
Adolescent Intermittent Ethanol Drives Modest Neuroinflammation but Does Not Escalate Drinking in Male Rats
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
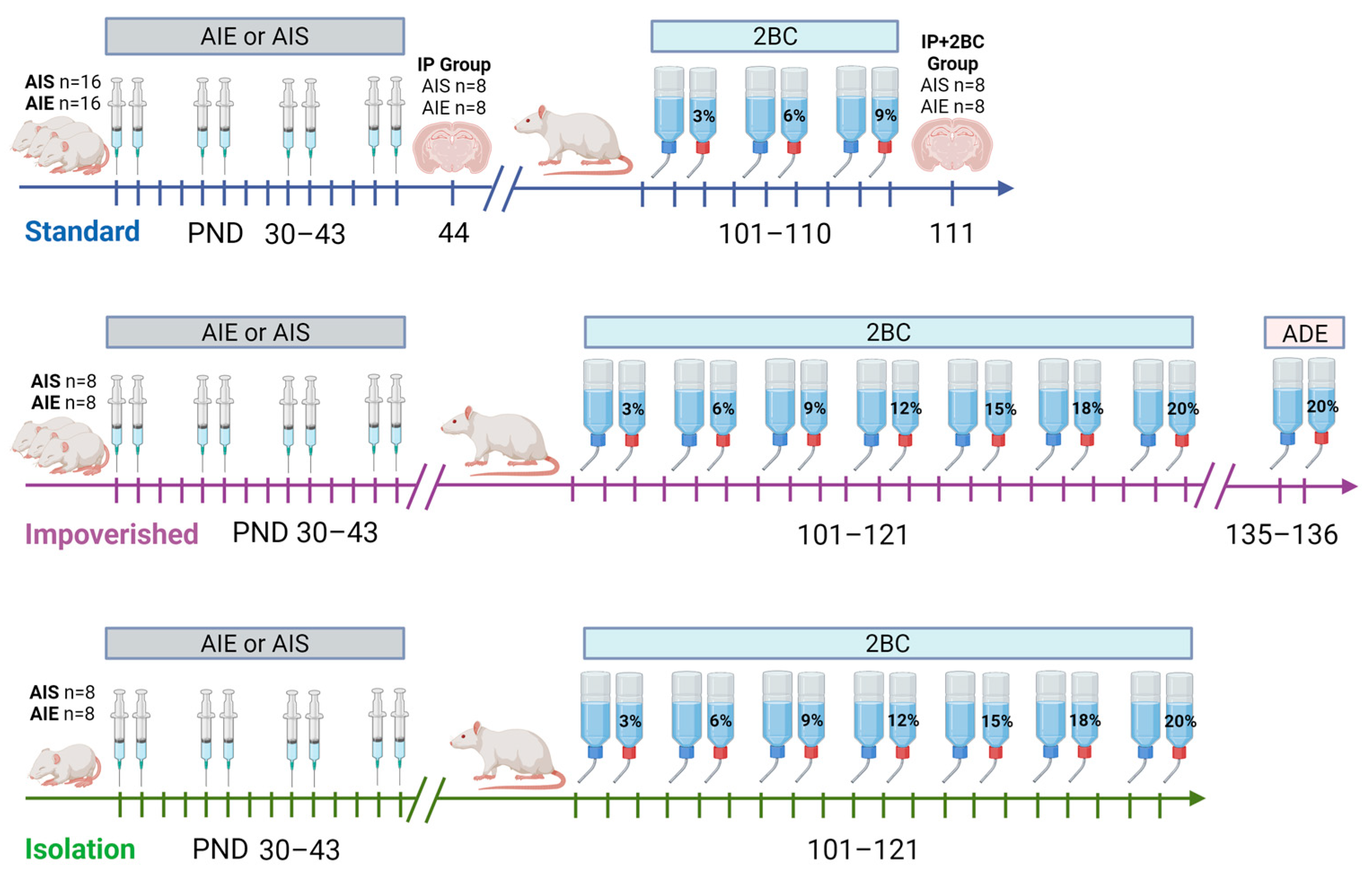
2.1. Animals and Housing Conditions
2.2. Adolescent Ethanol Exposure
2.3. Alcohol Drinking in Adulthood
2.4. Histology
2.5. Quantification
2.6. Statistical Approaches
3. Results
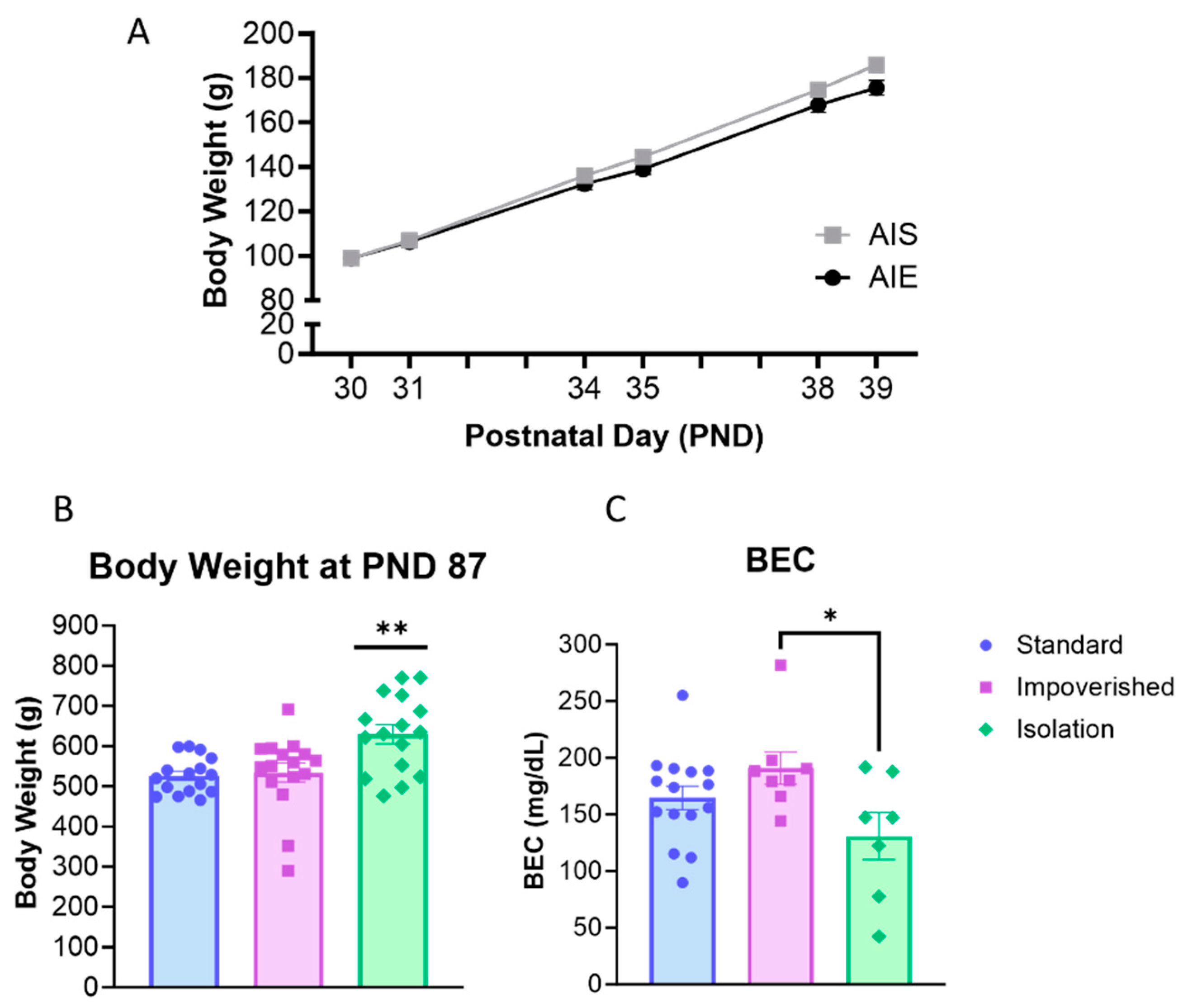
3.1. Social Isolation Increased Body Weight
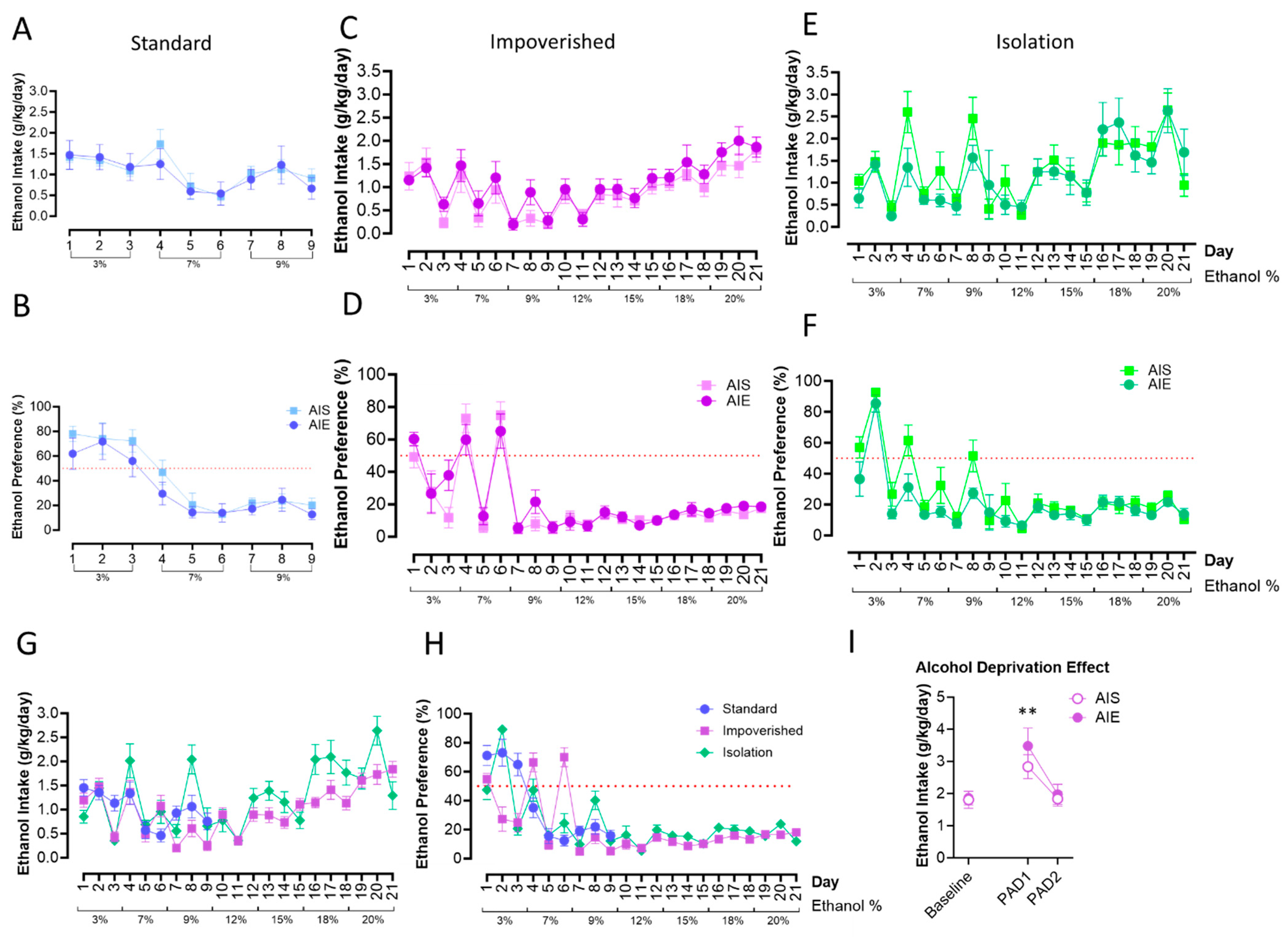
3.2. AIE Did Not Alter Adult Alcohol Drinking, with or without Social Isolation
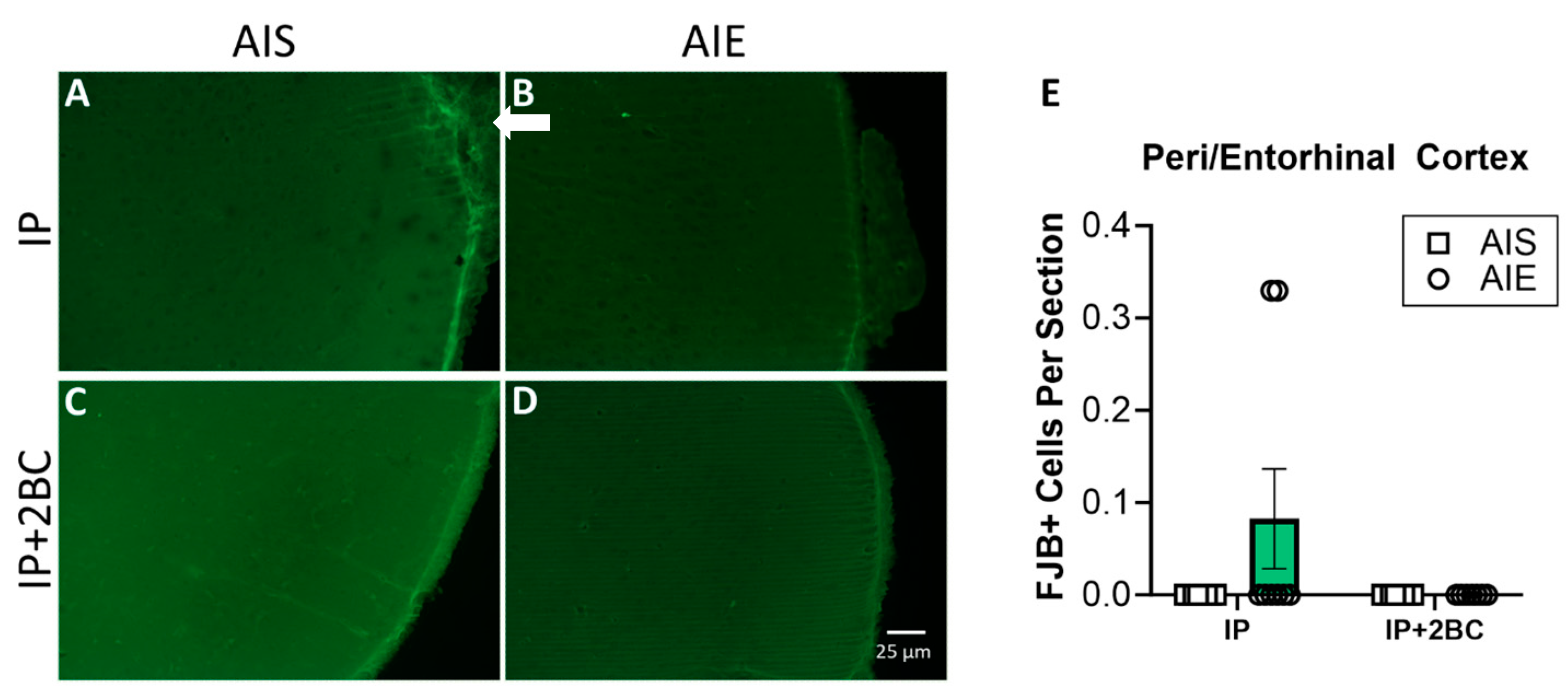
3.3. Neuronal Cell Death Was Not Modified by AIE or Adult Alcohol Consumption
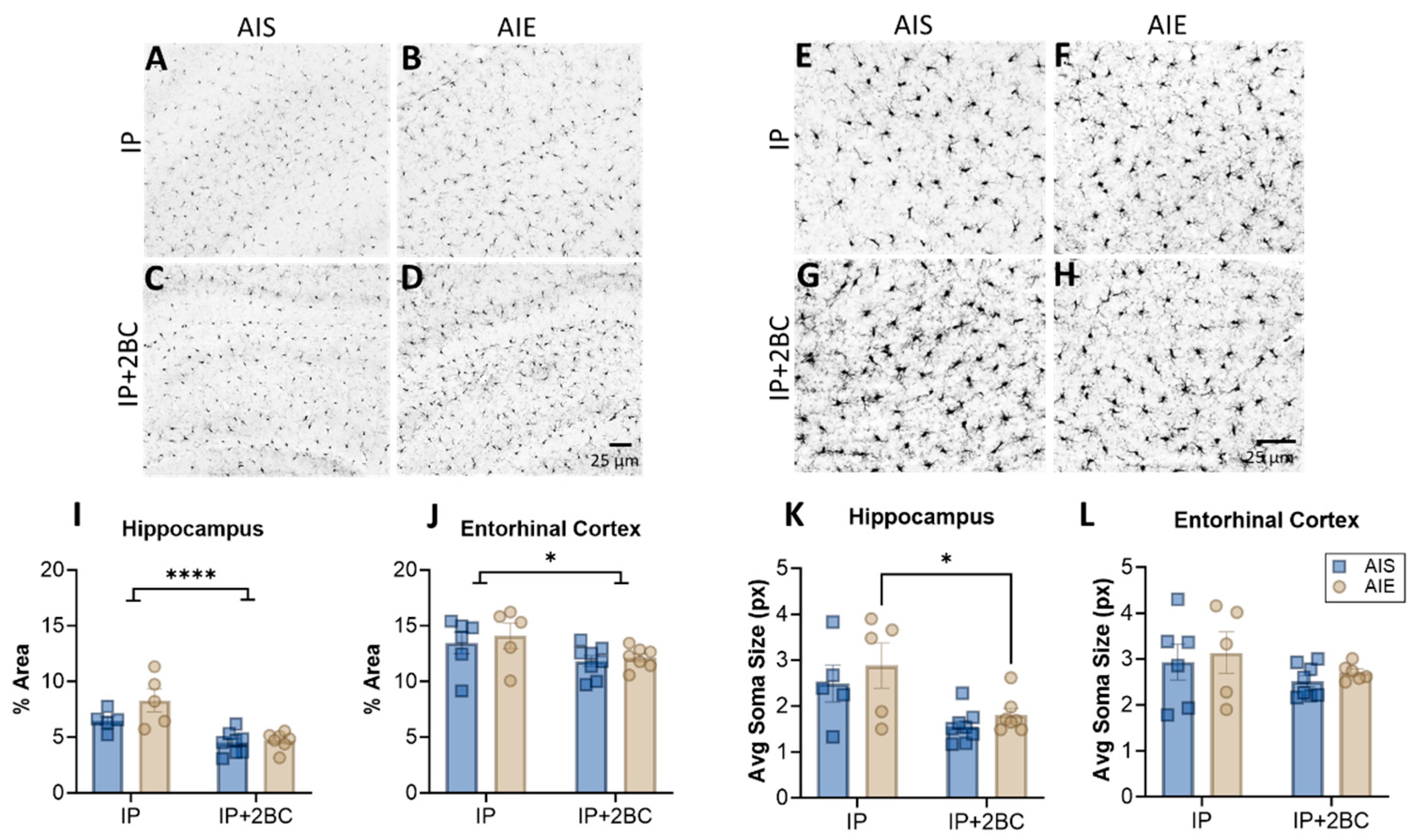
3.4. AIE Does Not Enhance Microglia Reactivity
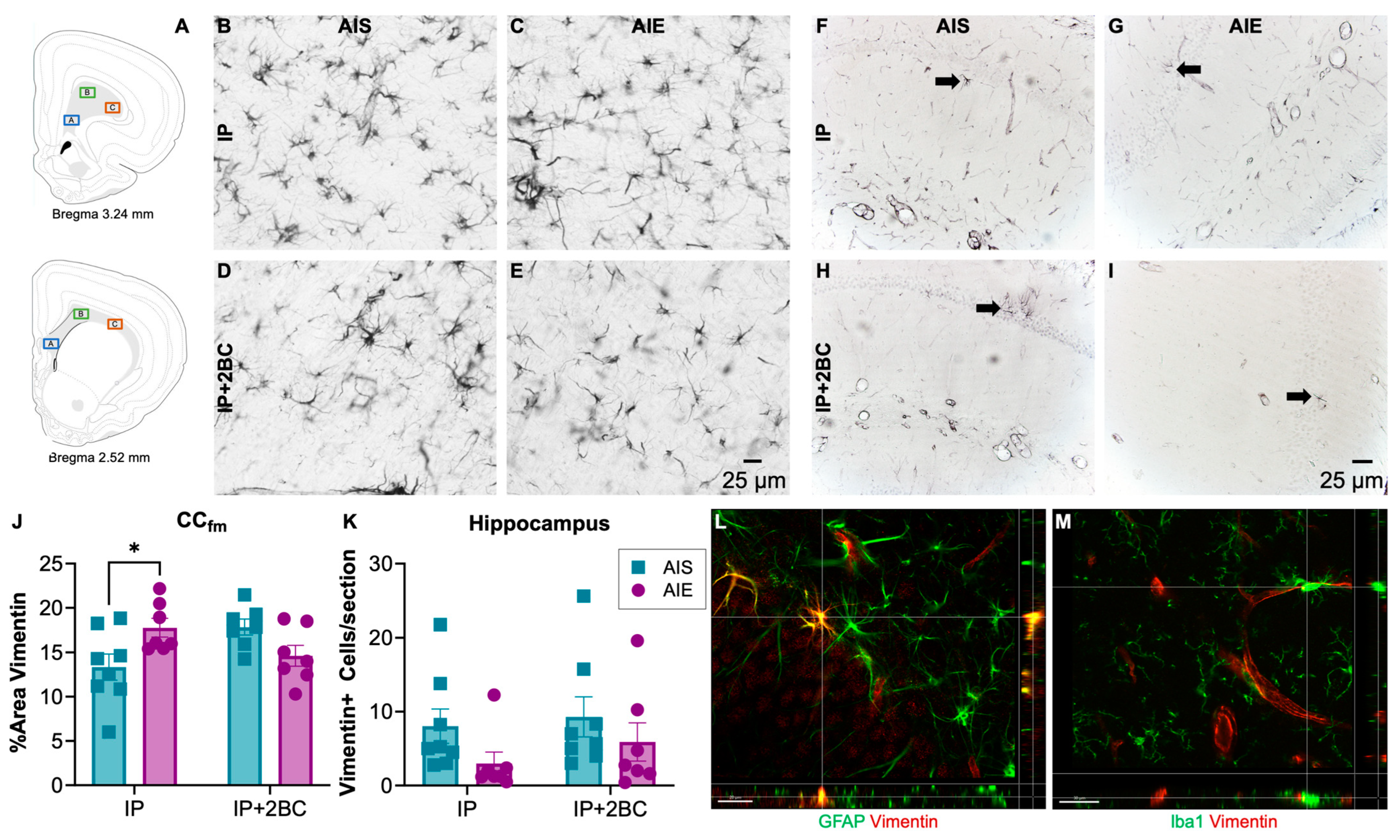
3.5. AIE Increased Astrocyte Reactivity in Adolescence, but Not Following Adult Alcohol Drinking
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Highlights: Child and Adolescent Health and Development: Progress Report 2006–2007; WHO: Geneva, Switzerland, 2008; p. 20.
- SAMHSA. Key Substance Use and Mental Health Indicators in the United States: Results from the 2021 National Survey on Drug Use and Health. 2022. Available online: https://www.samhsa.gov/data/report/2021-nsduh-annual-national-report (accessed on 28 October 2023).
- Miech, R.A.; Johnston, L.D.; Patrick, M.E.; O’Malley, P.M.; Bachman, J.G.; Schulenberg, J.E. Monitoring the Future National Survey Results on Drug Use, 1975–2022: Secondary School Students; Institute for Social Research, University of Michigan: Ann Arbor, MI, USA, 2023. [Google Scholar]
- NIAAA. NIAAA Council approves definition of binge drinking. NIAAA Newsl. 2004, 3, 3. [Google Scholar]
- Chung, T.; Creswell, K.G.; Bachrach, R.; Clark, D.B.; Martin, C.S. Adolescent Binge Drinking. Alcohol. Res. Curr. Rev. 2018, 39, 5–15. [Google Scholar]
- Patrick, M.E.; Terry-McElrath, Y.M. Prevalence of High-Intensity Drinking from Adolescence through Young Adulthood: National Data from 2016–2017. Subst. Abus. 2019, 13, 1178221818822976. [Google Scholar] [CrossRef] [PubMed]
- Patrick, M.E.; Terry-McElrath, Y.M. High-intensity drinking by underage young adults in the United States. Addiction 2017, 112, 82–93. [Google Scholar] [CrossRef]
