
Acute Stress Regulates Sex-Related Molecular Responses in the Human Jejunal Mucosa: Implications for Irritable Bowel Syndrome
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Cold Pain Stress (CPS)
2.3. Jejunal Biopsy
2.4. Systemic Response to CPS
2.4.1. Hand Pain Perception
2.4.2. Autonomic Response
2.4.3. Psychological Response
2.4.4. Hormonal Response
2.5. Intestinal Response to CPS
2.5.1. Molecular Response
2.5.2. Mucosal Inflammation and NR3C1 Expression
2.6. Experimental Design and Procedures (Figure 1)
2.7. Statistical Analysis
3. Results
3.1. Demographics, Baseline Stress, and Depression Levels
3.2. Systemic Response to CPS
3.2.1. Autonomic Response
3.2.2. Psychological Response
3.2.3. Hand Pain Perception
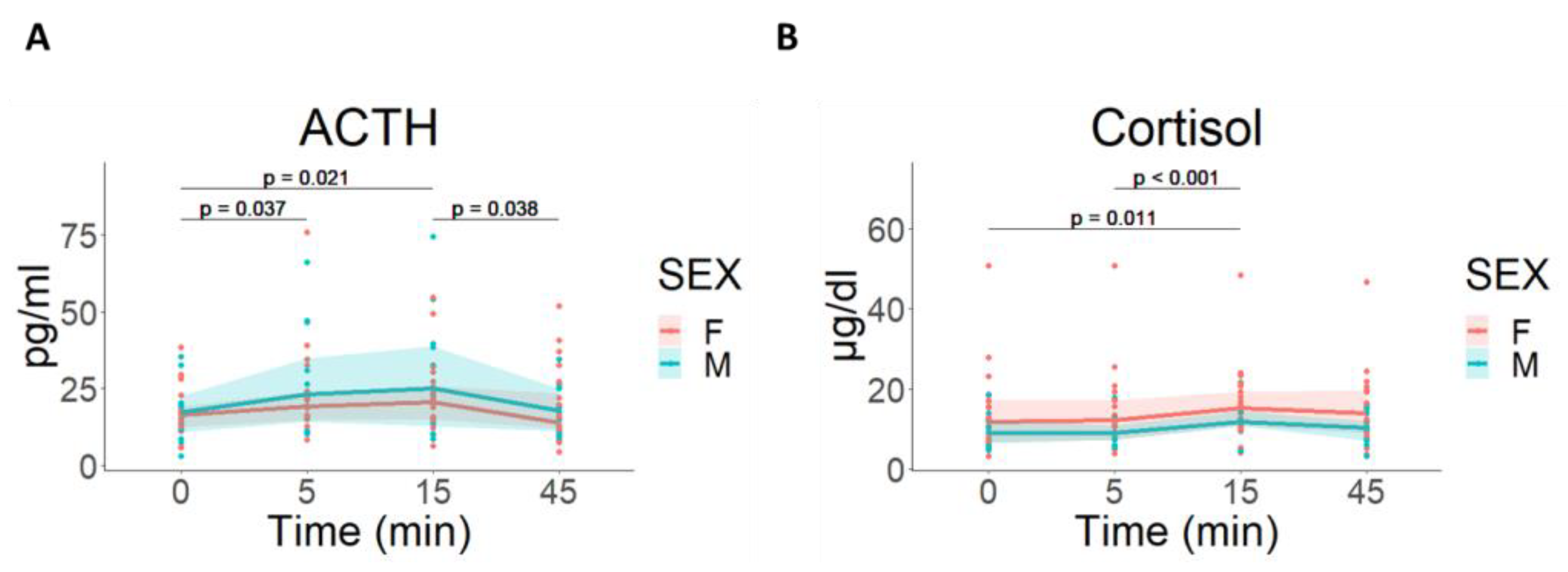
3.2.4. Hormonal Response
3.3. Intestinal Response to CPS
3.3.1. Mucosal Gene Expression
- (a)
- Gene expression levels in the jejunal mucosa at baseline
- (b) Gene expression levels in the jejunal mucosa after CPS
3.3.2. Mucosal Inflammation and NR3C1 Protein Expression
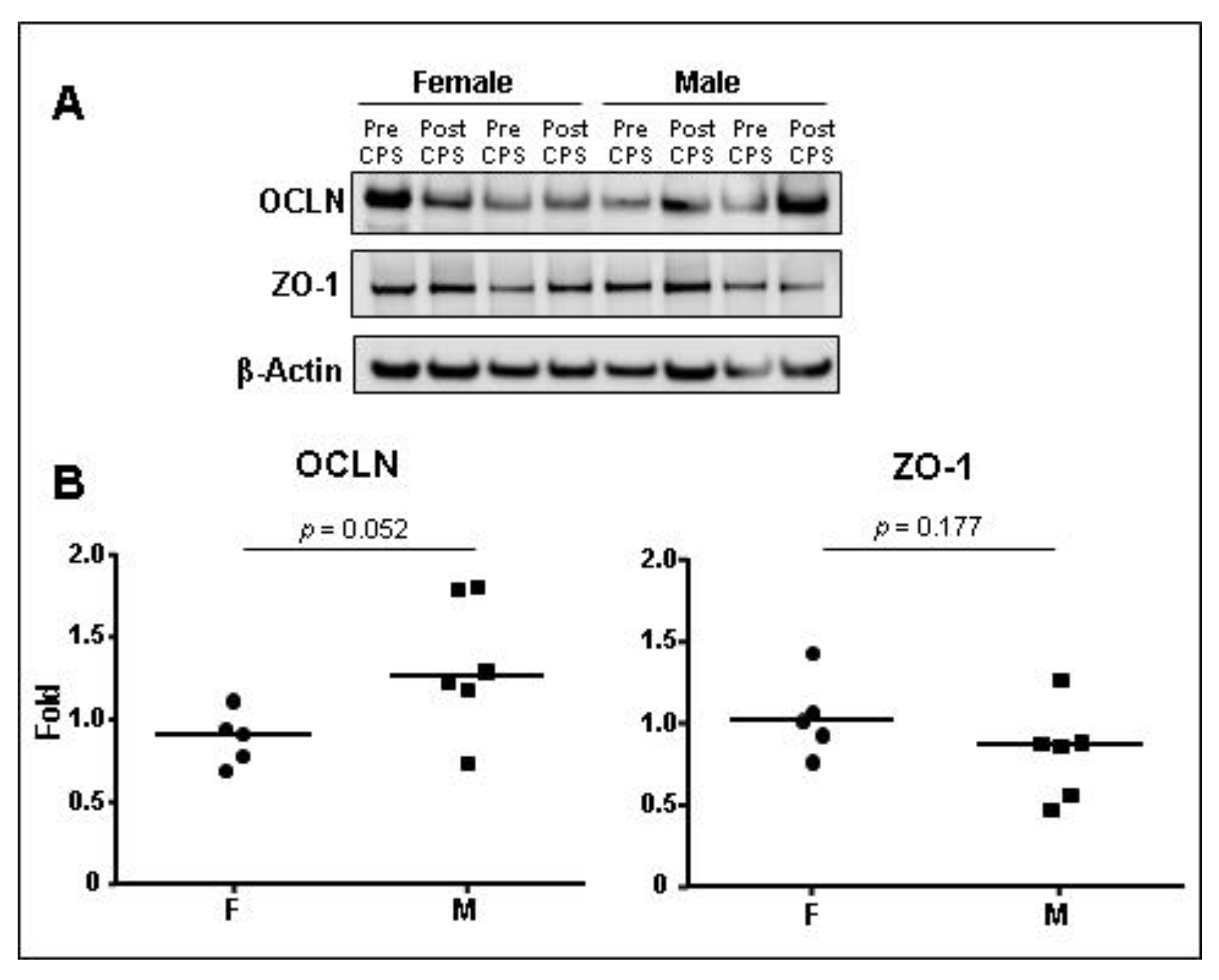
3.3.3. TJ Protein Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nater, U.M.; Maloney, E.; Heim, C.; Reeves, W.C. Cumulative Life Stress in Chronic Fatigue Syndrome. Psychiatry Res. 2011, 189, 318–320. [Google Scholar] [CrossRef] [PubMed]
- Enck, P.; Aziz, Q.; Barbara, G.; Farmer, A.D.; Fukudo, S.; Mayer, E.A.; Niesler, B.; Quigley, E.M.M.; Rajilić-Stojanović, M.; Schemann, M.; et al. Irritable Bowel Syndrome. Nat. Rev. Dis. Prim. 2016, 2, 16014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galvez-Sánchez, C.M.; Duschek, S.; Reyes Del Paso, G.A. Psychological Impact of Fibromyalgia: Current Perspectives. Psychol. Res. Behav. Manag. 2019, 12, 117–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, A.; Bhatt, R.R.; Naliboff, B.D.; Kutch, J.J.; Labus, J.S.; Vora, P.P.; Alaverdyan, M.; Schrepf, A.; Lutgendorf, S.; Mayer, E.A.; et al. Impact of Early Adverse Life Events and Sex on Functional Brain Networks in Patients with Urological Chronic Pelvic Pain Syndrome (UCPPS): A MAPP Research Network Study. PLoS ONE 2019, 14, e0217610. [Google Scholar] [CrossRef] [Green Version]
- Lacy, B.E.; Mearin, F.; Chang, L.; Chey, W.D.; Lembo, A.J.; Simren, M.; Spiller, R. Bowel Disorders. Gastroenterology 2016, 150, 1393–1407.e5. [Google Scholar] [CrossRef] [Green Version]
- Sperber, A.D.; Bangdiwala, S.I.; Drossman, D.A.; Ghoshal, U.C.; Simren, M.; Tack, J.; Whitehead, W.E.; Dumitrascu, D.L.; Fang, X.; Fukudo, S.; et al. Worldwide Prevalence and Burden of Functional Gastrointestinal Disorders, Results of Rome Foundation Global Study. Gastroenterology 2021, 160, 99–114.e3. [Google Scholar] [CrossRef]
