

Light-Enhanced Cytotoxicity of Doxorubicin by Photoactivation
 , , , , , , , , ,
, , , , , , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
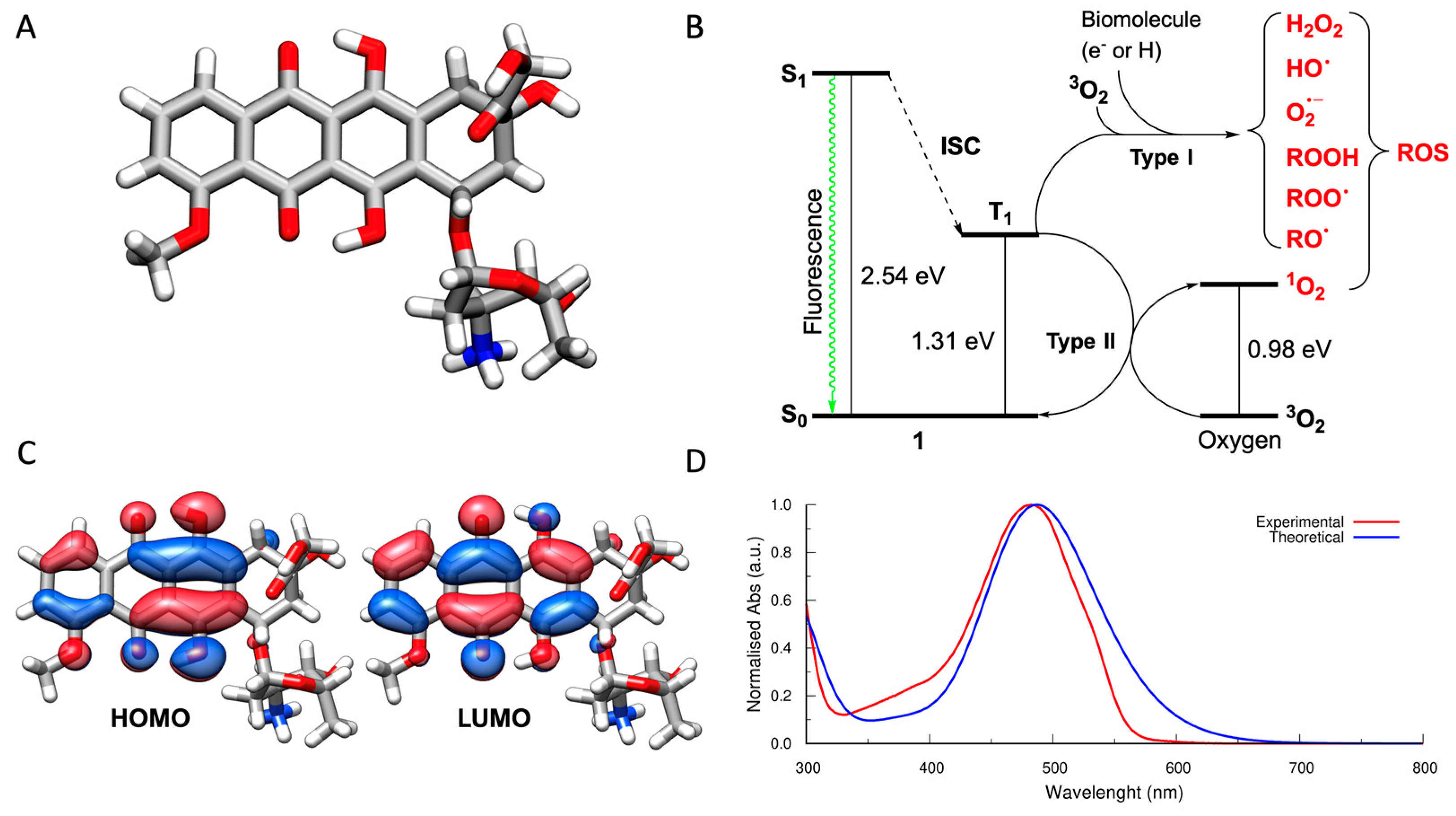
2.1. Computational Details
2.2. Quantification of Reactive Oxygen Species
2.3. Cell Culture
2.4. Cell Treatment and Irradiation
2.5. Cell Viability Assays
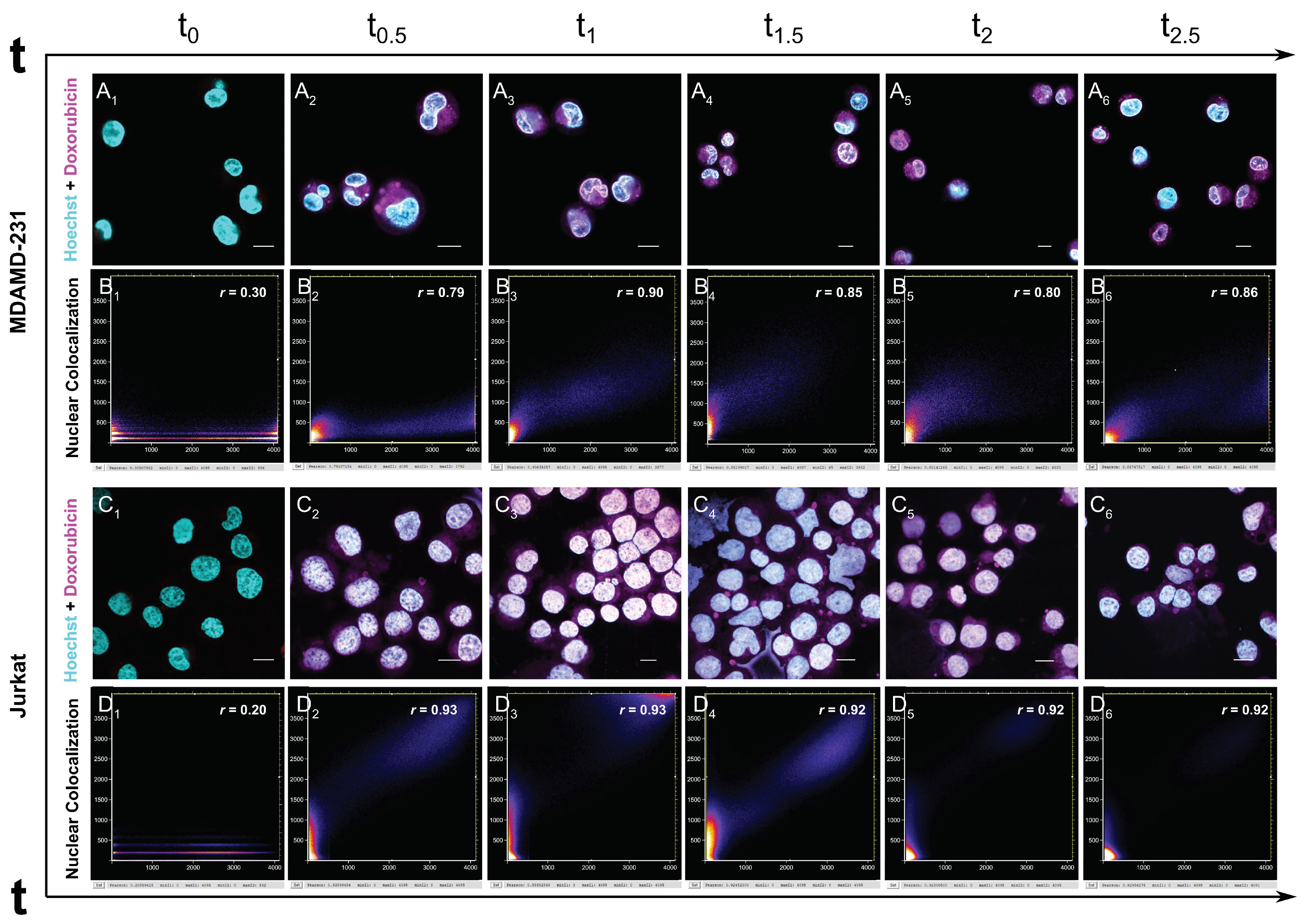
2.6. Microscopic Analysis
2.7. Analysis of Caspase-3 Activity
2.8. Necrotic Cell Death Determined by SYTOX™ Green Nucleic Acid Stain
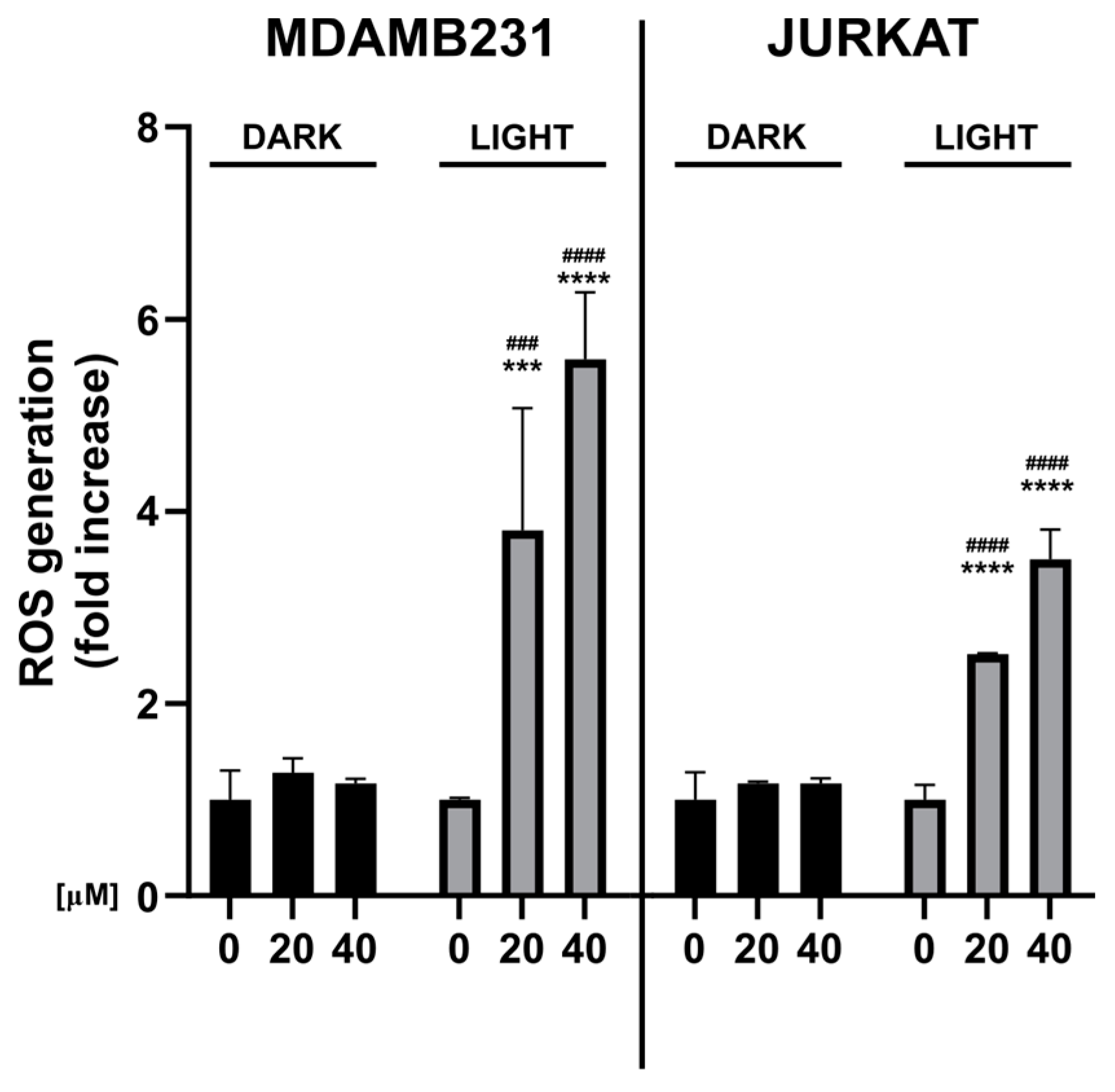
2.9. Intracellular ROS Generation
2.10. Statistical Analysis
3. Results and Discussion
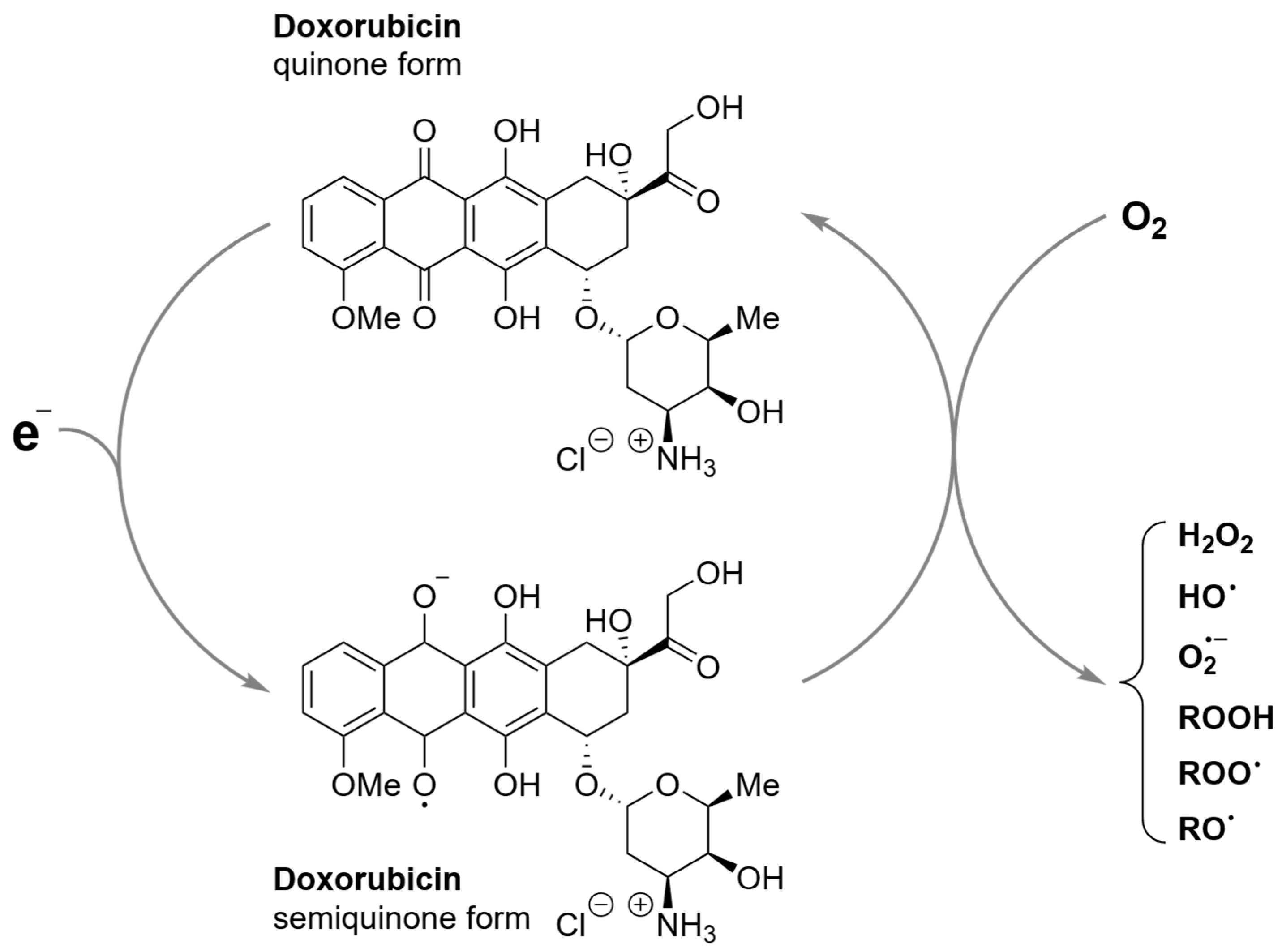
3.1. Light-Dependent Generation of ROS by Doxorubicin
3.2. Photoactivation Enhances the Cytotoxic Activity of Doxorubicin
3.3. Uptake and Nuclear Localization of Doxorubicin
3.4. Photoactivated Doxorubicin Induces Both Caspase-Dependent and -Independent Cell Death
3.5. Photoactivated Doxorubicin Generates Intracellular ROS
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gunaydin, G.; Gedik, M.E.; Ayan, S. Photodynamic Therapy for the Treatment and Diagnosis of Cancer–A Review of the Current Clinical Status. Front. Chem. 2021, 9, 686303. [Google Scholar] [CrossRef] [PubMed]
- van Straten, D.; Mashayekhi, V.; de Bruijn, H.S.; Oliveira, S.; Robinson, D.J. Oncologic Photodynamic Therapy: Basic Principles, Current Clinical Status and Future Directions. Cancers 2017, 9, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, D.; Carter, K.A.; Miranda, D.; Lovell, J.F. Chemophototherapy: An Emerging Treatment Option for Solid Tumors. Adv. Sci. 2017, 4, 1600106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunaydin, G.; Gedik, M.E.; Ayan, S. Photodynamic Therapy—Current Limitations and Novel Approaches. Front. Chem. 2021, 9, 691697. [Google Scholar] [CrossRef]
- Thorn, C.F.; Oshiro, C.; Marsh, S.; Hernandez-Boussard, T.; McLeod, H.; Klein, T.E.; Altman, R.B. Doxorubicin Pathways: Pharmacodynamics and Adverse Effects. Pharm. Genom. 2011, 21, 440–446. [Google Scholar] [CrossRef]