- DeWit, D.J.; Adlaf, E.M.; Offord, D.R.; Ogborne, A.C. Age at first alcohol use: A risk factor for the development of alcohol disorders. Am. J. Psychiatry 2000, 157, 745–750. [Google Scholar] [CrossRef]
- Grant, B.F.; Dawson, D.A. Age at onset of alcohol use and its association with DSM-IV alcohol abuse and dependence: Results from the National Longitudinal Alcohol Epidemiologic Survey. J. Subst. Abus. 1997, 9, 103–110. [Google Scholar] [CrossRef]
- Rohde, P.; Lewinsohn, P.M.; Seeley, J.R. Psychiatric comorbidity with problematic alcohol use in high school students. J. Am. Acad. Child Adolesc. Psychiatry 1996, 35, 101–109. [Google Scholar] [CrossRef]
- Waller, R.; Murray, L.; Shaw, D.S.; Forbes, E.E.; Hyde, L.W. Accelerated alcohol use across adolescence predicts early adult symptoms of alcohol use disorder via reward-related neural function. Psychol. Med. 2019, 49, 675–684. [Google Scholar] [CrossRef]
- Addolorato, G.; Vassallo, G.A.; Antonelli, G.; Antonelli, M.; Tarli, C.; Mirijello, A.; Agyei-Nkansah, A.; Mentella, M.C.; Ferrarese, D.; Mora, V.; et al. Author Correction: Binge Drinking among adolescents is related to the development of Alcohol Use Disorders: Results from a Cross-Sectional Study. Sci. Rep. 2018, 8, 15476. [Google Scholar] [CrossRef]
- Nixon, K.; McClain, J.A. Adolescence as a critical window for developing an alcohol use disorder: Current findings in neuroscience. Curr. Opin. Psychiatry 2010, 23, 227–232. [Google Scholar] [CrossRef]
- Spear, L.P. Adolescent neurobehavioral characteristics, alcohol sensitivities, and intake: Setting the stage for alcohol use disorders? Child Dev. Perspect. 2011, 5, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Coleman, L.G., Jr.; Crews, F.T.; Vetreno, R.P. The persistent impact of adolescent binge alcohol on adult brain structural, cellular, and behavioral pathology: A role for the neuroimmune system and epigenetics. Int. Rev. Neurobiol. 2021, 160, 1–44. [Google Scholar] [CrossRef] [PubMed]
- Casey, B.J.; Jones, R.M. Neurobiology of the adolescent brain and behavior: Implications for substance use disorders. J. Am. Acad. Child Adolesc. Psychiatry 2010, 49, 1189–1201, quiz 1285. [Google Scholar] [CrossRef] [PubMed]
- Spear, L.P. Adolescents and alcohol: Acute sensitivities, enhanced intake, and later consequences. Neurotox. Teratol. 2014, 41, 51–59. [Google Scholar] [CrossRef]