- Chey, W.D. Food: The Main Course to Wellness and Illness in Patients with Irritable Bowel Syndrome. Am. J. Gastroenterol. 2016, 111, 366–371. [Google Scholar] [CrossRef]
- Dinan, T.G.; Cryan, J.; Shanahan, F.; Keeling, P.W.N.; Quigley, E.M.M. IBS: An Epigenetic Perspective. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 465–471. [Google Scholar] [CrossRef]
- Henström, M.; D’Amato, M. Genetics of Irritable Bowel Syndrome. Mol. Cell. Pediatr. 2016, 3, 7. [Google Scholar] [CrossRef] [Green Version]
- Gazouli, M.; Wouters, M.M.; Kapur-Pojskić, L.; Bengtson, M.-B.; Friedman, E.; Nikčević, G.; Demetriou, C.A.; Mulak, A.; Santos, J.; Niesler, B. Lessons Learned—Resolving the Enigma of Genetic Factors in IBS. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 77–87. [Google Scholar] [CrossRef]
- Rodiño-Janeiro, B.K.; Pardo-Camacho, C.; Santos, J.; Martínez, C. Mucosal RNA and Protein Expression as the next Frontier in IBS: Abnormal Function despite Morphologically Intact Small Intestinal Mucosa. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 316, G701–G719. [Google Scholar] [CrossRef]
- Guilarte, M.; Santos, J.; de Torres, I.; Alonso, C.; Vicario, M.; Ramos, L.; Martínez, C.; Casellas, F.; Saperas, E.; Malagelada, J.R. Diarrhoea-Predominant IBS Patients Show Mast Cell Activation and Hyperplasia in the Jejunum. Gut 2007, 56, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Guilarte, M.; Vicario, M.; Martínez, C.; de Torres, I.; Lobo, B.; Pigrau, M.; González-Castro, A.; Rodiño-Janeiro, B.K.; Salvo-Romero, E.; Fortea, M.; et al. Peripheral Corticotropin-Releasing Factor Triggers Jejunal Mast Cell Activation and Abdominal Pain in Patients With Diarrhea-Predominant Irritable Bowel Syndrome. Am. J. Gastroenterol. 2020, 115, 2047–2059. [Google Scholar] [CrossRef]
- Vicario, M.; González-Castro, A.M.; Martínez, C.; Lobo, B.; Pigrau, M.; Guilarte, M.; de Torres, I.; Mosquera, J.L.; Fortea, M.; Sevillano-Aguilera, C.; et al. Increased Humoral Immunity in the Jejunum of Diarrhoea-Predominant Irritable Bowel Syndrome Associated with Clinical Manifestations. Gut 2015, 64, 1379–1388. [Google Scholar] [CrossRef]
- Martin-Viñas, J.J.; Quigley, E.M.M. Immune Response in Irritable Bowel Syndrome: A Systematic Review of Systemic and Mucosal Inflammatory Mediators. J. Dig. Dis. 2016, 17, 572–581. [Google Scholar] [CrossRef]
- Martínez, C.; Rodiño-Janeiro, B.K.; Lobo, B.; Stanifer, M.L.; Klaus, B.; Granzow, M.; González-Castro, A.M.; Salvo-Romero, E.; Alonso-Cotoner, C.; Pigrau, M.; et al. MiR-16 and MiR-125b Are Involved in Barrier Function Dysregulation through the Modulation of Claudin-2 and Cingulin Expression in the Jejunum in IBS with Diarrhoea. Gut 2017, 66, 1537–1538. [Google Scholar] [CrossRef] [Green Version]
