
Genome-Wide Gene Expression Analyses of the AtfA/AtfB-Mediated Menadione Stress Response in Aspergillus nidulans
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Condition
2.2. RNA Sequencing
2.3. Evaluation of the Transcriptome Data
- -
- Both AtfA and AtfB are positively acting transcription factors. Their target genes (AtfA-dependent genes and AtfB-dependent genes) should be downregulated in the double mutant (where both atfA and atfB were deleted).
- -
- There are genes that are AtfA-dependent but not AtfB-dependent (AA genes) and genes that are AtfB-dependent but not AtfA-dependent (BB genes).
- -
- There are genes regulated by both AtfA and AtfB. Some of these genes need both atfA and atfB to reach their normal physiological activity detected in the control (deletion of either gene reduces their activities at least to the level of the double mutant; AB genes). Some of the genes, regulated by both AtfA and AtfB, need either atfA or atfB for their normal activity (deletion of either gene does not reduce their transcriptional activity; A/B genes). Some of them need both atfA and atfB for normal activity and deletion of at least one of the two genes reduces the activity to a level between the control and the double mutant (A-B genes). For clarity, see Figure S1.
3. Results
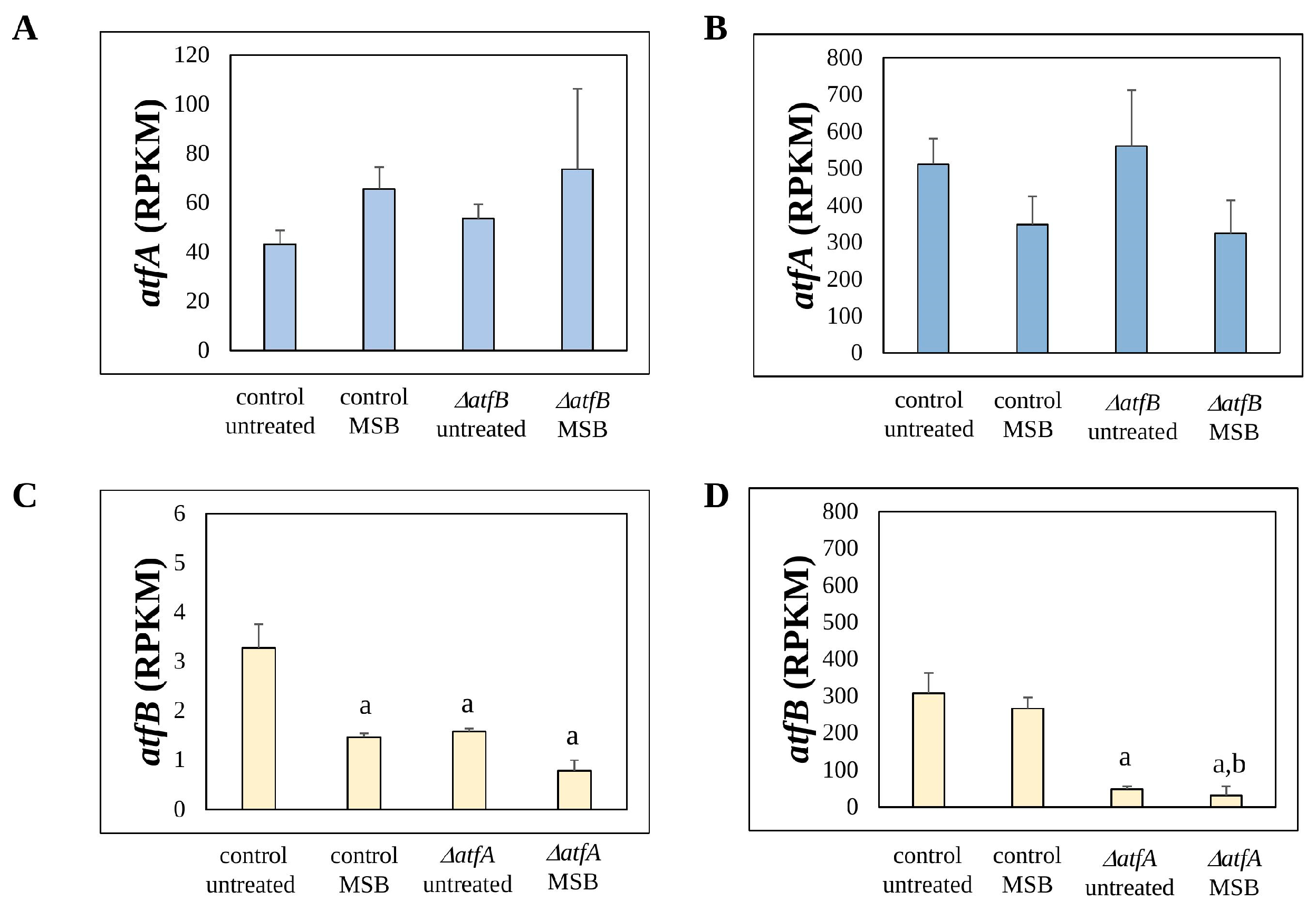
3.1. Deletion of atfA Downregulates atfB
3.2. Most of the AtfB-Dependent Genes Show AtfA-Dependence in Mycelia of Untreated Cultures
3.3. AtfB Regulates Only Few Genes in Mycelia of MSB-Treated Cultures
3.4. AtfB, Similarly to AtfA, Increases Importance in Conidia
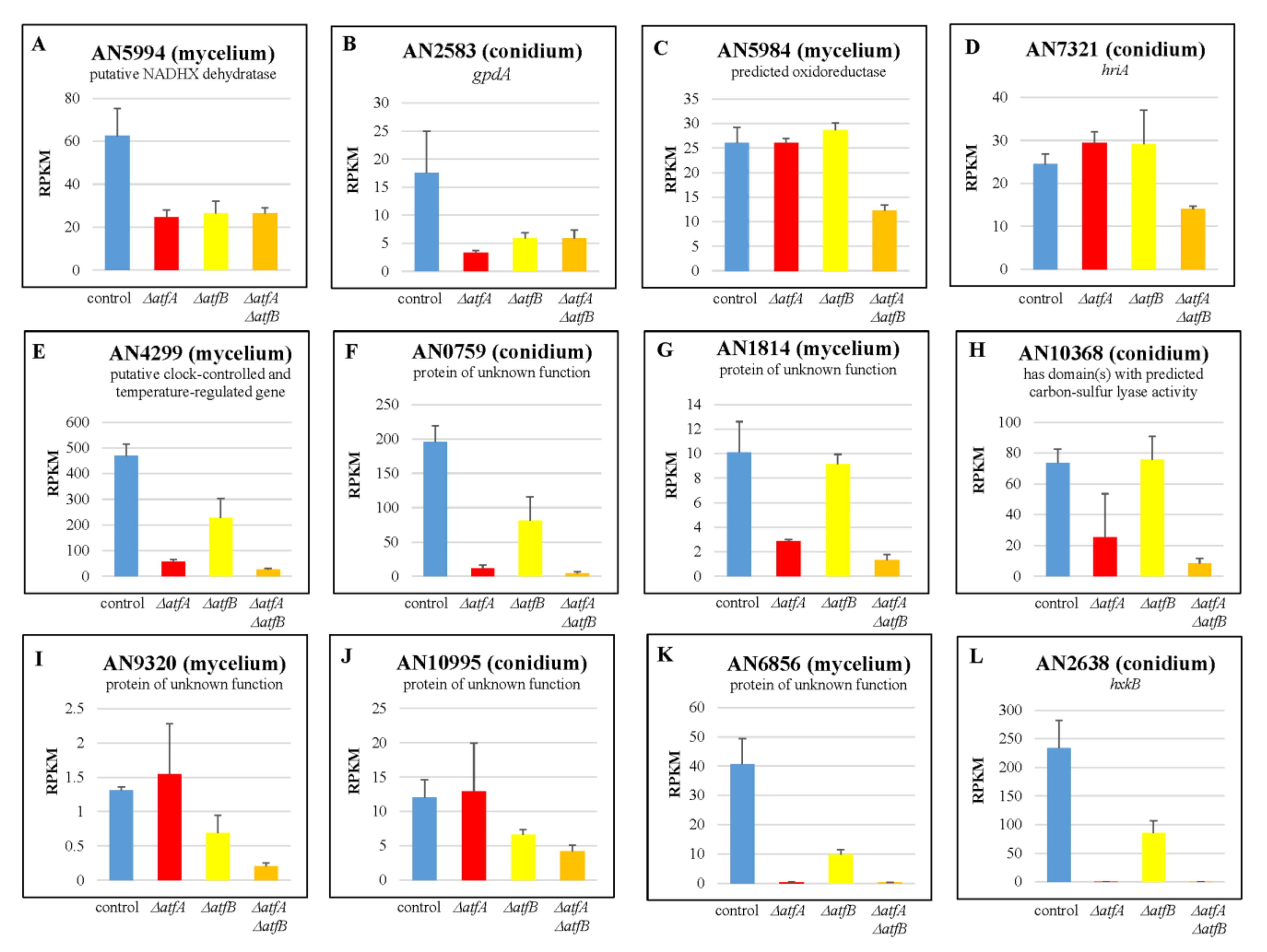
3.5. AtfA Affects Carbohydrate Metabolism and Light Dependent Processes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jindrich, K.; Degnan, B.M. The diversification of the basic leucine zipper family in eukaryotes correlates with the evolution of multicellularity. BMC Evol. Biol. 2016, 16, 28. [Google Scholar] [CrossRef] [Green Version]
- Lara-Rojas, F.; Sanchez, O.; Kawasaki, L.; Aguirre, J. Aspergillus nidulans transcription factor AtfA interacts with the MAPK SakA to regulate general stress responses, development and spore functions. Mol. Microbiol. 2011, 80, 436–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nathues, E.; Joshi, S.; Tenberge, K.B.; von den Driesch, M.; Oeser, B.; Bäumer, N.; Mihlan, M.; Tudzynski, P. CPTF1, a CREB-like transcription factor, is involved in the oxidative stress response in the phytopathogen Claviceps purpurea and modulates ROS level in its host Secale cereale. Mol. Plant Microbe Interact. 2004, 17, 383–393. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, K.; Shiozawa, A.; Watanabe, S.; Fukumori, F.; Kimura, M.; Fujimura, M. ATF-1 transcription factor regulates the expression of ccg-1 and cat-1 genes in response to fludioxonil under OS-2 MAP kinase in Neurospora crassa. Fungal Genet. Biol. 2008, 45, 1562–1569. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Guo, W.; Chen, Y.; Dong, S.; Zhang, X.; Zhang, H.; Song, W.; Wang, W.; Wang, Q.; Lv, R.; et al. The basic leucine zipper transcription factor Moatf1 mediates oxidative stress responses and is necessary for full virulence of the rice blast fungus Magnaporthe oryzae. Mol. Plant Microbe Interact. 2010, 23, 1053–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temme, N.; Oeser, B.; Massaroli, M.; Heller, J.; Simon, A.; Collado, I.G.; Viaud, M.; Tudzynski, P. BcAtf1, a global regulator, controls various differentiation processes and phytotoxin production in Botrytis cinerea. Mol. Plant Pathol. 2012, 13, 704–718. [Google Scholar] [CrossRef]
- Van Nguyen, T.; Kroger, C.; Bonnighausen, J.; Schafer, W.; Bormann, J. The ATF/CREB transcription factor Atf1 is essential for full virulence, deoxynivalenol production, and stress tolerance in the cereal pathogen Fusarium graminearum. Mol. Plant-Microbe Interact. 2013, 26, 1378–1394. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.Z.; Guo, L.J.; Yang, L.Y.; Huang, Y.S. Foatf1, a bZIP transcription factor of Fusarium oxysporum f. sp cubense, is involved in pathogenesis by regulating the oxidative stress responses of Cavendish banana (Musa spp.). Physiol. Mol. Plant Pathol. 2013, 84, 76–85. [Google Scholar] [CrossRef]
- Szabó, Z.; Pákozdi, K.; Murvai, K.; Pusztahelyi, T.; Kecskeméti, Á.; Gáspár, A.; Logrieco, A.F.; Emri, T.; Ádám, A.L.; Leiter, É.; et al. FvatfA regulates growth, stress tolerance as well as mycotoxin and pigment productions in Fusarium verticillioides. Appl. Microbiol. Biotechnol. 2020, 104, 7879–7899. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.P.; Alves de Castro, P.; Reis, T.F.; Paziani, M.H.; Von Zeska Kress, M.R.; Riaño-Pachón, D.M.; Hagiwara, D.; Ries, L.N.; Brown, N.A.; Goldman, G.H. Genome-wide transcriptome analysis of Aspergillus fumigatus exposed to osmotic stress reveals regulators of osmotic and cell wall stresses that are SakA HOG1 and MpkC dependent. Cell. Microbiol. 2016, 19, e12681. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.P.; Horta, M.A.C.; Goldman, G.H. Genetic interactions between Aspergillus fumigatus basic leucine zipper (bZIP) transcription factors AtfA, AtfB, AtfC and AtfD. Front. Fungal Biol. 2021, 2, 632048. [Google Scholar] [CrossRef]
- Hagiwara, D.; Asano, Y.; Yamashino, T.; Mizuno, T. Characterization of bZip-type transcription factor AtfA with reference to stress responses of conidia of Aspergillus nidulans. Biosci. Biotechnol. Biochem. 2008, 72, 2756–2760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagiwara, D.; Asano, Y.; Marui, J.; Yoshimi, A.; Mizuno, T.; Abe, K. Transcriptional profiling for Aspergillus nidulans HogA MAPK signaling pathway in response to fludioxonil and osmotic stress. Fungal Genet. Biol. 2009, 46, 868–878. [Google Scholar] [CrossRef] [PubMed]