- Spear, L.P. Effects of adolescent alcohol consumption on the brain and behaviour. Nat. Rev. Neurosci. 2018, 19, 197–214. [Google Scholar] [CrossRef]
- Crews, F.T.; Braun, C.J.; Hoplight, B.; Switzer, R.C., 3rd; Knapp, D.J. Binge ethanol consumption causes differential brain damage in young adolescent rats compared with adult rats. Alcohol. Clin. Exp. Res. 2000, 24, 1712–1723. [Google Scholar] [CrossRef]
- Lacaille, H.; Duterte-Boucher, D.; Liot, D.; Vaudry, H.; Naassila, M.; Vaudry, D. Comparison of the deleterious effects of binge drinking-like alcohol exposure in adolescent and adult mice. J. Neurochem. 2015, 132, 629–641. [Google Scholar] [CrossRef]
- Ransohoff, R.M.; Perry, V.H. Microglial physiology: Unique stimuli, specialized responses. Ann. Rev. Immunol. 2009, 27, 119–145. [Google Scholar] [CrossRef]
- He, J.; Crews, F.T. Increased MCP-1 and microglia in various regions of the human alcoholic brain. Exp. Neurol. 2008, 210, 349–358. [Google Scholar] [CrossRef]
- Melbourne, J.K.; Chandler, C.M.; Van Doorn, C.E.; Bardo, M.T.; Pauly, J.R.; Peng, H.; Nixon, K. Primed for addiction: A critical review of the role of microglia in the neurodevelopmental consequences of adolescent alcohol drinking. Alcohol. Clin. Exp. Res. 2021, 45, 1908–1926. [Google Scholar] [CrossRef]
- Pascual, M.; Montesinos, J.; Guerri, C. Role of the innate immune system in the neuropathological consequences induced by adolescent binge drinking. J. Neurosci. Res. 2018, 96, 765–780. [Google Scholar] [CrossRef] [PubMed]
- Chastain, L.G.; Sarkar, D.K. Role of microglia in regulation of ethanol neurotoxic action. Int. Rev. Neurobiol. 2014, 118, 81–103. [Google Scholar] [CrossRef] [PubMed]
- Crews, F.T.; Vetreno, R.P. Neuroimmune basis of alcoholic brain damage. Int. Rev. Neurobiol. 2014, 118, 315–357. [Google Scholar] [CrossRef] [PubMed]
- Marshall, S.A.; McClain, J.A.; Kelso, M.L.; Hopkins, D.M.; Pauly, J.R.; Nixon, K. Microglial activation is not equivalent to neuroinflammation in alcohol-induced neurodegeneration: The importance of microglia phenotype. Neurobiol. Dis. 2013, 54, 239–251. [Google Scholar] [CrossRef]
- McClain, J.A.; Morris, S.A.; Deeny, M.A.; Marshall, S.A.; Hayes, D.M.; Kiser, Z.M.; Nixon, K. Adolescent binge alcohol exposure induces long-lasting partial activation of microglia. Brain Behav. Immun. 2011, 25 (Suppl. S1), S120–S128. [Google Scholar] [CrossRef]
