
Emerging Role of the Mast Cell–Microbiota Crosstalk in Cutaneous Homeostasis and Immunity

Abstract
:1. Introduction
2. Skin Innate Immunity
3. Mast Cells in the Skin—A Brief Overview
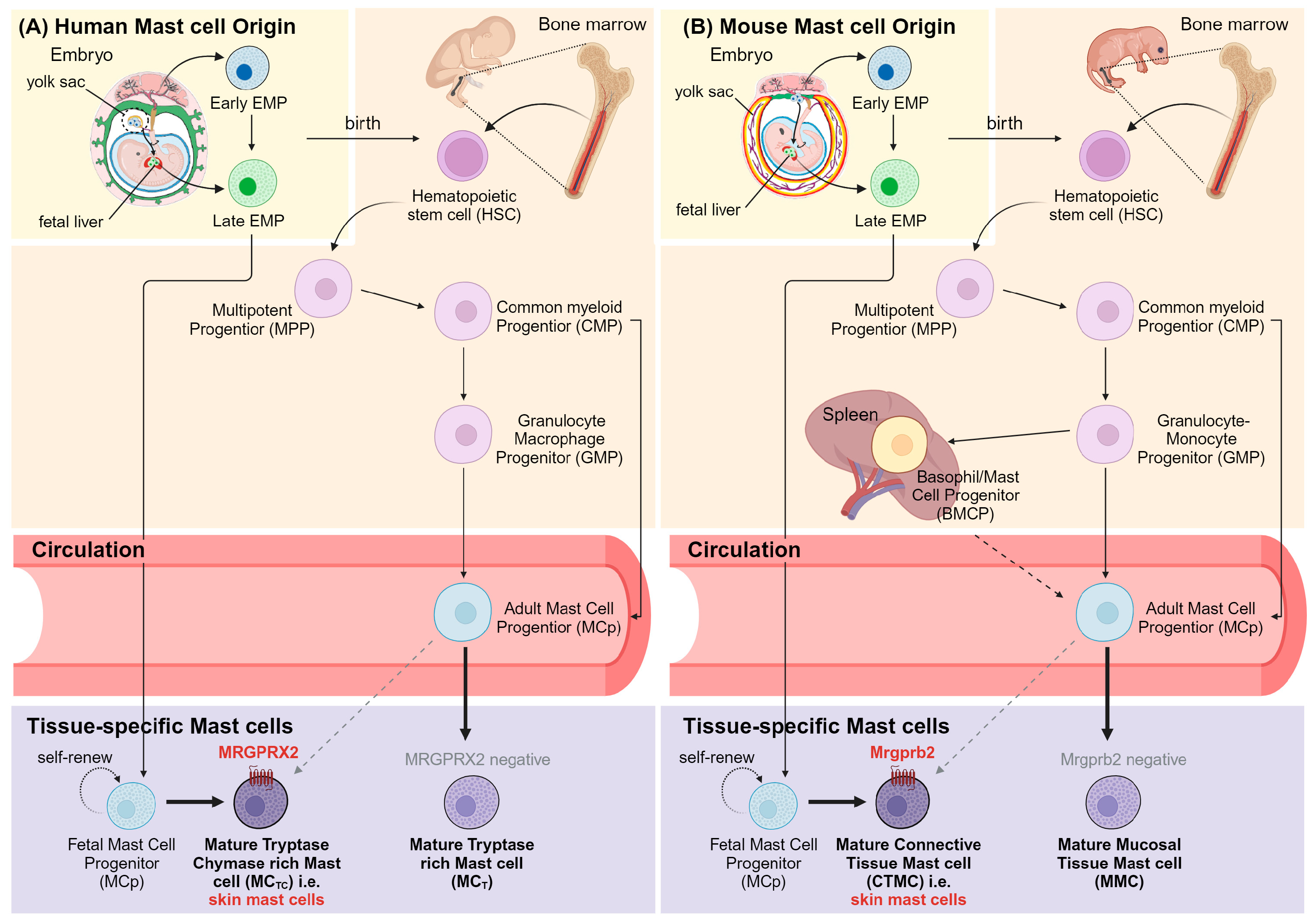
3.1. Mast Cell Subtypes and Development
3.2. Anatomic Location of Mast Cells in the Skin
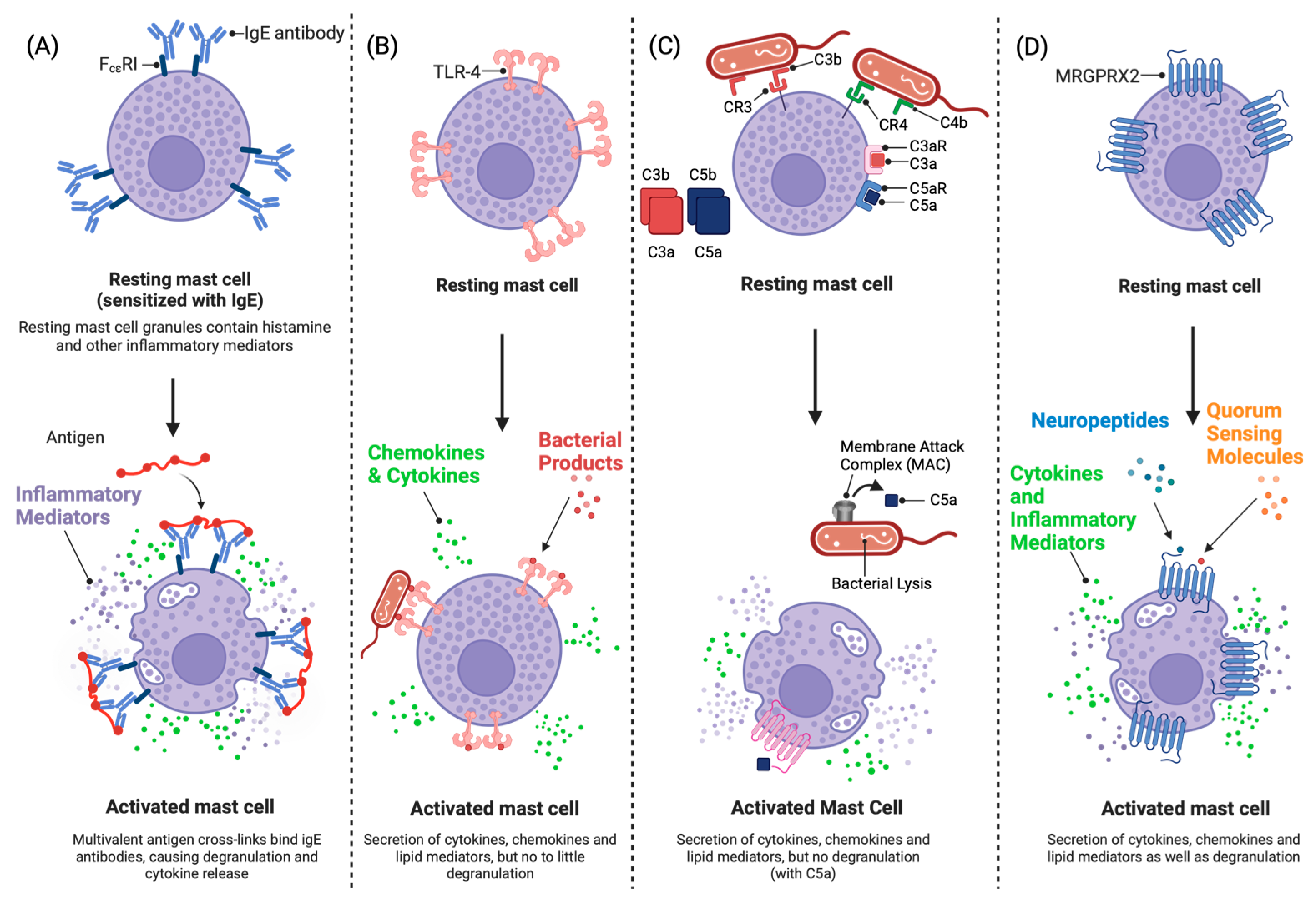
3.3. Phenotypic Characteristics of Skin Mast Cells
4. Cutaneous Microbial Diversity—Healthy vs. Inflamed Skin
4.1. The Cutaneous Microbiome in Health
4.2. The Cutaneous Microbiome in Disease
4.3. Mast Cells Interact with Commensal Bacteria
5. The Role of the Skin Microbiome in Mast Cell Development
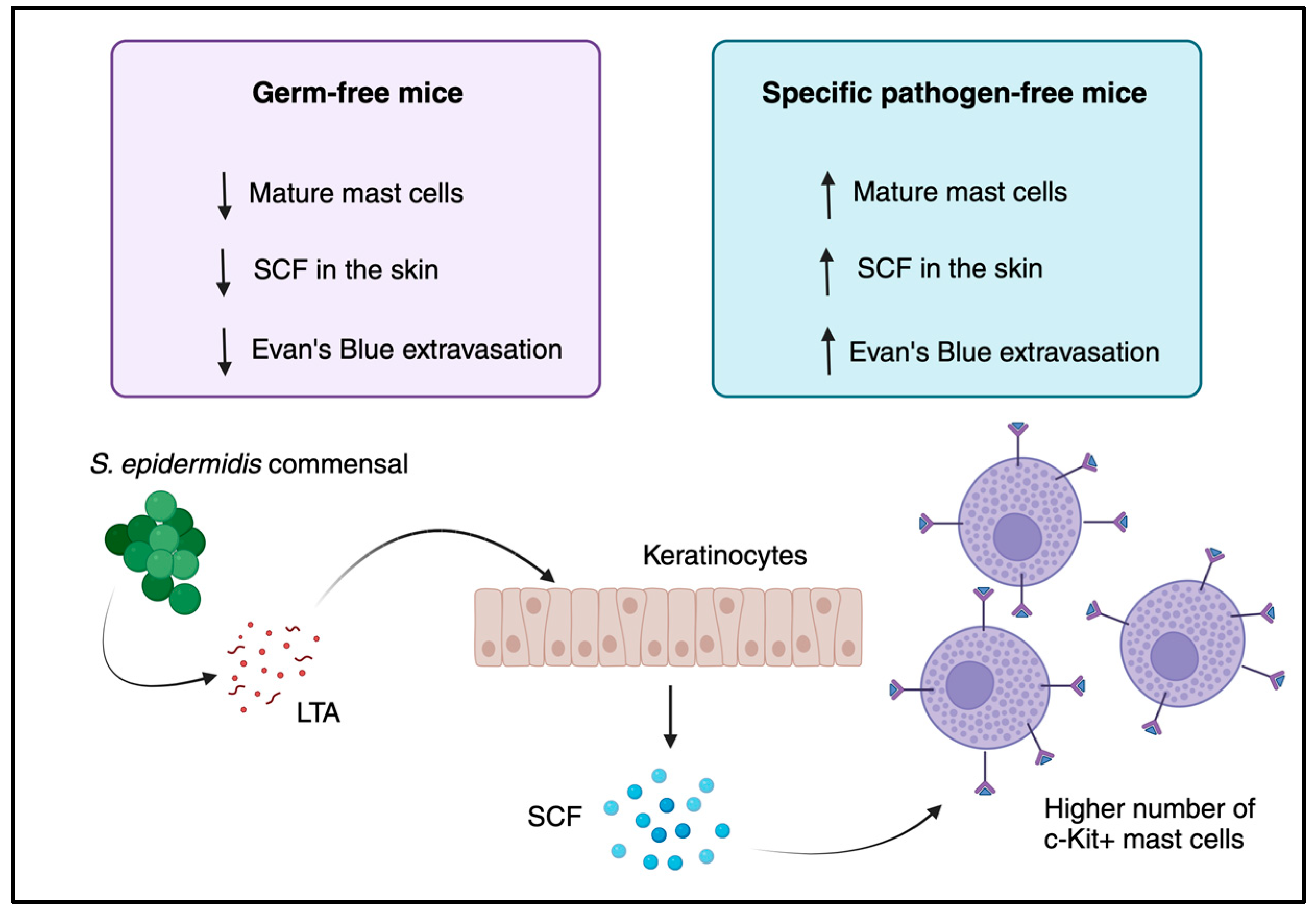
6. Mast Cells in Skin Barrier Function—Germ-Free vs. Conventional Mice
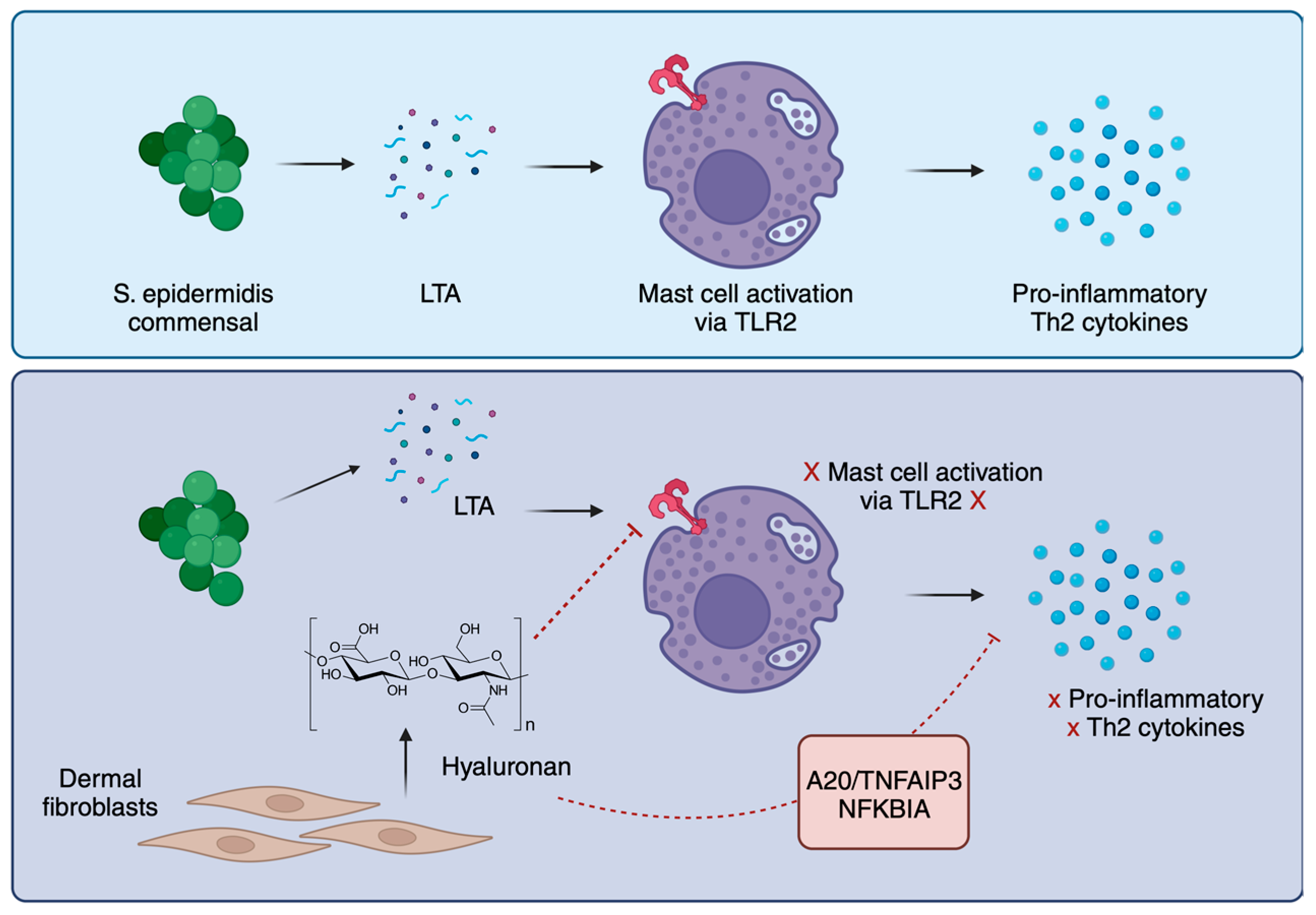
7. Mast Cell Tolerance to Commensal Bacteria
8. Commensal Bacteria Prime Mast Cells against Pathogens
9. Intra- and Interspecies Communication of Bacteria Influences Mast-Cell-Mediated Cutaneous Inflammation
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kobayashi, T.; Imanishi, I. Epithelial-immune crosstalk with the skin microbiota in homeostasis and atopic dermatitis—A mini review. Vet. Dermatol. 2021, 32, 533-e147. [Google Scholar] [CrossRef] [PubMed]
- Salmon, J.K.; Armstrong, C.A.; Ansel, J.C. The skin as an immune organ. West. J. Med. 1994, 160, 146–152. [Google Scholar] [PubMed]
- Kabashima, K.; Honda, T.; Ginhoux, F.; Egawa, G. The immunological anatomy of the skin. Nat. Rev. Immunol. 2019, 19, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Voss, M.; Kotrba, J.; Gaffal, E.; Katsoulis-Dimitriou, K.; Dudeck, A. Mast cells in the skin: Defenders of integrity or offenders in Inflammation? Int. J. Mol. Sci. 2021, 22, 4589. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Segre, J.A. Dialogue between skin microbiota and immunity. Science 2014, 346, 954–959. [Google Scholar] [CrossRef] [PubMed]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Dwyer, D.F.; Barrett, N.A.; Austen, K.F. Expression profiling of constitutive mast cells reveals a unique identity within the immune system. Nat. Immunol. 2016, 17, 878–887. [Google Scholar] [CrossRef]
- Quaresma, J.A.S. Organization of the skin immune system and compartmentalized immune responses in infectious diseases. Clin. Microbiol. Rev. 2019, 32, e00034‐18. [Google Scholar] [CrossRef]
- Pasparakis, M.; Haase, I.; Nestle, F.O. Mechanisms regulating skin immunity and inflammation. Nat. Rev. Immunol. 2014, 14, 289–301. [Google Scholar] [CrossRef]
- Zhang, C.; Merana, G.R.; Harris-Tryon, T.; Scharschmidt, T.C. Skin immunity: Dissecting the complex biology of our body’s outer barrier. Mucosal Immunol. 2022, 15, 551–561. [Google Scholar] [CrossRef]
- Komi, D.E.A.; Khomtchouk, K.; Santa Maria, P.L. A review of the contribution of mast cells in wound healing: Involved molecular and cellular mechanisms. Clin. Rev. Allergy Immunol. 2020, 58, 298–312. [Google Scholar] [CrossRef] [PubMed]