- Meredith, A.-M.; Dass, C.R. Increasing Role of the Cancer Chemotherapeutic Doxorubicin in Cellular Metabolism. J. Pharm. Pharmacol. 2016, 68, 729–741. [Google Scholar] [CrossRef] [Green Version]
- van der Zanden, S.Y.; Qiao, X.; Neefjes, J. New Insights into the Activities and Toxicities of the Old Anticancer Drug Doxorubicin. FEBS J. 2021, 288, 6095–6111. [Google Scholar] [CrossRef]
- Streckyte, G.; Didziapetriene, J.; Grazeliene, G.; Prasmickiene, G.; Sukeliene, D.; Kazlauskaite, N.; Characiejus, D.; Griciute, L.; Rotomskis, R. Effects of Photodynamic Therapy in Combination with Adriamycin. Cancer Lett. 1999, 146, 73–86. [Google Scholar] [CrossRef]
- Brophy, P.F.; Keller, S.M. Adriamycin Enhanced in vitro and in vivo Photodynamic Therapy of Mesothelioma. J. Surg. Res. 1992, 52, 631–634. [Google Scholar] [CrossRef]
- Casas, A.; Fukuda, H.; Riley, P.; Alcira, A.M. Enhancement of Aminolevulinic Acid Based Photodynamic Therapy by Adriamycin. Cancer Lett. 1997, 121, 105–113. [Google Scholar] [CrossRef]
- Canti, G.; Nicolin, A.; Cubeddu, R.; Taroni, P.; Bandieramonte, G.; Valentini, G. Antitumor Efficacy of the Combination of Photodynamic Therapy and Chemotherapy in Murine Tumors. Cancer Lett. 1998, 125, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Nahabedian, M.Y.; Cohen, R.A.; Contino, M.F.; Terem, T.M.; Wright, W.H.; Berns, M.W.; Wile, A.G. Combination Cytotoxic Chemotherapy with Cisplatin or Doxorubicin and Photodynamic Therapy in Murine Tumors. J. Natl. Cancer Inst. 1988, 80, 739–743. [Google Scholar] [CrossRef] [PubMed]
- Yousefi Sadeghloo, A.; Khorsandi, K.; Kianmehr, Z. Synergistic Effect of Photodynamic Treatment and Doxorubicin on Triple Negative Breast Cancer Cells. Photochem. Photobiol. Sci. 2020, 19, 1580–1589. [Google Scholar] [CrossRef] [PubMed]