- Kelso, M.L.; Liput, D.J.; Eaves, D.W.; Nixon, K. Upregulated vimentin suggests new areas of neurodegeneration in a model of an alcohol use disorder. Neuroscience 2011, 197, 381–393. [Google Scholar] [CrossRef]
- Erickson, E.K.; Blednov, Y.A.; Harris, R.A.; Mayfield, R.D. Glial gene networks associated with alcohol dependence. Sci. Rep. 2019, 9, 10949. [Google Scholar] [CrossRef]
- Prinz, M.; Jung, S.; Priller, J. Microglia Biology: One Century of Evolving Concepts. Cell 2019, 179, 292–311. [Google Scholar] [CrossRef]
- Ransohoff, R.M. A polarizing question: Do M1 and M2 microglia exist? Nat. Neurosci. 2016, 19, 987–991. [Google Scholar] [CrossRef]
- Mayfield, J.; Ferguson, L.; Harris, R.A. Neuroimmune signaling: A key component of alcohol abuse. Curr. Opin. Neurobiol. 2013, 23, 513–520. [Google Scholar] [CrossRef]
- Agrawal, R.G.; Hewetson, A.; George, C.M.; Syapin, P.J.; Bergeson, S.E. Minocycline reduces ethanol drinking. Brain Behav. Immun. 2011, 25 (Suppl. S1), S165–S169. [Google Scholar] [CrossRef]
- Warden, A.S.; Wolfe, S.A.; Khom, S.; Varodayan, F.P.; Patel, R.R.; Steinman, M.Q.; Bajo, M.; Montgomery, S.E.; Vlkolinsky, R.; Nadav, T.; et al. Microglia Control Escalation of Drinking in Alcohol-Dependent Mice: Genomic and Synaptic Drivers. Biol. Psychiatry 2020, 88, 910–921. [Google Scholar] [CrossRef]
- Pandey, S.C.; Sakharkar, A.J.; Tang, L.; Zhang, H. Potential role of adolescent alcohol exposure-induced amygdaloid histone modifications in anxiety and alcohol intake during adulthood. Neurobiol. Dis. 2015, 82, 607–619. [Google Scholar] [CrossRef]
- Alaux-Cantin, S.; Warnault, V.; Legastelois, R.; Botia, B.; Pierrefiche, O.; Vilpoux, C.; Naassila, M. Alcohol intoxications during adolescence increase motivation for alcohol in adult rats and induce neuroadaptations in the nucleus accumbens. Neuropharmacology 2013, 67, 521–531. [Google Scholar] [CrossRef]
- Criado, J.R.; Ehlers, C.L. Effects of adolescent ethanol exposure on event-related oscillations (EROs) in the hippocampus of adult rats. Behav. Brain Res. 2010, 210, 164–170. [Google Scholar] [CrossRef]
- Pascual, M.; Boix, J.; Felipo, V.; Guerri, C. Repeated alcohol administration during adolescence causes changes in the mesolimbic dopaminergic and glutamatergic systems and promotes alcohol intake in the adult rat. J. Neurochem. 2009, 108, 920–931. [Google Scholar] [CrossRef]