- Pundir, P.; Kulka, M. The role of G protein-coupled receptors in mast cell activation by antimicrobial peptides: Is there a connection? Immunol. Cell Biol. 2010, 88, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Pundir, P.; Catalli, A.; Leggiadro, C.; Douglas, S.E.; Kulka, M. Pleurocidin, a novel antimicrobial peptide, induces human mast cell activation through the FPRL1 receptor. Mucosal Immunol. 2014, 7, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Pundir, P.; MacDonald, C.A.; Kulka, M. The novel receptor C5aR2 is required for c5a-mediated human mast cell adhesion, migration, and proinflammatory mediator production. J. Immunol. 2015, 195, 2774–2787. [Google Scholar] [CrossRef]
- Tauber, M.; Basso, L.; Martin, J.; Bostan, L.; Pinto, M.M.; Thierry, G.R.; Houmadi, R.; Serhan, N.; Loste, A.; Bleriot, C.; et al. Landscape of mast cell populations across organs in mice and humans. J. Exp. Med. 2023, 220, e20230570. [Google Scholar] [CrossRef]
- Gentek, R.; Ghigo, C.; Hoeffel, G.; Bulle, M.J.; Msallam, R.; Gautier, G.; Launay, P.; Chen, J.; Ginhoux, F.; Bajénoff, M. Hemogenic endothelial fate mapping reveals dual developmental origin of mast cells. Immunity 2018, 48, 1160–1171.e5. [Google Scholar] [CrossRef]
- Gurish, M.F.; Austen, K.F. Developmental origin and functional specialization of mast cell subsets. Immunity 2012, 37, 25–33. [Google Scholar] [CrossRef]
- Li, Z.; Liu, S.; Xu, J.; Zhang, X.; Han, D.; Liu, J.; Xia, M.; Yi, L.; Shen, Q.; Xu, S.; et al. Adult connective tissue-resident mast cells originate from late erythro-myeloid progenitors. Immunity 2018, 49, 640–653.e5. [Google Scholar] [CrossRef]
- St John, A.L.; Rathore, A.P.S.; Ginhoux, F. New perspectives on the origins and heterogeneity of mast cells. Nat. Rev. Immunol. 2023, 23, 55–68. [Google Scholar] [CrossRef]
- Kitamura, Y.; Oboki, K.; Ito, A. Development of mast cells. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2007, 83, 164–174. [Google Scholar] [CrossRef]
- Li, Y.; Liu, S.; Xu, J.; Zhang, X.; Han, D.; Liu, J.; Xia, M.; Yi, L.; Shen, Q.; Xu, S.; et al. Mast cells and acupuncture Analgesia. Cells 2022, 11, 860. [Google Scholar] [CrossRef] [PubMed]
- Jamur, M.C.; Oliver, C. Origin, maturation and recruitment of mast cell precursors. Front. Biosci. (Schol. Ed.) 2011, 3, 1390–1406. [Google Scholar] [PubMed]
- Toyoshima, S.; Okayama, Y. Neuro-allergology: Mast cell-nerve cross-talk. Allergol. Int. 2022, 71, 288–293. [Google Scholar] [CrossRef]
- Ashina, K.; Tsubosaka, Y.; Nakamura, T.; Omori, K.; Kobayashi, K.; Hori, M.; Ozaki, H.; Murata, T. Histamine induces vascular hyperpermeability by increasing blood flow and endothelial barrier disruption in vivo. PLoS ONE 2015, 10, e0132367. [Google Scholar] [CrossRef] [PubMed]
- Marone, G.; Varricchi, G.; Loffredo, S.; Granata, F. Mast cells and basophils in inflammatory and tumor angiogenesis and lymphangiogenesis. Eur. J. Pharmacol. 2016, 778, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Tsilioni, I.; Ren, H. Recent advances in our understanding of mast cell activation or should it be mast cell mediator disorders? Expert Rev. Clin. Immunol. 2019, 15, 639–656. [Google Scholar] [CrossRef]