- Cacaccio, J.C.; Durrani, F.A.; Missert, J.R.; Pandey, R.K. Photodynamic Therapy in Combination with Doxorubicin Is Superior to Monotherapy for the Treatment of Lung Cancer. Biomedicines 2022, 10, 857. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-González, R.; Milán, P.; Bresolí-Obach, R.; Stockert, J.C.; Villanueva, A.; Cañete, M.; Nonell, S. Photodynamic Synergistic Effect of Pheophorbide a and Doxorubicin in Combined Treatment against Tumoral Cells. Cancers 2017, 9, 18. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.L.; Syu, W.J.; Nishiyama, N.; Kataoka, K.; Lai, P.S. Dendrimer Phthalocyanine-Encapsulated Polymeric Micelle-Mediated Photochemical Internalization Extends the Efficacy of Photodynamic Therapy and Overcomes Drug-Resistance in Vivo. J. Control. Release 2011, 155, 458–464. [Google Scholar] [CrossRef]
- Snyder, J.W.; Greco, W.R.; Bellnier, D.A.; Vaughan, L.; Henderson, B.W. Photodynamic Therapy: A Means to Enhanced Drug Delivery to Tumors. Cancer Res. 2003, 63, 8126–8131. [Google Scholar]
- Khdair, A.; Chen, D.; Patil, Y.; Ma, L.; Dou, Q.P.; Shekhar, M.P.V.; Panyam, J. Nanoparticle-Mediated Combination Chemotherapy and Photodynamic Therapy Overcomes Tumor Drug Resistance. J. Control. Release 2010, 141, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Cowled, P.A.; Mackenzie, L.; Forbes, I.J. Pharmacological Modulation of Photodynamic Therapy with Hematoporphyrin Derivative and Light. Cancer Res 1987, 47, 971–974. [Google Scholar]
- Shah, S.; Chandra, A.; Kaur, A.; Sabnis, N.; Lacko, A.; Gryczynski, Z.; Fudala, R.; Gryczynski, I. Fluorescence Properties of Doxorubicin in PBS Buffer and PVA Films. J. Photochem. Photobiol. B 2017, 170, 1580–1589. [Google Scholar] [CrossRef] [Green Version]
- Lanks, K.W.; Gao, J.P.; Sharma, T. Photodynamic Enhancement of Doxorubicin Cytotoxicity. Cancer Chemother Pharm. 1994, 35, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.P.; Lanks, K.W.; Rosen, M.; Lai, B.T. Mechanism of Action and Spectrum of Cell Types Susceptible to Doxorubicin Photochemotherapy. Cancer Chemother Pharm. 1997, 40, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Quintos-Meneses, H.A.; Aranda-Lara, L.; Morales-Ávila, E.; Torres-García, E.; Camacho-López, M.Á.; Sánchez-Holguín, M.; Luna-Gutiérrez, M.A.; Ramírez-Durán, N.; Isaac-Olivé, K. In Vitro Irradiation of Doxorubicin with 18F-FDG Cerenkov Radiation and Its Potential Application as a Theragnostic System. J. Photochem. Photobiol. B 2020, 210, 111961. [Google Scholar] [CrossRef] [PubMed]