- Milivojevic, V.; Covault, J. Alcohol exposure during late adolescence increases drinking in adult Wistar rats, an effect that is not reduced by finasteride. Alcohol Alcohol. 2013, 48, 28–38. [Google Scholar] [CrossRef]
- Sakharkar, A.J.; Kyzar, E.J.; Gavin, D.P.; Zhang, H.; Chen, Y.; Krishnan, H.R.; Grayson, D.R.; Pandey, S.C. Altered amygdala DNA methylation mechanisms after adolescent alcohol exposure contribute to adult anxiety and alcohol drinking. Neuropharmacology 2019, 157, 107679. [Google Scholar] [CrossRef]
- Chandler, C.M.; Shaykin, J.D.; Peng, H.; Pauly, J.R.; Nixon, K.; Bardo, M.T. Effects of voluntary adolescent intermittent alcohol exposure and social isolation on adult alcohol intake in male rats. Alcohol 2022, 104, 13–21. [Google Scholar] [CrossRef]
- Chandler, C.M.; Hamid, U.; Maggio, S.E.; Peng, H.; Pauly, J.R.; Beckmann, J.; Nixon, K.; Bardo, M.T. Effects of adolescent alcohol exposure via oral gavage on adult alcohol drinking and co-use of alcohol and nicotine in Sprague Dawley rats. Drug Alcohol. Depend. 2022, 232, 109298. [Google Scholar] [CrossRef]
- Gilpin, N.W.; Karanikas, C.A.; Richardson, H.N. Adolescent binge drinking leads to changes in alcohol drinking, anxiety, and amygdalar corticotropin releasing factor cells in adulthood in male rats. PLoS ONE 2012, 7, e31466. [Google Scholar] [CrossRef] [PubMed]
- Hamidullah, S.; Lutelmowski, C.D.; Creighton, S.D.; Luciani, K.R.; Frie, J.A.; Winters, B.D.; Khokhar, J.Y. Effects of vapourized THC and voluntary alcohol drinking during adolescence on cognition, reward, and anxiety-like behaviours in rats. Prog. Neuro-Psychopharm. Biol. Psych. 2021, 106, 110141. [Google Scholar] [CrossRef] [PubMed]
- Mugantseva, E.; Hyytia, P.; Latvala, A. Voluntary Adolescent-Onset Alcohol Drinking Fails to Influence Alcohol Consumption or Anxiety-Like Behaviour in Adulthood in Female Alcohol-Preferring Rats. Alcohol Alcohol. 2022, 57, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Criado, J.R.; Ehlers, C.L. Effects of adolescent onset voluntary drinking followed by ethanol vapor exposure on subsequent ethanol consumption during protracted withdrawal in adult Wistar rats. Pharm. Biochem. Behav. 2013, 103, 622–630. [Google Scholar] [CrossRef] [PubMed]
- 2010/63/EU; DIRECTIVE 2010/63/EU on the Protection of Animals Used for Scientific Purposes. EU Council Directive: Brussels, Belgium, 2010.