- Balázs, A.; Pócsi, I.; Hamari, Z.; Leiter, É.; Emri, T.; Miskei, M.; Oláh, J.; Tóth, V.; Hegedűs, N.; Prade, R.A.; et al. AtfA BZIP-type transcription factor regulates oxidative and osmotic stress responses in Aspergillus Nidulans. Mol. Genet. Genom. 2010, 283, 289–303. [Google Scholar] [CrossRef] [PubMed]
- Leiter, É.; Emri, T.; Pákozdi, K.; Hornok, L.; Pócsi, I. The impact of bZIP Atf1 ortholog global regulators in fungi. Appl. Microbiol. Biotechnol. 2021, 105, 5769–5783. [Google Scholar] [CrossRef]
- Emri, T.; Szarvas, V.; Orosz, E.; Antal, K.; Park, H.; Han, K.H.; Yu, J.-H.; Pócsi, I. Core oxidative stress response in Aspergillus nidulans. BMC Genom. 2015, 16, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orosz, E.; Antal, K.; Gazdag, Z.; Szabó, Z.; Han, K.H.; Yu, J.-H.; Pócsi, I.; Emri, T. Transcriptome-based modeling reveals that oxidative stress induces modulation of the AtfA-dependent signaling networks in Aspergillus nidulans. Int. J. Genom. 2017, 2017, 6923849. [Google Scholar]
- Antal, K.; Gila, B.C.; Pócsi, I.; Emri, T. General stress response or adaptation to rapid growth in Aspergillus nidulans? Fungal Biol. 2020, 124, 376–386. [Google Scholar] [CrossRef]
- Kocsis, B.; Lee, M.-K.; Yu, J.-H.; Nagy, T.; Daróczi, L.; Batta, G.; Pócsi, I.; Leiter, É. Functional analysis of the bZIP-type transcription factors AtfA and AtfB in Aspergillus nidulans. Front. Microbiol. 2022, 13, 1003709. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Arima, T.H.; Iwashita, K.; Yamada, O.; Gomi, K.; Akita, O. Aspergillus oryzae atfB encodes a transcription factor required for stress tolerance in conidia. Fungal Genet. Biol. 2008, 45, 922–932. [Google Scholar] [CrossRef]
- Sakamoto, K.; Iwashita, K.; Yamada, O.; Kobayashi, K.; Mizuno, A.; Akita, O.; Mikami, S.; Shimoi, H.; Gomi, K. Aspergillus oryzae atfA controls conidial germination and stress tolerance. Fungal Genet. Biol. 2009, 46, 887–897. [Google Scholar] [CrossRef]
- Reinke, A.W.; Baek, J.; Ashenberg, O.; Keating, A.E. Networks of bZIP protein-protein interactions diversified over a billion years of evolution. Science 2013, 340, 730–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, T.; Toda, T.; Kominami, K.; Kohnosu, A.; Yanagida, M.; Jones, N. Schizosaccharomyces pombe atf1+ encodes a transcription factor required for sexual development and entry into stationary phase. EMBO J. 1995, 14, 6193–6208. [Google Scholar] [CrossRef] [PubMed]
- Sansó, M.; Gogol, M.; Ayté, J.; Seidel, C.; Hidalgo, E. Transcription factors Pcr1 and Atf1 have distinct roles in stress- and Sty1-dependent gene regulation. Eukaryot. Cell 2008, 7, 826–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eshaghi, M.; Lee, J.H.; Zhu, L.; Poon, S.Y.; Li, J.; Cho, K.H.; Chu, Z.; Karuturi, R.K.; Liu, J. Genomic binding profiling of the fission yeast stress-activated MAPK Sty1 and the bZIP transcriptional activator Atf1 in response to H2O2. PloS ONE 2010, 5, e11620. [Google Scholar] [CrossRef]