- Andreoni, A.; Colasanti, A.; Malatesta, V.; Roberti, G. Photocytotoxicity of Anthracyclines upon Laser Excitation in Their Long-Wavelength Absorption Bands. Radiat. Res. 1991, 127, 24–29. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.E.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Farkas, O.; et al. Gaussian 16; Gaussian, Inc.: Wallingford, CT, USA, 2016. [Google Scholar]
- Becke, A.D. Density-Functional Thermochemistry. III. The Role of Exact Exchange. J. Chem. Phys. 1993, 98, 5648. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, R.; Binkley, J.S.; Seeger, R.; Pople, J.A. Self-Consistent Molecular Orbital Methods. XX. A Basis Set for Correlated Wave Functions. J. Chem. Phys. 1980, 72, 650. [Google Scholar] [CrossRef]
- Runge, E.; Gross, E.K.U. Density-Functional Theory for Time-Dependent Systems. Phys. Rev. Lett. 1984, 52, 997. [Google Scholar] [CrossRef]
- Tomasi, J.; Mennucci, B.; Cammi, R. Quantum Mechanical Continuum Solvation Models. Chem. Rev. 2005, 105, 2999–3094. [Google Scholar] [CrossRef]
- Cantelli, A.; Piro, F.; Pecchini, P.; di Giosia, M.; Danielli, A.; Calvaresi, M. Concanavalin A-Rose Bengal Bioconjugate for Targeted Gram-Negative Antimicrobial Photodynamic Therapy. J. Photochem. Photobiol. B 2020, 206, 111852. [Google Scholar] [CrossRef]
- Ulfo, L.; Cantelli, A.; Petrosino, A.; Costantini, P.E.; Nigro, M.; Starinieri, F.; Turrini, E.; Zadran, S.K.; Zuccheri, G.; Saporetti, R.; et al. Orthogonal Nanoarchitectonics of M13 Phage for Receptor Targeted Anticancer Photodynamic Therapy. Nanoscale 2022, 14, 632–641. [Google Scholar] [CrossRef]