- NRC. Guide for the Care and Use of Laboratory Animals, 8th ed.; The National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Heidbreder, C.A.; Weiss, I.C.; Domeney, A.M.; Pryce, C.; Homberg, J.; Hedou, G.; Feldon, J.; Moran, M.C.; Nelson, P. Behavioral, neurochemical and endocrinological characterization of the early social isolation syndrome. Neuroscience 2000, 100, 749–768. [Google Scholar] [CrossRef]
- Ehlers, C.L.; Walker, B.M.; Pian, J.P.; Roth, J.L.; Slawecki, C.J. Increased alcohol drinking in isolate-housed alcohol-preferring rats. Behav. Neurosci. 2007, 121, 111–119. [Google Scholar] [CrossRef]
- Lopez, M.F.; Doremus-Fitzwater, T.L.; Becker, H.C. Chronic social isolation and chronic variable stress during early development induce later elevated ethanol intake in adult C57BL/6J mice. Alcohol 2011, 45, 355–364. [Google Scholar] [CrossRef]
- Bardo, M.T.; Klebaur, J.E.; Valone, J.M.; Deaton, C. Environmental enrichment decreases intravenous self-administration of amphetamine in female and male rats. Psychopharmacology 2001, 155, 278–284. [Google Scholar] [CrossRef]
- Stairs, D.J.; Klein, E.D.; Bardo, M.T. Effects of environmental enrichment on extinction and reinstatement of amphetamine self-administration and sucrose-maintained responding. Behav. Pharmacol. 2006, 17, 597–604. [Google Scholar] [CrossRef]
- Deehan, G.A., Jr.; Cain, M.E.; Kiefer, S.W. Differential rearing conditions alter operant responding for ethanol in outbred rats. Alcohol. Clin. Exp. Res. 2007, 31, 1692–1698. [Google Scholar] [CrossRef]
- McCool, B.A.; Chappell, A.M. Early social isolation in male Long-Evans rats alters both appetitive and consummatory behaviors expressed during operant ethanol self-administration. Alcohol. Clin. Exp. Res. 2009, 33, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Jang, M.; Kim, W.; Noh, J. Differential effects of pair housing on voluntary nicotine consumption: A comparison between male and female adolescent rats. Psychopharmacology 2017, 234, 2463–2473. [Google Scholar] [CrossRef] [PubMed]
- Gross, A.N.; Richter, S.H.; Engel, A.K.; Wurbel, H. Cage-induced stereotypies, perseveration and the effects of environmental enrichment in laboratory mice. Behav. Brain Res. 2012, 234, 61–68. [Google Scholar] [CrossRef]
- van Praag, H.; Kempermann, G.; Gage, F.H. Neural consequences of environmental enrichment. Nat. Rev. Neurosci. 2000, 1, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Baroncelli, L.; Braschi, C.; Spolidoro, M.; Begenisic, T.; Sale, A.; Maffei, L. Nurturing brain plasticity: Impact of environmental enrichment. Cell Death Diff. 2010, 17, 1092–1103. [Google Scholar] [CrossRef]
- Kempermann, G. Environmental enrichment, new neurons and the neurobiology of individuality. Nat. Rev. Neurosci. 2019, 20, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Pascual, M.; Blanco, A.M.; Cauli, O.; Minarro, J.; Guerri, C. Intermittent ethanol exposure induces inflammatory brain damage and causes long-term behavioural alterations in adolescent rats. Eur. J. Neurosci. 2007, 25, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Vengeliene, V.; Bilbao, A.; Spanagel, R. The alcohol deprivation effect model for studying relapse behavior: A comparison between rats and mice. Alcohol 2014, 48, 313–320. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, Compact 6th ed.; Elsevier: London, UK; Academic Press: London, UK, 2009. [Google Scholar]
- Schmued, L.C.; Hopkins, K.J. Fluoro-Jade B: A high affinity fluorescent marker for the localization of neuronal degeneration. Brain Res. 2000, 874, 123–130. [Google Scholar] [CrossRef]
- Guerin, S.P.; Melbourne, J.K.; Dang, H.Q.; Shaji, C.A.; Nixon, K. Astrocyte Reactivity and Neurodegeneration in the Female Rat Brain Following Alcohol Dependence. Neuroscience 2023, 529, 183–199. [Google Scholar] [CrossRef]
- Leasure, J.L.; Nixon, K. Exercise neuroprotection in a rat model of binge alcohol consumption. Alcohol. Clin. Exp. Res. 2010, 34, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Obernier, J.A.; Bouldin, T.W.; Crews, F.T. Binge ethanol exposure in adult rats causes necrotic cell death. Alcohol. Clin. Exp. Res. 2002, 26, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Ito, D.; Imai, Y.; Ohsawa, K.; Nakajima, K.; Fukuuchi, Y.; Kohsaka, S. Microglia-specific localisation of a novel calcium binding protein, Iba1. Mol. Brain Res. 1998, 57, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Kastner, R.; Szymas, J.; Hossmann, K.A. Immunohistochemical study of glial reaction and serum-protein extravasation in relation to neuronal damage in rat hippocampus after ischemia. Neuroscience 1990, 38, 527–540. [Google Scholar] [CrossRef]
- Zhang, S.C. Defining glial cells during CNS development. Nat. Rev. Neurosci. 2001, 2, 840–843. [Google Scholar] [CrossRef]
- Noori, H.R.; Fornal, C.A. The appropriateness of unbiased optical fractionators to assess cell proliferation in the adult hippocampus. Front. Neurosci. 2011, 5, 140. [Google Scholar] [CrossRef]
- White, A.M.; Swartzwelder, H.S. Hippocampal function during adolescence: A unique target of ethanol effects. Ann. N. Y. Acad. Sci. 2004, 1021, 206–220. [Google Scholar] [CrossRef]
- Wooden, J.I.; Thompson, K.R.; Guerin, S.P.; Nawarawong, N.N.; Nixon, K. Consequences of adolescent alcohol use on adult hippocampal neurogenesis and hippocampal integrity. Int. Rev. Neurobiol. 2021, 160, 281–304. [Google Scholar] [CrossRef]