- Bashashati, M.; Moossavi, S.; Cremon, C.; Barbaro, M.R.; Moraveji, S.; Talmon, G.; Rezaei, N.; Hughes, P.A.; Bian, Z.X.; Choi, C.H.; et al. Colonic Immune Cells in Irritable Bowel Syndrome: A Systematic Review and Meta-Analysis. Neurogastroenterol. Motil. 2018, 30, e13192. [Google Scholar] [CrossRef]
- Mavrangelos, C.; Campaniello, M.A.; Andrews, J.M.; Bampton, P.A.; Hughes, P.A. Longitudinal Analysis Indicates Symptom Severity Influences Immune Profile in Irritable Bowel Syndrome. Gut 2018, 67, 398–399. [Google Scholar] [CrossRef]
- Martínez, C.; González-Castro, A.; Vicario, M.; Santos, J. Cellular and Molecular Basis of Intestinal Barrier Dysfunction in the Irritable Bowel Syndrome. Gut Liver 2012, 6, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Martínez, C.; Lobo, B.; Pigrau, M.; Ramos, L.; González-Castro, A.M.; Alonso, C.; Guilarte, M.; Guilá, M.; de Torres, I.; Azpiroz, F.; et al. Diarrhoea-Predominant Irritable Bowel Syndrome: An Organic Disorder with Structural Abnormalities in the Jejunal Epithelial Barrier. Gut 2013, 62, 1160–1168. [Google Scholar] [CrossRef]
- Vanuytsel, T.; van Wanrooy, S.; Vanheel, H.; Vanormelingen, C.; Verschueren, S.; Houben, E.; Salim Rasoel, S.; Tόth, J.; Holvoet, L.; Farré, R.; et al. Psychological Stress and Corticotropin-Releasing Hormone Increase Intestinal Permeability in Humans by a Mast Cell-Dependent Mechanism. Gut 2014, 63, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- Rodiño-Janeiro, B.K.; Alonso-Cotoner, C.; Pigrau, M.; Lobo, B.; Vicario, M.; Santos, J. Role of Corticotropin-Releasing Factor in Gastrointestinal Permeability. J. Neurogastroenterol. Motil. 2015, 21, 33–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barclay, G.R.; Turnberg, L.A. Effect of Psychological Stress on Salt and Water Transport in the Human Jejunum. Gastroenterology 1987, 93, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Barclay, G.R.; Turnberg, L.A. Effect of Cold-Induced Pain on Salt and Water Transport in the Human Jejunum. Gastroenterology 1988, 94, 994–998. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.; Saperas, E.; Nogueiras, C.; Mourelle, M.; Antolín, M.; Cadahia, A.; Malagelada, J.R. Release of Mast Cell Mediators into the Jejunum by Cold Pain Stress in Humans. Gastroenterology 1998, 114, 640–648. [Google Scholar] [CrossRef]
- Zheng, G.; Victor Fon, G.; Meixner, W.; Creekmore, A.; Zong, Y.; KDame, M.; Colacino, J.; Dedhia, P.H.; Hong, S.; Wiley, J.W. Chronic Stress and Intestinal Barrier Dysfunction: Glucocorticoid Receptor and Transcription Repressor HES1 Regulate Tight Junction Protein Claudin-1 Promoter. Sci. Rep. 2017, 7, 4502. [Google Scholar] [CrossRef] [Green Version]