- Vo, T.V.; Das, J.; Meyer, M.J.; Cordero, N.A.; Akturk, N.; Wei, X.; Fair, B.J.; Degatano, A.G.; Fragoza, R.; Liu, L.G.; et al. A proteome-wide fission yeast interactome reveals network evolution principles from yeasts to human. Cell 2016, 164, 310–323. [Google Scholar] [CrossRef] [Green Version]
- Barratt, R.W.; Johnson, G.B.; Ogata, W.N. Wild-type and mutant stocks of Aspergillus nidulans. Genetics 1965, 52, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P. A reagent for the single-step simultaneous isolation of RNA, DNA and proteins from cell and tissue samples. BioTechniques 1993, 15, 532–534, 536–537. [Google Scholar] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, U.; Gruben, B.S.; Madrid, S.; Mulder, H.; Nikolaev, I.; de Vries, R.P. Unique regulatory mechanism for D-galactose utilization in Aspergillus nidulans. Appl. Environ. Microbiol. 2011, 77, 7084–7087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fekete, E.; Karaffa, L.; Seiboth, B.; Fekete, E.; Kubicek, C.P.; Flipphi, M. Identification of a permease gene involved in lactose utilisation in Aspergillus nidulans. Fungal Genet. Biol. 2012, 49, 415–425. [Google Scholar] [CrossRef]
- Fekete, E.; Orosz, A.; Kulcsár, L.; Kavalecz, N.; Flipphi, M.; Karaffa, L. Characterization of a second physiologically relevant lactose permease gene (lacpB) in Aspergillus nidulans. Microbiology 2016, 162, 837–847. [Google Scholar] [CrossRef] [PubMed]
- Gila, B.C.; Antal, K.; Birkó, Z.; Keserű, J.S.; Pócsi, I.; Emri, T. Strategies shaping the transcription of carbohydrate-active enzyme genes in Aspergillus nidulans. J. Fungi 2022, 8, 79. [Google Scholar] [CrossRef]
- Gila, B.C.; Moon, H.; Antal, K.; Hajdu, M.; Kovács, R.; Jónás, A.P.; Pusztahelyi, T.; Yu, J.-H.; Pócsi, I.; Emri, T. The DUG pathway governs degradation of intracellular glutathione in Aspergillus nidulans. Appl. Environ. Microbiol. 2021, 87. [Google Scholar] [CrossRef]
- Flipphi, M.; Sun, J.; Robellet, X.; Karaffa, L.; Fekete, E.; Zeng, A.-P.; Kubicek, C.P. Biodiversity and evolution of primary carbon metabolism in Aspergillus nidulans and other Aspergillus spp. Fungal Genet. Biol. 2009, 46, S19–S44. [Google Scholar] [CrossRef]
- Inglis, D.O.; Binkley, J.; Skrzypek, M.S.; Arnaud, M.B.; Cerqueira, G.C.; Shah, P.; Wymore, F.; Wortman, J.R.; Sherlock, G. Comprehensive annotation of secondary metabolite biosynthetic genes and gene clusters of Aspergillus nidulans, A. fumigatus, A. niger and A. oryzae. BMC Microbiol. 2013, 13, 91. [Google Scholar]
- Roze, L.V.; Chanda, A.; Wee, J.; Awad, D.; Linz, J.E. Stress-related transcription factor AtfB integrates secondary metabolism with oxidative stress response in aspergilli. J. Biol. Chem. 2011, 286, 35137–35148. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.B.; Amaike, S.; Wohlbach, D.J.; Gasch, A.P.; Chiang, Y.M.; Wang, C.C.; Bok, J.W.; Rohlfs, M.; Keller, N.P. An Aspergillus nidulans bZIP response pathway hardwired for defensive secondary metabolism operates through aflR. Mol. Microbiol. 