- Cheng, L.E.; Hartmann, K.; Roers, A.; Krummel, M.F.; Locksley, R.M. Perivascular mast cells dynamically probe cutaneous blood vessels to capture IgE. Immunity 2013, 38, 166–175. [Google Scholar] [CrossRef]
- Sumpter, T.L.; Balmert, S.C.; Kaplan, D.H. Cutaneous immune responses mediated by dendritic cells and mast cells. JCI Insight 2019, 4, e123947. [Google Scholar] [CrossRef]
- Ito, T.; Smrz, D.; Jung, M.Y.; Bandara, G.; Desai, A.; Smrzová, S.; Kuehn, H.S.; Beaven, M.A.; Metcalfe, D.D.; Gilfillan, A.M. Stem cell factor programs the mast cell activation phenotype. J. Immunol. 2012, 188, 5428–5437. [Google Scholar] [CrossRef]
- Tsai, M.; Takeishi, T.; Thomposn, H.; Langley, K.E.; Zsebo, K.M.; Metcalfe, D.D.; Geissler, E.N.; Galli, S.J. Induction of mast cell proliferation, maturation, and heparin synthesis by the rat c-kit ligand, stem cell factor. Proc. Natl. Acad. Sci. USA 1991, 88, 6382–6386. [Google Scholar] [CrossRef]
- Artuc, M.; Steckelings, U.M.; Henz, B.M. Mast cell-fibroblast interactions: Human mast cells as source and inducers of fibroblast and epithelial growth factors. J. Investig. Dermatol. 2002, 118, 391–395. [Google Scholar] [CrossRef]
- Pincha, N.; Hajam, E.Y.; Badrinath, K.; Batta, S.P.R.; Masudi, T.; Dey, R.; Andreasen, P.; Kawakami, T.; Samuel, R.; George, R.; et al. PAI1 mediates fibroblast-mast cell interactions in skin fibrosis. J. Clin. Investig. 2018, 128, 1807–1819. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Murota, H.; Serada, S.; Fujimoto, M.; Kudo, A.; Naka, T.; Katayama, I. Histamine contributes to tissue remodeling via periostin expression. J. Investig. Dermatol. 2014, 134, 2105–2113. [Google Scholar] [CrossRef] [PubMed]
- Halaban, R.; Kwon, B.S.; Ghosh, S.; Delli Bovi, P.; Baird, A. bFGF as an autocrine growth factor for human melanomas. Oncogene Res. 1988, 3, 177–186. [Google Scholar] [PubMed]
- Schadendorf, D.; Moller, A.; Algermissen, B.; Worm, M.; Sticherling, M.; Czarnetzki, B.M. Il-8 produced by human malignant melanoma cells in vitro is an essential autocrine growth factor. J. Immunol. 1993, 151, 2667–2675. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Barrera, R.; Torres-Alvarez, B.; Castanedo-Cazares, J.P.; Oros-Ovalle, C.; Moncada, B. Solar elastosis and presence of mast cells as key features in the pathogenesis of melasma. Clin. Exp. Dermatol. 2008, 33, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Tomita, Y.; Maeda, K.; Tagami, H. Histamine stimulates normal human melanocytes in vitro: One of the possible inducers of hyperpigmentation in urticaria pigmentosa. J. Dermatol. Sci. 1993, 6, 146–154. [Google Scholar] [CrossRef]
- Yoshida, M.; Takahashi, Y.; Inoue, S. Histamine induces melanogenesis and morphologic changes by protein kinase a activation via H2 receptors in human normal melanocytes. J. Investig. Dermatol. 2000, 114, 334–342. [Google Scholar] [CrossRef]
- Wernersson, S.; Pejler, G. Mast cell secretory granules: Armed for battle. Nat. Rev. Immunol. 2014, 14, 478–494. [Google Scholar] [CrossRef]