- Alonso, C.; Guilarte, M.; Vicario, M.; Ramos, L.; Rezzi, S.; Martínez, C.; Lobo, B.; Martin, F.-P.; Pigrau, M.; González-Castro, A.M.; et al. Acute Experimental Stress Evokes a Differential Gender-Determined Increase in Human Intestinal Macromolecular Permeability. Neurogastroenterol. Motil. 2012, 24, 740–746, e348–e349. [Google Scholar] [CrossRef]
- Holmes, T.H.; Rahe, R.H. The Social Readjustment Rating Scale. J. Psychosom. Res. 1967, 11, 213–218. [Google Scholar] [CrossRef]
- Cohen, S.; Kamarck, T.; Mermelstein, R. A Global Measure of Perceived Stress. J. Health Soc. Behav. 1983, 24, 385–396. [Google Scholar] [CrossRef]
- Beck, A.T.; Ward, C.H.; Mendelson, M.; Mock, J.; Erbaugh, J. An Inventory for Measuring Depression. Arch. Gen. Psychiatry 1961, 4, 561–571. [Google Scholar] [CrossRef]
- Lovallo, W. The Cold Pressor Test and Autonomic Function: A Review and Integration. Psychophysiology 1975, 12, 268–282. [Google Scholar] [CrossRef]
- Naliboff, B.D.; Benton, D.; Solomon, G.F.; Morley, J.E.; Fahey, J.L.; Bloom, E.T.; Makinodan, T.; Gilmore, S.L. Immunological Changes in Young and Old Adults during Brief Laboratory Stress. Psychosom. Med. 1991, 53, 121–132. [Google Scholar] [CrossRef]
- Alonso, C.; Guilarte, M.; Vicario, M.; Ramos, L.; Ramadan, Z.; Antolín, M.; Martínez, C.; Rezzi, S.; Saperas, E.; Kochhar, S.; et al. Maladaptive Intestinal Epithelial Responses to Life Stress May Predispose Healthy Women to Gut Mucosal Inflammation. Gastroenterology 2008, 135, 163–172.e1. [Google Scholar] [CrossRef]
- Bekhbat, M.; Neigh, G.N. Sex Differences in the Neuro-Immune Consequences of Stress: Focus on Depression and Anxiety. Brain Behav. Immun. 2018, 67, 1–12. [Google Scholar] [CrossRef]
- Meester, I.; Rivera-Silva, G.F.; González-Salazar, F. Immune System Sex Differences May Bridge the Gap Between Sex and Gender in Fibromyalgia. Front. Neurosci. 2019, 13, 1414. [Google Scholar] [CrossRef]
- Zheng, G.; Wu, S.-P.; Hu, Y.; Smith, D.E.; Wiley, J.W.; Hong, S. Corticosterone Mediates Stress-Related Increased Intestinal Permeability in a Region-Specific Manner. Neurogastroenterol. Motil. 2013, 25, e127–e139. [Google Scholar] [CrossRef]
- Matsuo, K.; Zhang, X.; Ono, Y.; Nagatomi, R. Acute Stress-Induced Colonic Tissue HSP70 Expression Requires Commensal Bacterial Components and Intrinsic Glucocorticoid. Brain Behav. Immun. 2009, 23, 108–115. [Google Scholar] [CrossRef]
- Creekmore, A.L.; Hong, S.; Zhu, S.; Xue, J.; Wiley, J.W. Chronic Stress-Associated Visceral Hyperalgesia Correlates with Severity of Intestinal Barrier Dysfunction. Pain 2018, 159, 1777–1789. [Google Scholar] [CrossRef]
