identification of Sex-Dimorphic Liver Genes as Novel Biomarkers of Sex Transition. Cells. 2023; 12(3):474. https://doi.org/10.3390/cells12030474
Chicago/Turabian StyleTassinari, Roberta, Alessia Tammaro, Gabriele Lori, Sabrina Tait, Andrea Martinelli, Luigia Cancemi, Paolo Frassanito, and Francesca Maranghi. 2023. "Risk Assessment of Transgender People: Development of Rodent Models Mimicking Gender-Affirming Hormone Therapies and Identification of Sex-Dimorphic Liver Genes as Novel Biomarkers of Sex Transition" Cells 12, no. 3: 474. https://doi.org/10.3390/cells12030474