- Cantelli, A.; Malferrari, M.; Soldà, A.; Simonetti, G.; Forni, S.; Toscanella, E.; Mattioli, E.J.; Zerbetto, F.; Zanelli, A.; di Giosia, M.; et al. Human Serum Albumin–Oligothiophene Bioconjugate: A Phototheranostic Platform for Localized Killing of Cancer Cells by Precise Light Activation. JACS Au 2021, 1, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Di Giosia, M.; Soldà, A.; Seeger, M.; Cantelli, A.; Arnesano, F.; Nardella, M.I.; Mangini, V.; Valle, F.; Montalti, M.; Zerbetto, F.; et al. A Bio-Conjugated Fullerene as a Subcellular-Targeted and Multifaceted Phototheranostic Agent. Adv. Funct. Mater. 2021, 31, 2101527. [Google Scholar] [CrossRef]
- di Giosia, M.; Nicolini, F.; Ferrazzano, L.; Soldà, A.; Valle, F.; Cantelli, A.; Marforio, T.D.; Bottoni, A.; Zerbetto, F.; Montalti, M.; et al. Stable and Biocompatible Monodispersion of C60 in Water by Peptides. Bioconjug. Chem. 2019, 30, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Bortot, B.; Apollonio, M.; Baj, G.; Andolfi, L.; Zupin, L.; Crovella, S.; di Giosia, M.; Cantelli, A.; Saporetti, R.; Ulfo, L.; et al. Advanced Photodynamic Therapy with an Engineered M13 Phage Targeting EGFR: Mitochondrial Localization and Autophagy Induction in Ovarian Cancer Cell Lines. Free Radic. Biol. Med. 2022, 179, 242–251. [Google Scholar] [CrossRef]
- Cantelli, A.; Malferrari, M.; Mattioli, E.J.; Marconi, A.; Mirra, G.; Soldà, A.; Marforio, T.D.; Zerbetto, F.; Rapino, S.; di Giosia, M.; et al. Enhanced Uptake and Phototoxicity of C60@albumin Hybrids by Folate Bioconjugation. Nanomaterials 2022, 12, 3501. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Davies, K.J.A.; Doroshow, J.H. Redox Cycling of Anthracyclines by Cardiac Mitochondria. I. Anthracycline Radical Formation by NADH Dehydrogenase. J. Biol. Chem. 1986, 261, 3060–3067. [Google Scholar] [CrossRef]
- Doroshow, J.H.; Davies, K.J.A. Redox Cycling of Anthracyclines by Cardiac Mitochondria. II. Formation of Superoxide Anion, Hydrogen Peroxide, and Hydroxyl Radical. J. Biol. Chem. 1986, 261, 3068–3074. [Google Scholar] [CrossRef]
- Berthiaume, J.M.; Wallace, K.B. Adriamycin-Induced Oxidative Mitochondrial Cardiotoxicity. Cell Biol. Toxicol. 2007, 23, 15–25. [Google Scholar] [CrossRef]