- Vargas, W.M.; Bengston, L.; Gilpin, N.W.; Whitcomb, B.W.; Richardson, H.N. Alcohol binge drinking during adolescence or dependence during adulthood reduces prefrontal myelin in male rats. J. Neurosci. 2014, 34, 14777–14782. [Google Scholar] [CrossRef]
- Swanson, L.W. Brain maps 4.0-Structure of the rat brain: An open access atlas with global nervous system nomenclature ontology and flatmaps. J. Comp. Neurol. 2018, 526, 935–943. [Google Scholar] [CrossRef]
- Patrick, M.E.; Schulenberg, J.E.; Martz, M.E.; Maggs, J.L.; O’Malley, P.M.; Johnston, L.D. Extreme binge drinking among 12th-grade students in the United States: Prevalence and predictors. JAMA Pediatr. 2013, 167, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Pang, T.Y.; Hannan, A.J.; Lawrence, A.J. Novel approaches to alcohol rehabilitation: Modification of stress-responsive brain regions through environmental enrichment. Neuropharmacology 2019, 145, 25–36. [Google Scholar] [CrossRef]
- Bloom, F.; Lad, P.; Pittman, Q.; Rogers, J. Blood alcohol levels in rats: Non-uniform yields from intraperitoneal doses based on body weight. Br. J. Pharmacol. 1982, 75, 251–254. [Google Scholar] [CrossRef] [PubMed]
- Gass, J.T.; Glen, W.B., Jr.; McGonigal, J.T.; Trantham-Davidson, H.; Lopez, M.F.; Randall, P.K.; Yaxley, R.; Floresco, S.B.; Chandler, L.J. Adolescent alcohol exposure reduces behavioral flexibility, promotes disinhibition, and increases resistance to extinction of ethanol self-administration in adulthood. Neuropsychopharmacology 2014, 39, 2570–2583. [Google Scholar] [CrossRef] [PubMed]
- Salguero, A.; Suarez, A.; Luque, M.; Ruiz-Leyva, L.; Cendan, C.M.; Moron, I.; Pautassi, R.M. Binge-Like, Naloxone-Sensitive, Voluntary Ethanol Intake at Adolescence Is Greater Than at Adulthood, but Does Not Exacerbate Subsequent Two-Bottle Choice Drinking. Front. Behav. Neurosci. 2020, 14, 50. [Google Scholar] [CrossRef]
- Younis, R.M.; Wolstenholme, J.T.; Bagdas, D.; Bettinger, J.C.; Miles, M.F.; Damaj, M.I. Adolescent but not adult ethanol binge drinking modulates ethanol behavioral effects in mice later in life. Pharm. Biochem. Behav. 2019, 184, 172740. [Google Scholar] [CrossRef]
- Blizard, D.A.; Vandenbergh, D.J.; Jefferson, A.L.; Chatlos, C.D.; Vogler, G.P.; McClearn, G.E. Effects of periadolescent ethanol exposure on alcohol preference in two BALB substrains. Alcohol 2004, 34, 177–185. [Google Scholar] [CrossRef]
- Maldonado-Devincci, A.M.; Makdisi, J.G.; Hill, A.M.; Waters, R.C.; Hall, N.I.; Shobande, M.J.; Kumari, A. Adolescent intermittent ethanol exposure induces sex-dependent divergent changes in ethanol drinking and motor activity in adulthood in C57BL/6J mice. J. Neurosci. Res. 2022, 100, 1560–1572. [Google Scholar] [CrossRef]
- Moore, E.M.; Mariani, J.N.; Linsenbardt, D.N.; Melon, L.C.; Boehm, S.L., 2nd. Adolescent C57BL/6J (but not DBA/2J) mice consume greater amounts of limited-access ethanol compared to adults and display continued elevated ethanol intake into adulthood. Alcohol. Clin. Exp. Res. 2010, 34, 734–742. [Google Scholar] [CrossRef]
- Crabbe, J.C.; Wahlsten, D.; Dudek, B.C. Genetics of mouse behavior: Interactions with laboratory environment. Science 1999, 284, 1670–1672. [Google Scholar] [CrossRef]
- Wahlsten, D.; Metten, P.; Phillips, T.J.; Boehm, S.L., 2nd; Burkhart-Kasch, S.; Dorow, J.; Doerksen, S.; Downing, C.; Fogarty, J.; Rodd-Henricks, K.; et al. Different data from different labs: Lessons from studies of gene-environment interaction. J. Neurobiol. 2003, 54, 283–311. [Google Scholar] [CrossRef] [PubMed]
- Sorge, R.E.; Martin, L.J.; Isbester, K.A.; Sotocinal, S.G.; Rosen, S.; Tuttle, A.H.; Wieskopf, J.S.; Acland, E.L.; Dokova, A.; Kadoura, B.; et al. Olfactory exposure to males, including men, causes stress and related analgesia in rodents. Nat. Methods 2014, 11, 629–632. [Google Scholar] [CrossRef] [PubMed]
- Castelhano-Carlos, M.J.; Baumans, V. The impact of light, noise, cage cleaning and in-house transport on welfare and stress of laboratory rats. Lab. Anim. 2009, 43, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Robinson, G.; Most, D.; Ferguson, L.B.; Mayfield, J.; Harris, R.A.; Blednov, Y.A. Neuroimmune pathways in alcohol consumption: Evidence from behavioral and genetic studies in rodents and humans. Int. Rev. Neurobiol. 2014, 118, 13–39. [Google Scholar] [CrossRef] [PubMed]
- Crews, F.T.; Vetreno, R.P. Mechanisms of neuroimmune gene induction in alcoholism. Psychopharmacology 2016, 233, 1543–1557. [Google Scholar] [CrossRef]
- Crews, F.T.; Lawrimore, C.J.; Walter, T.J.; Coleman, L.G., Jr. The role of neuroimmune signaling in alcoholism. Neuropharmacology 2017, 122, 56–73. [Google Scholar] [CrossRef]
- Mayfield, J.; Harris, R.A. The Neuroimmune Basis of Excessive Alcohol Consumption. Neuropsychopharmacology 2017, 42, 376. [Google Scholar] [CrossRef]
- Blednov, Y.A.; Ponomarev, I.; Geil, C.; Bergeson, S.; Koob, G.F.; Harris, R.A. Neuroimmune regulation of alcohol consumption: Behavioral validation of genes obtained from genomic studies. Addict. Biol. 2012, 17, 108–120. [Google Scholar] [CrossRef]
- Crews, F.T.; Vetreno, R.P.; Broadwater, M.A.; Robinson, D.L. Adolescent Alcohol Exposure Persistently Impacts Adult Neurobiology and Behavior. Pharm. Rev. 2016, 68, 1074–1109. [Google Scholar] [CrossRef]
- Crews, F.T.; Qin, L.; Sheedy, D.; Vetreno, R.P.; Zou, J. High mobility group box 1/Toll-like receptor danger signaling increases brain neuroimmune activation in alcohol dependence. Biol. Psychiatry 2013, 73, 602–612. [Google Scholar] [CrossRef]