2012, 83, 1024–1034. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.B.; Reinke, A.W.; Szilágyi, M.; Emri, T.; Chiang, Y.M.; Keating, A.E.; Pócsi, I.; Wang, C.C.; Keller, N.P. bZIP transcription factors affecting secondary metabolism, sexual development and stress responses in Aspergillus nidulans. Microbiology 2013, 159, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Bákány, B.; Yin, W.-B.; Dienes, B.; Nagy, T.; Leiter, É.; Emri, T.; Keller, N.P.; Pócsi, I. Study on the bZIP-type transcription factors NapA and RsmA in the regulation of intracellular reactive species levels and sterigmatocystin production of Aspergillus nidulans. Int. J. Mol. Sci. 2021, 22, 1577. [Google Scholar] [CrossRef]
- Hagiwara, D.; Takahashi, H.; Kusuya, Y.; Kawamoto, S.; Kamei, K.; Gonoi, T. Comparative transcriptome analysis revealing dormant conidia and germination associated genes in Aspergillus species: An essential role for AtfA in conidial dormancy. BMC Genomics. 2016, 17, 358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Leeuwen, M.R.; Krijgsheld, P.; Bleichrodt, R.; Menke, H.; Stam, H.; Stark, J.; Wösten, H.A.; Dijksterhuis, J. Germination of conidia of Aspergillus niger is accompanied by major changes in RNA profiles. Stud. Mycol. 2013, 74, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, D.; Suzuki, S.; Kamei, K.; Gonoi, T.; Kawamoto, S. The role of AtfA and HOG MAPK pathway in stress tolerance in conidia of Aspergillus fumigatus. Fungal Genet. Biol. 2014, 73, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Armant, O.; Fischer, R. Fungi use the SakA (HogA) pathway for phytochrome-dependent light signalling. Nat. Microbiol. 2016, 1, 16019. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Fischer, R. Light sensing and responses in fungi. Nat. Rev. Microbiol. 2019, 17, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Bayram, O.; Biesemann, C.; Krappmann, S.; Galland, P.; Braus, G.H. More than a repair enzyme: Aspergillus nidulans photolyase-like CryA is a regulator of sexual development. Mol. Biol. Cell 2008, 19, 3254–3262. [Google Scholar] [CrossRef] [Green Version]
- Hallsworth, J.E.; Magan, N. Improved biological control by changing polyols/trehalose in conidia of entomopathogens. Bright Crop Prot. Conf. Pests Dis. 1994, 8D, 1091–1096. [Google Scholar]
- Rangel, D.E.N.; ·Braga, G.U.L.; Fernandes, É.K.K.; Keyser, C.A.; Hallsworth, J.E.; Roberts, D.W. Stress tolerance and virulence of insect-pathogenic fungi are determined by environmental conditions during conidial formation. Curr. Genet. 2015, 61, 383–404. [Google Scholar] [CrossRef]
- Hagiwara, D.; Sakai, K.; Suzuki, S.; Umemura, M.; Nogawa, T.; Kato, N.; Osada, H.; Watanabe, A.; Kawamoto, S.; Gonoi, T.; et al. Temperature during conidiation affects stress tolerance, pigmentation, and trypacidin accumulation in the conidia of the airborne pathogen Aspergillus fumigatus. PLoS ONE 2017, 12, e0177050. [Google Scholar] [CrossRef] [PubMed]