- Ulfo, L.; Costantini, P.E.; di Giosia, M.; Danielli, A.; Calvaresi, M. EGFR-Targeted Photodynamic Therapy. Pharmaceutics 2022, 14, 241. [Google Scholar] [CrossRef]
- Qiu, H.; Tan, M.; Ohulchanskyy, T.Y.; Lovell, J.F.; Chen, G. Recent Progress in Upconversion Photodynamic Therapy. Nanomaterials 2018, 8, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shafirstein, G.; Bellnier, D.; Oakley, E.; Hamilton, S.; Potasek, M.; Beeson, K.; Parilov, E. Interstitial Photodynamic Therapy—A Focused Review. Cancers 2017, 9, 12. [Google Scholar] [CrossRef]
- Doxorubicin Hydrochloride. Available online: https://www.Cancer.Gov/about-Cancer/Treatment/Drugs/Doxorubicinhydrochloride (accessed on 2 December 2022).
- Yu, K.D.; Cai, Y.W.; Wu, S.Y.; Shui, R.H.; Shao, Z.M. Estrogen Receptor-Low Breast Cancer: Biology Chaos and Treatment Paradox. Cancer Commun. 2021, 41, 968–980. [Google Scholar] [CrossRef] [PubMed]
- Fleisher, B.; Lezeau, J.; Werkman, C.; Jacobs, B.; Ait-Oudhia, S. In Vitro to Clinical Translation of Combinatorial Effects of Doxorubicin and Abemaciclib in Rb-Positive Triple Negative Breast Cancer: A Systems-Based Pharmacokinetic/Pharmacodynamic Modeling Approach. Breast Cancer Targets Ther. 2021, 13, 87–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Li, X.; Yoon, J. Organelle-Targeted Photosensitizers for Precision Photodynamic Therapy. ACS Appl. Mater. Interfaces 2021, 13, 19543–19571. [Google Scholar] [CrossRef]
- Akhlynina, T.V.; Jans, D.A.; Rosenkranz, A.A.; Statsyuk, N.V.; Balashova, I.Y.; Toth, G.; Pavo, I.; Rubin, A.B.; Sobolev, A.S. Nuclear Targeting of Chlorin E6 Enhances Its Photosensitizing Activity. J. Biol. Chem. 1997, 272, 20328–20331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnabosco, G.; di Giosia, M.; Polishchuk, I.; Weber, E.; Fermani, S.; Bottoni, A.; Zerbetto, F.; Pelicci, P.G.; Pokroy, B.; Rapino, S.; et al. Calcite Single Crystals as Hosts for Atomic-Scale Entrapment and Slow Release of Drugs. Adv. Heal. Mater 2015, 4, 1510–1516. [Google Scholar] [CrossRef] [Green Version]