- Komi, D.E.A.; Wöhrl, S.; Bielory, L. Mast cell biology at molecular level: A comprehensive review. Clin. Rev. Allergy Immunol. 2020, 58, 342–365. [Google Scholar] [CrossRef]
- McNeil, B.D.; Pundir, P.; Meekar, S.; Han, L.; Undem, B.J.; Kulka, M.; Dong, X. Identification of a mast-cell-specific receptor crucial for pseudo-allergic drug reactions. Nature 2015, 519, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Xing, W.; Austen, K.F.; Gurish, M.F.; Jones, T.G. Protease phenotype of constitutive connective tissue and of induced mucosal mast cells in mice is regulated by the tissue. Proc. Natl. Acad. Sci. USA 2011, 108, 14210–14215. [Google Scholar] [CrossRef] [PubMed]
- Motakis, E.; Guhl, S.; Ishizu, Y.; Itoh, M.; Kawaji, H.; Hoon, M.D.; Lassman, T.; Carninci, P.; Hayaschizaki, Y.; Zuberbier, T.; et al. Redefinition of the human mast cell transcriptome by deep-CAGE sequencing. Blood 2014, 123, e58–e67. [Google Scholar] [CrossRef] [PubMed]
- Green, D.P.; Limjunyawong, N.; Gour, N.; Pundir, P.; Dong, X. A mast-cell-specific receptor mediates neurogenic inflammation and pain. Neuron 2019, 101, 412–420.e3. [Google Scholar] [CrossRef]
- Pundir, P.; Liu, R.; Vasvada, C.; Serhan, N.; Limjunyawong, N.; Yee, R.; Zhan, Y.; Dong, X.; Wu, X.; Zhang, Y.; et al. A connective tissue mast-cell-specific receptor detects bacterial quorum-sensing molecules and mediates antibacterial immunity. Cell Host Microbe 2019, 26, 114–122.e8. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, R.; Che, D.; Pundir, P.; Wang, N.; Han, S.; Cao, J.; Lv, Y.; Dong, H.; Fang, F.; et al. A mast cell-specific receptor is critical for granuloma induced by intrathecal morphine infusion. J. Immunol. 2019, 203, 1701–1714. [Google Scholar] [CrossRef]
- Liu, R.; Che, D.; Zhao, T.; Pundir, P.; Cao, J.; Lv, Y.; Wang, J.; Ma, P.; Fu, J.; Wang, N.; et al. MRGPRX2 is essential for sinomenine hydrochloride induced anaphylactoid reactions. Biochem. Pharmacol. 2017, 146, 214–223. [Google Scholar] [CrossRef]
- Zhang, T.; Che, D.; Liu, R.; Han, S.; Wang, N.; Zhan, Y.; Pundir, P.; Cao, J.; Lv, Y.; Wang, L.; et al. Typical antimicrobials induce mast cell degranulation and anaphylactoid reactions via MRGPRX2 and its murine homologue MRGPRB2. Eur. J. Immunol. 2017, 47, 1949–1958. [Google Scholar] [CrossRef]
- Grice, E.A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; NISC Comparative Sequencing Program; Bouffard, G.G.; Blakesley, R.W.; Muray, P.R.; et al. Topographical and temporal diversity of the human skin microbiome. Science 2009, 324, 1190–1192. [Google Scholar] [CrossRef]
- Grice, E.A.; Segre, J.A. The skin microbiome. Nat. Rev. Microbiol. 2011, 9, 244–253. [Google Scholar] [CrossRef]
- Oh, J.; Byrd, A.L.; Park, M.; NISC Comparative Sequencing Program; Kong, H.H.; Segre, J.A. Temporal stability of the human skin microbiome. Cell 2016, 165, 854–866. [Google Scholar] [CrossRef] [PubMed]