- Zong, Y.; Zhu, S.; Zhang, S.; Zheng, G.; Wiley, J.W.; Hong, S. Chronic Stress and Intestinal Permeability: Lubiprostone Regulates Glucocorticoid Receptor-Mediated Changes in Colon Epithelial Tight Junction Proteins, Barrier Function, and Visceral Pain in the Rodent and Human. Neurogastroenterol. Motil. 2018, 31, e13477. [Google Scholar] [CrossRef]
- Kawedia, J.D.; Nieman, M.L.; Boivin, G.P.; Melvin, J.E.; Kikuchi, K.-I.; Hand, A.R.; Lorenz, J.N.; Menon, A.G. Interaction between Transcellular and Paracellular Water Transport Pathways through Aquaporin 5 and the Tight Junction Complex. Proc. Natl. Acad. Sci. USA 2007, 104, 3621–3626. [Google Scholar] [CrossRef]
- Lee, H.S.; Kim, D.-K.; Kim, Y.B.; Lee, K.J. Effect of Acute Stress on Immune Cell Counts and the Expression of Tight Junction Proteins in the Duodenal Mucosa of Rats. Gut Liver 2013, 7, 190–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.H.; Xiao, K.; Luan, Z.S.; Song, J. Early Weaning Increases Intestinal Permeability, Alters Expression of Cytokine and Tight Junction Proteins, and Activates Mitogen-Activated Protein Kinases in Pigs. J. Anim. Sci. 2013, 91, 1094–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Itallie, C.M.; Fanning, A.S.; Bridges, A.; Anderson, J.M. ZO-1 Stabilizes the Tight Junction Solute Barrier through Coupling to the Perijunctional Cytoskeleton. Mol. Biol. Cell 2009, 20, 3930–3940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulzke, J.D.; Gitter, A.H.; Mankertz, J.; Spiegel, S.; Seidler, U.; Amasheh, S.; Saitou, M.; Tsukita, S.; Fromm, M. Epithelial Transport and Barrier Function in Occludin-Deficient Mice. Biochim. Biophys. Acta Biomembr. 2005, 1669, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Saitou, M.; Fujimoto, K.; Doi, Y.; Itoh, M.; Fujimoto, T.; Furuse, M.; Takano, H.; Noda, T.; Tsukita, S. Occludin-Deficient Embryonic Stem Cells Can Differentiate into Polarized Epithelial Cells Bearing Tight Junctions. J. Cell Biol. 1998, 141, 397–408. [Google Scholar] [CrossRef]
- Raleigh, D.R.; Boe, D.M.; Yu, D.; Weber, C.R.; Marchiando, A.M.; Bradford, E.M.; Wang, Y.; Wu, L.; Schneeberger, E.E.; Shen, L.; et al. Occludin S408 Phosphorylation Regulates Tight Junction Protein Interactions and Barrier Function. J. Cell Biol. 2011, 193, 565–582. [Google Scholar] [CrossRef]
- Buschmann, M.M.; Shen, L.; Rajapakse, H.; Raleigh, D.R.; Wang, Y.; Wang, Y.; Lingaraju, A.; Zha, J.; Abbott, E.; McAuley, E.M.; et al. Occludin OCEL-Domain Interactions Are Required for Maintenance and Regulation of the Tight Junction Barrier to Macromolecular Flux. MBoC 2013, 24, 3056–3068. [Google Scholar] [CrossRef]