- Magnabosco, G.; Ianiro, A.; Stefani, D.; Soldà, A.; Rapino, S.; Falini, G.; Calvaresi, M. Doxorubicin-Loaded Squid Pen Plaster: A Natural Drug Delivery System for Cancer Cells. ACS Appl. Bio Mater. 2020, 3, 1514–1519. [Google Scholar] [CrossRef]
- Skladanowski, A.; Konopa, J. Adriamycin and Daunomycin Induce Programmed Cell Death (Apoptosis) in Tumour Cells. Biochem. Pharm. 1993, 46, 375–382. [Google Scholar] [CrossRef]
- Di, X.; Shiu, R.P.; Newsham, I.F.; Gewirtz, D.A. Apoptosis, Autophagy, Accelerated Senescence and Reactive Oxygen in the Response of Human Breast Tumor Cells to Adriamycin. Biochem. Pharm. 2009, 77, 1139–1150. [Google Scholar] [CrossRef]
- Vu, M.; Kassouf, N.; Ofili, R.; Lund, T.; Bell, C.; Appiah, S. Doxorubicin Selectively Induces Apoptosis through the Inhibition of a Novel Isoform of Bcl-2 in Acute Myeloid Leukaemia MOLM-13 Cells with Reduced Beclin 1 Expression. Int. J. Oncol. 2020, 57, 113–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Qare, A.W.; Abou-Donia, M.B. Biomarkers of Apoptosis: Release of Cytochrome c, Activation of Caspase-3, Induction of 8-Hydroxy-2′-Deoxyguanosine, Increased 3-Nitrotyrosine, and Alteration of P53 Gene. J. Toxicol. Environ. Health B Crit. Rev. 2001, 4, 113–121. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, D.A.; Macey, M.G.; Allen, P.D. A Simple, Novel, Procedure for Monitoring Membrane Scrambling and Permeability in Microparticles, Platelets, and Leukocytes in Whole Blood Samples. Exp. Hematol. 2008, 36, 909–921. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; Elbadawi, M.; Efferth, T. Multiple Cell Death Modalities and Their Key Features (Review). World Acad. Sci. J. 2020, 2, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Sarmiento-Salinas, F.L.; Delgado-Magallón, A.; Montes-Alvarado, J.B.; Ramírez-Ramírez, D.; Flores-Alonso, J.C.; Cortés-Hernández, P.; Reyes-Leyva, J.; Herrera-Camacho, I.; Anaya-Ruiz, M.; Pelayo, R.; et al. Breast Cancer Subtypes Present a Differential Production of Reactive Oxygen Species (ROS) and Susceptibility to Antioxidant Treatment. Front. Oncol. 2019, 9, 480. [Google Scholar] [CrossRef] [Green Version]