- Findley, K.; Oh, J.; Yang, J.; Conlan, S.; Deming, C.; Meyer, J.A.; Schoenfeld, D.; Nomicos, E.; Park, M.; NIH Intramural Sequencing Center Comparative Sequencing Program; et al. Topographic diversity of fungal and bacterial communities in human skin. Nature 2013, 498, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Koh, L.F.; Ong, R.Y.; Common, J.E. Skin microbiome of atopic dermatitis. Allergol. Int. 2022, 71, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, S.; Iwamoto, T.; Takada, K.; Okuda, K.I.; Tajima, A.; Iwase, T.; Mizunoe, Y. Staphylococcus epidermidis esp degrades specific proteins associated with staphylococcus aureus biofilm formation and host-pathogen interaction. J. Bacteriol. 2013, 195, 1645–1655. [Google Scholar] [CrossRef]
- Naik, S.; Bouladoux, N.; Wilhelm, C.; Molloy, M.J.; Salcedo, R.; Kastenmuller, W.; Deming, C.; Quinones, M.; Koo, L.; Conlan, S.; et al. Compartmentalized control of skin immunity by resident commensals. Science 2012, 337, 1115–1119. [Google Scholar] [CrossRef]
- Lai, Y.; Di Nardo, A.; Nakatsuji, T.; Leichtle, A.; Yang, Y.; Cogen, A.L.; Wu, Z.R.; Hooper, L.V.; Aulock, S.V.; Radek, K.A.; et al. Commensal bacteria regulate TLR3-dependent inflammation following skin injury. Nat. Med. 2009, 15, 1377–1382. [Google Scholar] [CrossRef]
- Kong, H.H.; Oh, J.; Deming, C.; Conlan, S.; Grice, E.A.; Beatson, M.A.; Nomicos, E.; Polley, E.C.; Komarow, H.D.; NISC Comparative Sequencing Program; et al. Temporal shifts in the skin microbiome associated with disease flares and treatment in children with atopic dermatitis. Genome Res. 2012, 22, 850–859. [Google Scholar] [CrossRef]
- Lewis, D.J.; Chan, W.H.; Hinojosa, T.; Hsu, S.; Feldman, S.R. Mechanisms of microbial pathogenesis and the role of the skin microbiome in psoriasis: A review. Clin. Dermatol. 2019, 37, 160–166. [Google Scholar] [CrossRef]
- Marshall, J.S. Mast-cell responses to pathogens. Nat. Rev. Immunol. 2004, 4, 787–799. [Google Scholar] [CrossRef]
- Yu, Y.; Blockhuis, B.R.; Garssen, J.; Redegeld, F.A. Non-IgE mediated mast cell activation. Eur. J. Pharmacol. 2016, 778, 33–43. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Chiang, H.I.; Jiang, S.B.; Nagarajan, H.; Zengler, K.; Gallo, R.L. The microbiome extends to subepidermal compartments of normal skin. Nat. Commun. 2013, 4, 1431. [Google Scholar] [CrossRef] [PubMed]
- Bay, L.; Barnes, C.J.; Fritz, B.G.; Thorsen, J.; Restrup, M.E.M.; Rasmussen, L.; Sorenson, J.K.; Hesselvig, A.B.; Odgaard, A.; Hansen, A.J.; et al. Universal dermal microbiome in human skin. MBio 2020, 11, e02945‐19. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Mascarenhas, N.; Eckmann, L.; Miyamoto, Y.; Sun, X.; Kawakami, T.; Nardo, A.N. Skin microbiome promotes mast cell maturation by triggering stem cell factor (SCF) production in keratinocytes. J. Allergy Clin. Immunol. 2017, 139, 1205–1216.e6. [Google Scholar] [CrossRef] [PubMed]
- Kunisada, T.; Lu, S.Z.; Yoshida, H.; Nishikawa, S.; Nishikawa, S.; Mizoguchi, M.; Tyrrell, L.; Willaims, D.A.; Wang, X.; Longley, B.J. Murine Cutaneous mastocytosis and epidermal melanocytosis induced by keratinocyte expression of transgenic stem cell factor. J. Exp. Med. 1998, 187, 1565–1573. [Google Scholar] [CrossRef]