- de Vries, R.P.; Riley, R.; Wiebenga, A.; Aguilar-Osorio, G.; Amillis, S.; Uchima, C.A.; Anderluh, G.; Asadollahi, M.; Askin, M.; Barry, K.; et al. Comparative genomics reveals high biological diversity and specific adaptations in the industrially and medically important fungal genus Aspergillus. Genome Biol. 2017, 18, 28. [Google Scholar] [PubMed] [Green Version]
- Hagiwara, D.; Sakamoto, K.; Abe, K.; Gomi, K. Signaling pathways for stress responses and adaptation in Aspergillus species: Stress biology in the post-genomic era. Biosci. Biotechnol. Biochem. 2016, 80, 1667–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belaish, R.; Sharon, H.; Levdansky, E.; Greenstein, S.; Shadkchan, Y.; Osherov, N. The Aspergillus nidulans cetA and calA genes are involved in conidial germination and cell wall morphogenesis. Fungal Genet. Biol. 2008, 45, 232–242. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene Set | Mycelium from Untreated Cultures | Mycelium from MSB-Treated Cultures | Overlap between MSB-Treated and Untreated Cultures |
|---|---|---|---|
| Responsive to atfA gene deletion (Set1 and Set1+) | 326 upregulated genes 865 downregulated genes | 255 upregulated genes 583 downregulated genes | 85 upregulated genes 336 downregulated genes |
| Responsive to atfB gene deletion (Set2 and Set2+) | 213 upregulated genes 159 downregulated genes | 201 upregulated genes 77 downregulated genes | 71 upregulated genes 12 downregulated genes |
| Responsive to atfA atfB double-gene deletion (Set3 and Set3+) | 200 upregulated genes 457 downregulated genes | 131 upregulated genes 296 downregulated genes | 45 upregulated genes 142 downregulated genes |
| AtfA-dependent genes a (Set26) | 329 genes (236 AA, 16 AB, 40 A/B, 1 A-B1, 3 A-B2 and 33 A-B4 genes) (10 upregulated and 218 b downregulated MSB stress-responsive genes) | 240 genes (232 AA, 2 AB, 4 A/B and 2 A-B3 genes) (17 upregulated and 143 b downregulated MSB stress-responsive genes) | 110 genes |
| AtfB-dependent genes (Set27) | 96 genes (3 BB, 16 AB, 40 A/B, 1 A-B1, 3 A-B2 and 33 A-B4 genes) (2 upregulated and 68 b downregulated MSB stress-responsive genes) | 9 genes (1 BB, 2 AB, 4 A/B and 2 A-B3 genes) (1 upregulated and 3 downregulated MSB stress-responsive genes) | 3 genes |
| Gene Set a | Conidium from Untreated Cultures | Conidium from MSB-Treated Cultures | Overlap between MSB-Treated and Untreated Cultures |
|---|---|---|---|
| Responsive to atfA gene deletion (Set1 and Set1+) | 1875 upregulated genes (82; 25% c) 2116 downregulated genes (326; 38%) | 1386 upregulated genes (57; 22%) 1480 downregulated genes (167; 29%) | 902 upregulated genes 1274 downregulated genes |
| Responsive to atfB gene deletion (Set2 and Set2+) | 117 upregulated genes (5; 2%) 396 downregulated genes (36; 23%) | 74 upregulated genes (7; 3%) 161 downregulated genes (1; 1%) | 30 upregulated genes 108 downregulated genes |