- Fernandez-Lizarbe, S.; Pascual, M.; Guerri, C. Critical role of TLR4 response in the activation of microglia induced by ethanol. J. Immunol. 2009, 183, 4733–4744. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Wang, D.; Zhang, Y.; Cai, Z.; Ye, T.; Tong, L.; Xu, X.; Lu, J.; Liu, F.; Lu, X.; et al. Apoptosis-triggered decline in hippocampal microglia mediates adolescent intermittent alcohol exposure-induced depression-like behaviors in mice. Neuropharmacology 2020, 170, 108054. [Google Scholar] [CrossRef] [PubMed]
- Marshall, S.A.; McClain, J.A.; Wooden, J.I.; Nixon, K. Microglia Dystrophy Following Binge-Like Alcohol Exposure in Adolescent and Adult Male Rats. Front. Neuroanat. 2020, 14, 52. [Google Scholar] [CrossRef] [PubMed]
- Blednov, Y.A.; Benavidez, J.M.; Geil, C.; Perra, S.; Morikawa, H.; Harris, R.A. Activation of inflammatory signaling by lipopolysaccharide produces a prolonged increase of voluntary alcohol intake in mice. Brain Behav. Immun. 2011, 25 (Suppl. S1), S92–S105. [Google Scholar] [CrossRef]
- McKenna, I.M.; Waalkes, M.P.; Chen, L.C.; Gordon, T. Comparison of inflammatory lung responses in Wistar rats and C57 and DBA mice following acute exposure to cadmium oxide fumes. Toxicol. Appl. Pharmacol. 1997, 146, 196–206. [Google Scholar] [CrossRef]
- Strong, M.N.; Yoneyama, N.; Fretwell, A.M.; Snelling, C.; Tanchuck, M.A.; Finn, D.A. “Binge” drinking experience in adolescent mice shows sex differences and elevated ethanol intake in adulthood. Horm. Behav. 2010, 58, 82–90. [Google Scholar] [CrossRef]
- Boschen, K.E.; Ptacek, T.S.; Berginski, M.E.; Simon, J.M.; Parnell, S.E. Transcriptomic analyses of gastrulation-stage mouse embryos with differential susceptibility to alcohol. Dis. Model. Mech. 2021, 14, dmm049012. [Google Scholar] [CrossRef]
- Ronchi, J.A.; Figueira, T.R.; Ravagnani, F.G.; Oliveira, H.C.; Vercesi, A.E.; Castilho, R.F. A spontaneous mutation in the nicotinamide nucleotide transhydrogenase gene of C57BL/6J mice results in mitochondrial redox abnormalities. Free Radic. Biol. Med. 2013, 63, 446–456. [Google Scholar] [CrossRef]
- Gano, A.; Vore, A.S.; Sammakia, M.N.; Deak, T. Assessment of Extracellular Cytokines in the Hippocampus of the Awake Behaving Rat Using Large-Molecule Microdialysis Combined with Multiplex Arrays After Acute and Chronic Ethanol Exposure. Alcohol. Clin. Exp. Res. 2019, 43, 640–654. [Google Scholar] [CrossRef]
- Peng, H.; Nixon, K. Microglia Phenotypes Following the Induction of Alcohol Dependence in Adolescent Rats. Alcohol. Clin. Exp. Res. 2021, 45, 105–116. [Google Scholar] [CrossRef]
- Doremus-Fitzwater, T.L.; Gano, A.; Paniccia, J.E.; Deak, T. Male adolescent rats display blunted cytokine responses in the CNS after acute ethanol or lipopolysaccharide exposure. Physiol. Behav. 2015, 148, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Marshall, S.A.; Geil, C.R.; Nixon, K. Prior Binge Ethanol Exposure Potentiates the Microglial Response in a Model of Alcohol-Induced Neurodegeneration. Brain Sci. 2016, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Beynon, S.B.; Walker, F.R. Microglial activation in the injured and healthy brain: What are we really talking about? Practical and theoretical issues associated with the measurement of changes in microglial morphology. Neuroscience 2012, 225, 162–171. [Google Scholar] [CrossRef]
- Vidal-Itriago, A.; Radford, R.A.W.; Aramideh, J.A.; Maurel, C.; Scherer, N.M.; Don, E.K.; Lee, A.; Chung, R.S.; Graeber, M.B.; Morsch, M. Microglia morphophysiological diversity and its implications for the CNS. Front. Immunol. 2022, 13, 997786. [Google Scholar] [CrossRef] [PubMed]
- Tesone-Coelho, C.; Varela, P.; Escosteguy-Neto, J.C.; Cavarsan, C.F.; Mello, L.E.; Santos-Junior, J.G. Effects of ethanol on hippocampal neurogenesis depend on the conditioned appetitive response. Addict. Biol. 2013, 18, 774–785. [Google Scholar] [CrossRef] [PubMed]
- Marshall, S.A.; Casachahua, J.D.; Rinker, J.A.; Blose, A.K.; Lysle, D.T.; Thiele, T.E. IL-1 receptor signaling in the basolateral amygdala modulates binge-like ethanol consumption in male C57BL/6J mice. Brain Behav. Immun. 2016, 51, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Crews, F.T.; Mdzinarishvili, A.; Kim, D.; He, J.; Nixon, K. Neurogenesis in adolescent brain is potently inhibited by ethanol. Neuroscience 2006, 137, 437–445. [Google Scholar] [CrossRef]
- Hayes, D.M.; Deeny, M.A.; Shaner, C.A.; Nixon, K. Determining the threshold for alcohol-induced brain damage: New evidence with gliosis markers. Alcohol. Clin. Exp. Res. 2013, 37, 425–434. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wooden, J.I.; Peacoe, L.E.; Anasooya Shaji, C.; Melbourne, J.K.; Chandler, C.M.; Bardo, M.T.; Nixon, K. Adolescent Intermittent Ethanol Drives Modest Neuroinflammation but Does Not Escalate Drinking in Male Rats. Cells 2023, 12, 2572. https://doi.org/10.3390/cells12212572
Wooden JI, Peacoe LE, Anasooya Shaji C, Melbourne JK, Chandler CM, Bardo MT, Nixon K. Adolescent Intermittent Ethanol Drives Modest Neuroinflammation but Does Not Escalate Drinking in Male Rats. Cells. 2023; 12(21):2572. https://doi.org/10.3390/cells12212572
Chicago/Turabian StyleWooden, Jessica I., Lauren E. Peacoe, Chinchusha Anasooya Shaji, Jennifer K. Melbourne, Cassie M. Chandler, Michael T. Bardo, and Kimberly Nixon. 2023. "Adolescent Intermittent Ethanol Drives Modest Neuroinflammation but Does Not Escalate Drinking in Male Rats" Cells 12, no. 21: 2572. https://doi.org/10.3390/cells12212572