- Huttunen, M.; Aalto, M.L.; Harvima, R.J.; Hormanheimo, M.; Harvima, I.T. Alterations in mast cells showing tryptase and chymase activity in epithelializating and chronic wounds: Mast cells and tryptase & chymase activity in chronic wounds. Exp. Dermatol. 2000, 9, 258–265. [Google Scholar]
- Canesso, M.C.C.; Vieira, A.T.; Castro, T.B.R.; Schirmer, B.G.A.; Cisalpino, D.; Martins, F.S.; Rachid, M.A.; Nicoli, J.R.; Teixeira, M.M.; Barcelos, L.S. Skin wound healing is accelerated and scarless in the absence of commensal microbiota. J. Immunol. 2014, 193, 5171–5180. [Google Scholar] [CrossRef]
- Ng, M.F.Y. The role of mast cells in wound healing. Int. Wound J. 2010, 7, 55–61. [Google Scholar] [CrossRef]
- Weller, K.; Foitzik, K.; Paus, R.; Syska, W.; Maurer, M. Mast cells are required for normal healing of skin wounds in mice. FASEB J. 2006, 20, 2366–2368. [Google Scholar] [CrossRef]
- Di Nardo, A.; Chang, Y.L.; Alimohammadi, S.; Masuda-Kuroki, K.; Wang, Z.; Sriram, K.; Insel, P.A. Mast cell tolerance in the skin microenvironment to commensal bacteria is controlled by fibroblasts. Cell Rep. 2023, 42, 112453. [Google Scholar] [CrossRef]
- Cayrol, C.; Girard, J.P. Interleukin-33 (IL-33): A critical review of its biology and the mechanisms involved in its release as a potent extracellular cytokine. Cytokine 2022, 156, 155891. [Google Scholar] [CrossRef]
- Sandig, H.; Jobbings, C.E.; Roldan, N.G.; Whittingham-Dowd, J.K.; Orinska, Z.; Takeuchi, O.; Akira, S.; Bulfone-Paus, S. IL-33 causes selective mast cell tolerance to bacterial cell wall products by inducing IRAK1 degradation. Eur. J. Immunol. 2013, 43, 979–988. [Google Scholar] [CrossRef] [PubMed]
- Magerl, M.; Lammel, V.; Siebenhaar, F.; Zuberbier, T.; Metz, M.; Maurer, M. Non-pathogenic commensal Escherichia coli bacteria can inhibit degranulation of mast cells. Exp. Dermatol. 2008, 17, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.W.; Abraham, S.N. Mast cell mediator responses and their suppression by pathogenic and commensal microorganisms. Mol. Immunol. 2015, 63, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Forsythe, P. Microbes taming mast cells: Implications for allergic inflammation and beyond. Eur. J. Pharmacol. 2016, 778, 169–175. [Google Scholar] [CrossRef]
- Kulka, M.; Fukushi, N.; Rottem, M.; Mekori, Y.A.; Metcalfe, D.D. Mast cells, which interact with Escherichia coli, up-regulate genes associated with innate immunity and become less responsive to Fc(epsilon)RI-mediated activation. J. Leukoc. Biol. 2006, 79, 339–350. [Google Scholar] [CrossRef]
- Kawahara, T. Inhibitory effect of heat-killed Lactobacillus strain on immunoglobulin E-mediated degranulation and late-phase immune reactions of mouse bone marrow-derived mast cells. Anim. Sci. J. 2010, 81, 714–721. [Google Scholar] [CrossRef]
- Forsythe, P.; Wang, B.; Khambati, I.; Kunze, W.A. Systemic effects of ingested Lactobacillus rhamnosus: Inhibition of mast cell membrane potassium (IKCa) current and degranulation. PLoS ONE 2012, 7, e41234. [Google Scholar] [CrossRef]