- Tamura, A.; Hayashi, H.; Imasato, M.; Yamazaki, Y.; Hagiwara, A.; Wada, M.; Noda, T.; Watanabe, M.; Suzuki, Y.; Tsukita, S. Loss of Claudin-15, but Not Claudin-2, Causes Na+ Deficiency and Glucose Malabsorption in Mouse Small Intestine. Gastroenterology 2011, 140, 913–923. [Google Scholar] [CrossRef]
- Van Itallie, C.M.; Holmes, J.; Bridges, A.; Gookin, J.L.; Coccaro, M.R.; Proctor, W.; Colegio, O.R.; Anderson, J.M. The Density of Small Tight Junction Pores Varies among Cell Types and Is Increased by Expression of Claudin-2. J. Cell. Sci. 2008, 121, 298–305. [Google Scholar] [CrossRef] [Green Version]
- Inai, T.; Kobayashi, J.; Shibata, Y. Claudin-1 Contributes to the Epithelial Barrier Function in MDCK Cells. Eur. J. Cell Biol. 1999, 78, 849–855. [Google Scholar] [CrossRef]
- Zuo, L.; Kuo, W.-T.; Turner, J.R. Tight Junctions as Targets and Effectors of Mucosal Immune Homeostasis. Cell. Mol. Gastroenterol. Hepatol. 2020, 10, 327–340. [Google Scholar] [CrossRef]
- Meddings, J.B.; Swain, M.G. Environmental Stress-Induced Gastrointestinal Permeability Is Mediated by Endogenous Glucocorticoids in the Rat. Gastroenterology 2000, 119, 1019–1028. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, Z.-Q.; Liu, X.-Y.; Yang, L.; Geng, X.-R.; Yang, G.; Liu, Z.-G.; Zheng, P.-Y.; Yang, P.-C. Stress-Derived Corticotropin Releasing Factor Breaches Epithelial Endotoxin Tolerance. PLoS ONE 2013, 8, e65760. [Google Scholar] [CrossRef] [Green Version]
- Bangasser, D.A.; Valentino, R.J. Sex Differences in Stress-Related Psychiatric Disorders: Neurobiological Perspectives. Front. Neuroendocrinol. 2014, 35, 303–319. [Google Scholar] [CrossRef] [Green Version]
- Vicario, M.; Guilarte, M.; Alonso, C.; Yang, P.; Martínez, C.; Ramos, L.; Lobo, B.; González, A.; Guilà, M.; Pigrau, M.; et al. Chronological Assessment of Mast Cell-Mediated Gut Dysfunction and Mucosal Inflammation in a Rat Model of Chronic Psychosocial Stress. Brain Behav. Immun. 2010, 24, 1166–1175. [Google Scholar] [CrossRef]
- Söderholm, J.D.; Yang, P.-C.; Ceponis, P.; Vohra, A.; Riddell, R.; Sherman, P.M.; Perdue, M.H. Chronic Stress Induces Mast Cell-Dependent Bacterial Adherence and Initiates Mucosal Inflammation in Rat Intestine. Gastroenterology 2002, 123, 1099–1108. [Google Scholar] [CrossRef]
- Kaushik, S.; Kaur, J. Effect of Chronic Cold Stress on Intestinal Epithelial Cell Proliferation and Inflammation in Rats. Stress 2005, 8, 191–197. [Google Scholar] [CrossRef]
- Santos, J.; Yang, P.C.; Söderholm, J.D.; Benjamin, M.; Perdue, M.H. Role of Mast Cells in Chronic Stress Induced Colonic Epithelial Barrier Dysfunction in the Rat. Gut 2001, 48, 630–636. [Google Scholar] [CrossRef] [Green Version]