- Rodic, S.; Vincent, M.D. Reactive Oxygen Species (ROS) Are a Key Determinant of Cancer’s Metabolic Phenotype. Int. J. Cancer 2018, 142, 440–448. [Google Scholar] [CrossRef] [Green Version]
- Szakács, G.; Annereau, J.P.; Lababidi, S.; Shankavaram, U.; Arciello, A.; Bussey, K.J.; Reinhold, W.; Guo, Y.; Kruh, G.D.; Reimers, M.; et al. Predicting Drug Sensitivity and Resistance: Profiling ABC Transporter Genes in Cancer Cells. Cancer Cell 2004, 6, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Plaetzer, K.; Kiesslich, T.; Verwanger, T.; Krammer, B. The Modes of Cell Death Induced by PDT: An Overview. Med. Laser Appl. 2003, 18, 7–19. [Google Scholar] [CrossRef]
- Fleury, C.; Mignotte, B.; Vayssière, J.L. Mitochondrial Reactive Oxygen Species in Cell Death Signaling. Biochimie 2002, 84, 131–141. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 ± SEM (μM) | ||
|---|---|---|
| Cell Line | DARK | LIGHT |
| Jurkat | 25.52 ± 9.11 | 6.12 ± 1.39 |
| A-431 | 16.75 ± 2.78 | 1.70 ± 0.27 |
| MCF-7 | 80.11 ± 32.42 | 17.05 ± 4.93 |
| MDA-MB-231 | 161.13 ± 55.12 | 22.23 ± 2.46 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Greco, G.; Ulfo, L.; Turrini, E.; Marconi, A.; Costantini, P.E.; Marforio, T.D.; Mattioli, E.J.; Di Giosia, M.; Danielli, A.; Fimognari, C.; et al. Light-Enhanced Cytotoxicity of Doxorubicin by Photoactivation. Cells 2023, 12, 392. https://doi.org/10.3390/cells12030392
Greco G, Ulfo L, Turrini E, Marconi A, Costantini PE, Marforio TD, Mattioli EJ, Di Giosia M, Danielli A, Fimognari C, et al. Light-Enhanced Cytotoxicity of Doxorubicin by Photoactivation. Cells. 2023; 12(3):392. https://doi.org/10.3390/cells12030392
Chicago/Turabian StyleGreco, Giulia, Luca Ulfo, Eleonora Turrini, Alessia Marconi, Paolo Emidio Costantini, Tainah Dorina Marforio, Edoardo Jun Mattioli, Matteo Di Giosia, Alberto Danielli, Carmela Fimognari, and et al. 2023. "Light-Enhanced Cytotoxicity of Doxorubicin by Photoactivation" Cells 12, no. 3: 392. https://doi.org/10.3390/cells12030392