- Oksaharju, A.; Kankainen, M.; Kekkonen, R.A.; Lindstedt, K.A.; Kovanen, P.T.; Korpela, R.; Miettinen, M. Probiotic Lactobacillus rhamnosus downregulates FCER1 and HRH4 expression in human mast cells. World J. Gastroenterol. 2011, 17, 750–759. [Google Scholar] [CrossRef]
- Schiffer, C.; Lalanne, A.I.; Cssard, L.; Mancardi, D.A.; Malbec, O.; Bruhns, P.; Dif, F.; Daeron, M. A strain of Lactobacillus casei inhibits the effector phase of immune inflammation. J. Immunol. 2011, 187, 2646–2655. [Google Scholar] [CrossRef]
- Harata, G.; He, F.; Takahashi, K.; Hosono, A.; Kawase, M.; Kubota, A.; Hiramatsu, M.; Kaminogawa, S. Bifidobacterium suppresses IgE-mediated degranulation of rat basophilic leukemia (RBL-2H3) cells. Microbiol. Immunol. 2010, 54, 54–57. [Google Scholar] [CrossRef]
- Inoue, R.; Nishio, A.; Fukushima, Y.; Ushida, K. Oral treatment with probiotic Lactobacillus johnsonii NCC533 (La1) for a specific part of the weaning period prevents the development of atopic dermatitis induced after maturation in model mice, NC/Nga. Br. J. Dermatol. 2007, 156, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Hacini-Rachinel, F.; Gheit, H.; Le Ludec, J.B.; Dif, F.; Nancey, S.; Kaiserlian, D. Oral probiotic control skin inflammation by acting on both effector and regulatory T cells. PLoS ONE 2009, 4, e4903. [Google Scholar] [CrossRef] [PubMed]
- Won, T.J.; Kim, B.; Lim, Y.T.; Song, D.S.; Park, S.-Y.; Park, E.S.; Lee, D.I.; Hwang, K.W. Oral administration of Lactobacillus strains from Kimchi inhibits atopic dermatitis in NC/Nga mice. J. Appl. Microbiol. 2011, 110, 1195–1202. [Google Scholar] [CrossRef] [PubMed]
- Sawada, J.; Morita, H.; Tanaka, A.; Salminen, S.; He, F.; Matsuda, H. Ingestion of heat-treated Lactobacillus rhamnosus GG prevents development of atopic dermatitis in NC/Nga mice. Clin. Exp. Allergy 2007, 37, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Qi, C.; Li, X.; Lu, M.; Zhang, H.; Zhou, J.; Dang, H.; Che, J.; Li, S.; Sun, J.; et al. Prevention of atopic dermatitis in mice by Lactobacillus reuteri fn041 through induction of regulatory t cells and modulation of the gut microbiota. Mol. Nutr. Food Res. 2022, 66, e2100699. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.S.; Lim, S.K.; Jang, J.-Y.; Lee, J.; Park, H.K.; Kim, N.; Yun, M.; Shin, M.-Y.; Jo, H.E.; Oh, Y.J.; et al. Lactobacillus sakei WIKIM30 ameliorates atopic dermatitis-like skin lesions by inducing regulatory T cells and altering gut microbiota structure in mice. Front. Immunol. 2018, 9, 1905. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.K.; Jang, Y.J.; Han, D.H.; Jeon, K.; Lee, C.; Han, H.S.; Ko, G. Lactobacillus paracasei KBL382 administration attenuates atopic dermatitis by modulating immune response and gut microbiota. Gut Microbes 2020, 12, 1819156. [Google Scholar] [CrossRef]
- de Andrade, P.; E Silva, J.M.; Carregaro, V.; Sacramento, L.A.; Roberti, L.R.; Aragon, D.C.; Carmona, F.; Roxo-Junior, P. Efficacy of probiotics in children and adolescents with atopic dermatitis: A randomized, double-blind, placebo-controlled study. Front. Nutr. 2021, 8, 833666. [Google Scholar] [CrossRef]