- Smith, F.; Clark, J.E.; Overman, B.L.; Tozel, C.C.; Huang, J.H.; Rivier, J.E.F.; Blikslager, A.T.; Moeser, A.J. Early Weaning Stress Impairs Development of Mucosal Barrier Function in the Porcine Intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G352–G363. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.; Yates, D.; Guilarte, M.; Vicario, M.; Alonso, C.; Perdue, M.H. Stress Neuropeptides Evoke Epithelial Responses via Mast Cell Activation in the Rat Colon. Psychoneuroendocrinology 2008, 33, 1248–1256. [Google Scholar] [CrossRef]
- Santos, J.; Benjamin, M.; Yang, P.C.; Prior, T.; Perdue, M.H. Chronic Stress Impairs Rat Growth and Jejunal Epithelial Barrier Function: Role of Mast Cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 278, G847–G854. [Google Scholar]
- Wallon, C.; Yang, P.-C.; Keita, A.V.; Ericson, A.-C.; McKay, D.M.; Sherman, P.M.; Perdue, M.H.; Söderholm, J.D. Corticotropin-Releasing Hormone (CRH) Regulates Macromolecular Permeability via Mast Cells in Normal Human Colonic Biopsies in Vitro. Gut 2008, 57, 50–58. [Google Scholar] [CrossRef]
- Jacob, C.; Yang, P.-C.; Darmoul, D.; Amadesi, S.; Saito, T.; Cottrell, G.S.; Coelho, A.-M.; Singh, P.; Grady, E.F.; Perdue, M.; et al. Mast Cell Tryptase Controls Paracellular Permeability of the Intestine. Role of Protease-Activated Receptor 2 and Beta-Arrestins. J. Biol. Chem. 2005, 280, 31936–31948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldblum, S.E.; Rai, U.; Tripathi, A.; Thakar, M.; De Leo, L.; Di Toro, N.; Not, T.; Ramachandran, R.; Puche, A.C.; Hollenberg, M.D.; et al. The Active Zot Domain (Aa 288-293) Increases ZO-1 and Myosin 1C Serine/Threonine Phosphorylation, Alters Interaction between ZO-1 and Its Binding Partners, and Induces Tight Junction Disassembly through Proteinase Activated Receptor 2 Activation. FASEB J. 2011, 25, 144–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergnolle, N. Protease Inhibition as New Therapeutic Strategy for GI Diseases. Gut 2016, 65, 1215–1224. [Google Scholar] [CrossRef] [Green Version]
- Ray, K. IBS: The Power of Protease Activity in IBS. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 139. [Google Scholar] [CrossRef]
- Rolland-Fourcade, C.; Denadai-Souza, A.; Cirillo, C.; Lopez, C.; Jaramillo, J.O.; Desormeaux, C.; Cenac, N.; Motta, J.-P.; Larauche, M.; Taché, Y.; et al. Epithelial Expression and Function of Trypsin-3 in Irritable Bowel Syndrome. Gut 2017, 66, 1767–1778. [Google Scholar] [CrossRef]
- Lau, C.; Lytle, C.; Straus, D.S.; DeFea, K.A. Apical and Basolateral Pools of Proteinase-Activated Receptor-2 Direct Distinct Signaling Events in the Intestinal Epithelium. Am. J. Physiol. Cell Physiol. 2011, 300, C113–C123. [Google Scholar] [CrossRef]