| Responsive to atfA atfB double-gene deletion (Set3 and Set3+) | 1604 upregulated genes (32; 16%) 2018 downregulated genes (248; 54%) | 1553 upregulated genes (27; 21%) 1547 downregulated genes (125; 42%) | 848 upregulated genes 1374 downregulated genes |
| AtfA-dependent genes (Set26) | 1496 genes (185; 56%) (1079 AA, 80 AB, 84 A/B, 13 A-B1, 75 A-B2, 4 A-B3 and 161 A-B4 genes) (6 upregulated and 319b downregulated MSB stress-responsive genes) | 1143 genes (105; 44%) (989 AA, 41 AB, 8 A/B, 2 A-B1, 12 A-B2 and 91 A-B4 genes) (21 upregulated and 154 b downregulated MSB stress-responsive genes) | 1043 genes |
| AtfB-dependent genes (Set27) | 439 genes (26; 27%) (22 BB, 80 AB, 84 A/B, 13 A-B1, 75 A-B2, 4 A-B3 and 161 A-B4 genes) (1 upregulated and 178 b downregulated MSB stress-responsive genes) | 155 genes (1; 11%) (1 BB, 41 AB, 8 A/B, 2 A-B1, 12 A-B2 and 91 A-B4 genes) (5 upregulated and 22 downregulated MSB stress-responsive genes) | 114 genes |
| Culture | AtfA-Dependent Genes (Set26) | AtfB-Dependent Genes (Set27) |
|---|---|---|
| Mycelium (untreated) | C-compound and carbohydrate metabolism; galactose metabolic process | |
| Mycelium (MSB-treated) | Amine/polyamine transport | |
| Conidium (untreated) | C-compound and carbohydrate metabolism; C-compound and carbohydrate transport; glycolysis and gluconeogenesis; pentose phosphate pathway; fructose and mannose metabolism; pyruvate metabolism; glyoxylate and dicarboxylate metabolism; homeostasis of phosphate; proton-driven antiporter; sodium-driven symporter; biosynthesis of secondary metabolites; cellular sensing and response to external stimulus; oxidative stress response | C-compound and carbohydrate metabolism; C-compound and carbohydrate transport; glycolysis and gluconeogenesis; starch and sucrose metabolism; valine, leucine and isoleucine degradation; biosynthesis of secondary metabolites; |
| Conidium (MSB-treated) | C-compound and carbohydrate metabolism; glycolysis and gluconeogenesis; pentose phosphate pathway; fructose and mannose metabolism; glyoxylate and dicarboxylate metabolism; TCA cycle; homeostasis of phosphate; biosynthesis of secondary metabolites; cellular sensing and response to external stimulus; peroxisome |
| Culture | AtfA-Dependent Genes (Set26) | AtfB-Dependent Genes (Set27) |
|---|---|---|
| Mycelium (untreated) | Secondary metabolism: No PKS/NRPS backbone cluster 1, Microperfuranone cluster, AN2924 cluster; AN9005 cluster; AN10297 cluster; Emericellamide cluster | Secondary metabolism: AN2924 cluster; AN10297 cluster; Emericellamide (eas) cluster |
| Mycelium (MSB-treated) | CAZyme genes Phosphorelay response regulator genes Secondary metabolism: Aspercryptin cluster, AN2924 cluster; AN10297 cluster; Emericellamide (eas) cluster | |