- Xie, A.; Chen, A.; Chen, Y.; Luo, Z.; Jiang, S.; Chen, D.; Yu, R. Lactobacillus for the treatment and prevention of atopic dermatitis: Clinical and experimental evidence. Front. Cell. Infect. Microbiol. 2023, 13, 1137275. [Google Scholar] [CrossRef]
- Wang, Z.; MacLeod, D.T.; Di Nardo, A. Commensal bacteria lipoteichoic acid increases skin mast cell antimicrobial activity against vaccinia viruses. J. Immunol. 2012, 189, 1551–1558. [Google Scholar] [CrossRef]
- Mukherjee, S.; Jemielita, M.; Stergioula, V.; Tikhonov, M.; Bassler, B.L. Photosensing and quorum sensing are integrated to control Pseudomonas aeruginosa collective behaviors. PLoS Biol. 2019, 17, e3000579. [Google Scholar] [CrossRef]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef]
- Rutherford, S.T.; Bassler, B.L. Bacterial quorum sensing: Its role in virulence and possibilities for its control. Cold Spring Harb. Perspect. Med. 2012, 2, a012427. [Google Scholar] [CrossRef]
- CorbiËre, A.; Loste, A.; Gaudenzio, N. MRGPRX2 sensing of cationic compounds—A bridge between nociception and skin diseases? Exp. Dermatol. 2021, 30, 193–200. [Google Scholar] [CrossRef]
- Monticelli, S.; Leoni, C. Epigenetic and transcriptional control of mast cell responses. F1000Research 2017, 6, 2064. [Google Scholar] [CrossRef] [PubMed]
- Medina-Tamayo, J.; Ibarra-Sanchez, A.; Padilla-Trejo, A.; Gonzalez-Espinosa, C. IgE-dependent sensitization increases responsiveness to LPS but does not modify development of endotoxin tolerance in mast cells. Inflamm. Res. 2011, 60, 19–27. [Google Scholar] [CrossRef] [PubMed]
- De Zuani, M.; Dal Secco, C.; Tonon, S.; Arzese, A.; Pucillo, C.E.M.; Frossi, B. LPS guides distinct patterns of training and tolerance in mast cells. Front. Immunol. 2022, 13, 835348. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
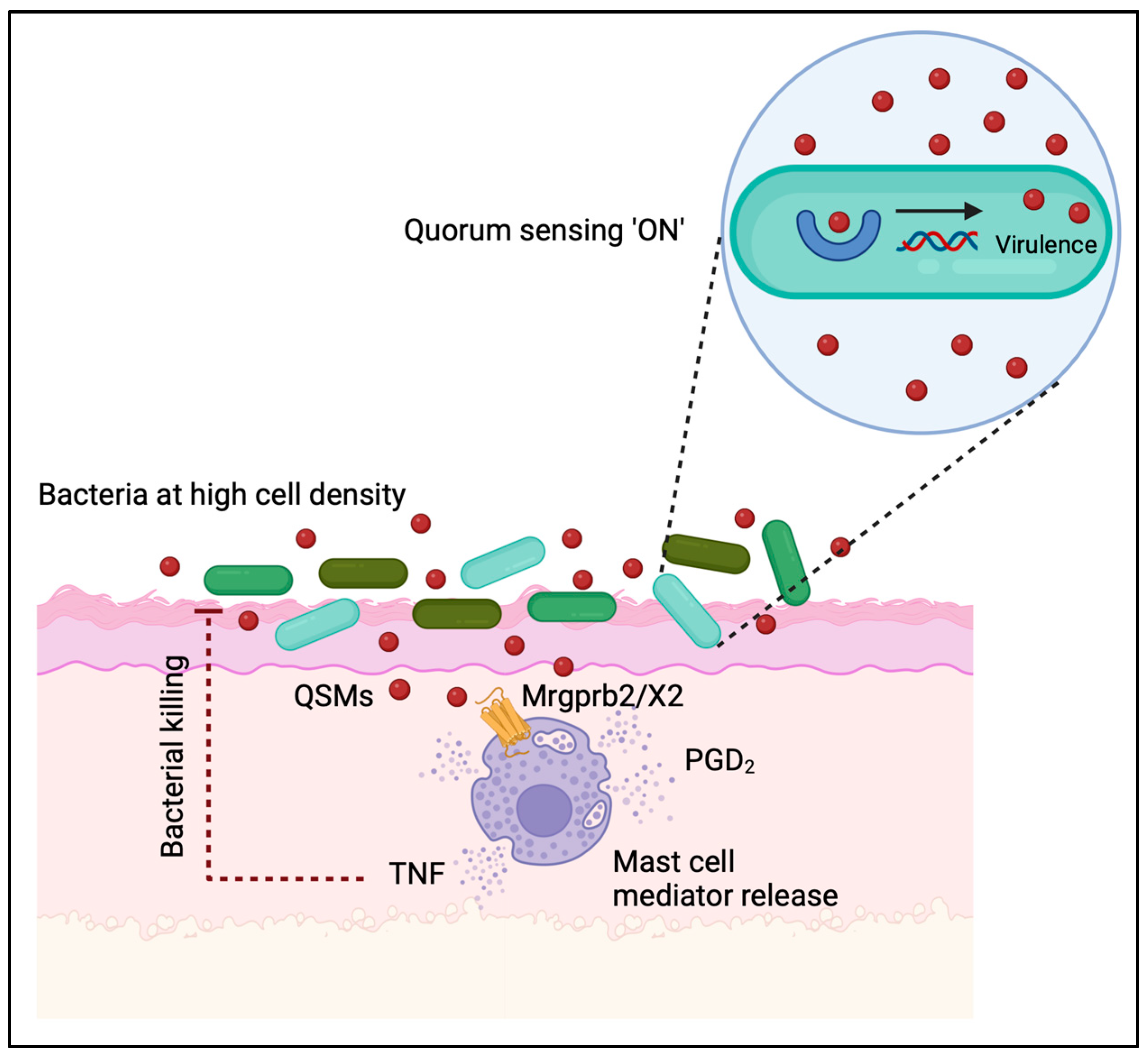{kind=link}
| Cytokine/Chemokine/Complement Receptors | MCTC (Connective) | MCT (Mucosal) |
|---|---|---|
| Tryptase | High | High |
| Chymase | High | Low |
| Heparin | Low | High |
| MrgprX2 | High | Negative |
| External TLR-1/-2/-4/-5/-6 | Low | High |
| Internal TLR-3/-8 | High | High |
| Internal TLR-7/-10 | Low | High |
| Internal TLR-9 | High | Negative |
| C3aR | High | Low |
| C5aR | High | Low/negative |
| Histamine receptor-1/-2 | High | Low |
| Histamine receptor-3/-4 | Low | High |
| CMA1, HEY1, and C5R1 | High | Negative |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bosveld, C.J.; Guth, C.; Limjunyawong, N.; Pundir, P. Emerging Role of the Mast Cell–Microbiota Crosstalk in Cutaneous Homeostasis and Immunity. Cells 2023, 12, 2624. https://doi.org/10.3390/cells12222624
Bosveld CJ, Guth C, Limjunyawong N, Pundir P. Emerging Role of the Mast Cell–Microbiota Crosstalk in Cutaneous Homeostasis and Immunity. Cells. 2023; 12(22):2624. https://doi.org/10.3390/cells12222624
Chicago/Turabian StyleBosveld, Cameron Jackson, Colin Guth, Nathachit Limjunyawong, and Priyanka Pundir. 2023. "Emerging Role of the Mast Cell–Microbiota Crosstalk in Cutaneous Homeostasis and Immunity" Cells 12, no. 22: 2624. https://doi.org/10.3390/cells12222624