- Greene, C.M.; Marciniak, S.J.; Teckman, J.; Ferrarotti, I.; Brantly, M.L.; Lomas, D.A.; Stoller, J.K.; McElvaney, N.G. A1-Antitrypsin Deficiency. Nat. Rev. Dis. Primers 2016, 2, 16051. [Google Scholar] [CrossRef]
- Geboes, K.; Ray, M.B.; Rutgeerts, P.; Callea, F.; Desmet, V.J.; Vantrappen, G. Morphological Identification of Alpha-I-Antitrypsin in the Human Small Intestine. Histopathology 1982, 6, 55–60. [Google Scholar] [CrossRef]
- Henley, D.E.; Lightman, S.L. New Insights into Corticosteroid-Binding Globulin and Glucocorticoid Delivery. Neuroscience 2011, 180, 1–8. [Google Scholar] [CrossRef]
- Bolton, J.L.; Hayward, C.; Direk, N.; Lewis, J.G.; Hammond, G.L.; Hill, L.A.; Anderson, A.; Huffman, J.; Wilson, J.F.; Campbell, H.; et al. Genome Wide Association Identifies Common Variants at the SERPINA6/SERPINA1 Locus Influencing Plasma Cortisol and Corticosteroid Binding Globulin. PLoS Genet. 2014, 10, e1004474. [Google Scholar] [CrossRef] [Green Version]
- Mackey, E.; Ayyadurai, S.; Pohl, C.S.; D’Costa, S.; Li, Y.; Moeser, A.J. Sexual Dimorphism in the Mast Cell Transcriptome and the Pathophysiological Responses to Immunological and Psychological Stress. Biol. Sex Differ. 2016, 7, 60. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | |||
|---|---|---|---|
| Variable | M (Q1–Q3) | F (Q1–Q3) | p Value |
| Age (years) | 22.9 (22.1–29.2) | 22.6 (21.7–25.2) | 0.462 |
| Holmes–Rahe scale (score) | 76.0 (39.5–98.5) | 89.0 (63.8–131.3) | 0.420 |
| Cohen’s scale (score) | 14.0 (11.0–23.5) | 19.0 (12.3–24.0) | 0.529 |
| Beck’s inventory (score) | 0.0 (0.0–0.5) | 0.5 (0.0–2.8) | 0.737 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodiño-Janeiro, B.K.; Pigrau, M.; Salvo-Romero, E.; Nieto, A.; Expósito, E.; González-Castro, A.M.; Galán, C.; de Torres, I.; Pribic, T.; Hernández, L.; et al. Acute Stress Regulates Sex-Related Molecular Responses in the Human Jejunal Mucosa: Implications for Irritable Bowel Syndrome. Cells 2023, 12, 423. https://doi.org/10.3390/cells12030423
Rodiño-Janeiro BK, Pigrau M, Salvo-Romero E, Nieto A, Expósito E, González-Castro AM, Galán C, de Torres I, Pribic T, Hernández L, et al. Acute Stress Regulates Sex-Related Molecular Responses in the Human Jejunal Mucosa: Implications for Irritable Bowel Syndrome. Cells. 2023; 12(3):423. https://doi.org/10.3390/cells12030423
Chicago/Turabian StyleRodiño-Janeiro, Bruno K., Marc Pigrau, Eloísa Salvo-Romero, Adoración Nieto, Elba Expósito, Ana M. González-Castro, Carmen Galán, Inés de Torres, Teodora Pribic, Laura Hernández, and et al. 2023. "Acute Stress Regulates Sex-Related Molecular Responses in the Human Jejunal Mucosa: Implications for Irritable Bowel Syndrome" Cells 12, no. 3: 423. https://doi.org/10.3390/cells12030423