| Conidium (untreated) | Glycolysis; Pentose-phosphate shunt; Leloir pathway Antioxidative enzyme genes; iron-sulfur cluster assembly Phosphorelay response regulator genes Secondary metabolism: AN9005 cluster; AN1594 cluster; AN10297 cluster; AN1242 cluster | Transcription factors Secondary metabolism: AN9005 cluster; AN10297 cluster |
| Conidium (MSB-treated) | Glycolysis; pentose-phosphate shunt; Leloir pathway; TCA cycle; respiration Iron-sulfur cluster assembly Phosphorelay response regulator genes Secondary metabolism: AN1594 cluster; AN10297 cluster; AN1242 cluster |
| Gene ID | Gene Name | Description | AtfA/AtfB-Dependence in | |||
|---|---|---|---|---|---|---|
| Mycelium (Untreated) | Mycelium (MSB-Treated) | Conidium (Untreated) | Conidium (MSB-Treated) | |||
| Light dependent regulation | ||||||
| AN0387 | cryA | senses UVA and blue light | AA | AA | AA | AA |
| AN5056 | induced by light | A-B | AA | AA | AA | |
| AN9285 | ccgA | induced by light | AA | AA | AA | A-B |
| AN4299 | induced by light | A-B | AA | AA | AA | |
| AN8638 | cetJ | induced by light | AA | AA | AA | AA |
| AN0045 | induced by light | A-B | AA | AA | AA | |
| AN0693 | induced by light | AA | AA | AA | AA | |
| AN5004 | induced by light | A-B | AA | A-B | AA | |
| AN8339 | induced by light | AA | AA | AA | AA | |
| AN8641 | induced by light | A-B | AA | A-B | AA | |
| Carbohydrate metabolism | ||||||
| AN8138 | aglC | α-galactosidase | A-B | AA | A-B | AA |
| AN2835 | predicted D-arabinono-1,4-lactone oxidase activity | AA | AA | A-B | AA | |
| AN8639 | putative α,α-trehalose-phosphate synthase | AA | AA | A-B | A-B | |
| AN10060 | putative α-amylase | AA | AA | AA | AA | |
| AN3200 | putative β-glucuronidase | AA | AA | AA | AA | |
| AN9180 | putative transketolase | AA | AA | AA | AA | |
| Other | ||||||
| AN2470 | cellular response to nitrosative stress | AA | AA | AA | AA | |
| AN8637 | catA | conidia-specific catalase | AA | AA | AA | AA |
| AN2581 | hk-8-1 | putative histidine-containing phosphotransfer protein | AA | AA | AA | AA |
| AN7945 | hk2 | putative histidine-containing phosphotransfer protein | AA | AA | AA | AA |
| AN9005 | putative polyketide synthase | AA | AA | AB | AB | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kocsis, B.; Lee, M.-K.; Antal, K.; Yu, J.-H.; Pócsi, I.; Leiter, É.; Emri, T. Genome-Wide Gene Expression Analyses of the AtfA/AtfB-Mediated Menadione Stress Response in Aspergillus nidulans. Cells 2023, 12, 463. https://doi.org/10.3390/cells12030463
Kocsis B, Lee M-K, Antal K, Yu J-H, Pócsi I, Leiter É, Emri T. Genome-Wide Gene Expression Analyses of the AtfA/AtfB-Mediated Menadione Stress Response in Aspergillus nidulans. Cells. 2023; 12(3):463. https://doi.org/10.3390/cells12030463
Chicago/Turabian StyleKocsis, Beatrix, Mi-Kyung Lee, Károly Antal, Jae-Hyuk Yu, István Pócsi, Éva Leiter, and Tamás Emri. 2023. "Genome-Wide Gene Expression Analyses of the AtfA/AtfB-Mediated Menadione Stress Response in Aspergillus nidulans" Cells 12, no. 3: 463. https://doi.org/10.3390/cells12030463