

Stability and Requirement for Thiamin in a Cell Culture Feed Used to Produce New Biological Entities

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Stability Studies in Feed
2.3. LC-MS Feature Determination and Structure Elucidation
2.4. Fed-Batch Cultivation in Spin Tubes
2.5. Antibody Purification and Critical Quality Attribute Analysis
2.6. Metabolomics
2.6.1. Extracellular Metabolomics
2.6.2. Intracellular Metabolomics
2.6.3. LC-MS/MS
2.7. Cytotoxicity Assay
2.8. Statistics
3. Results
3.1. Thiamin Stability
3.1.1. Thiamin Stability in Cell Culture Feed
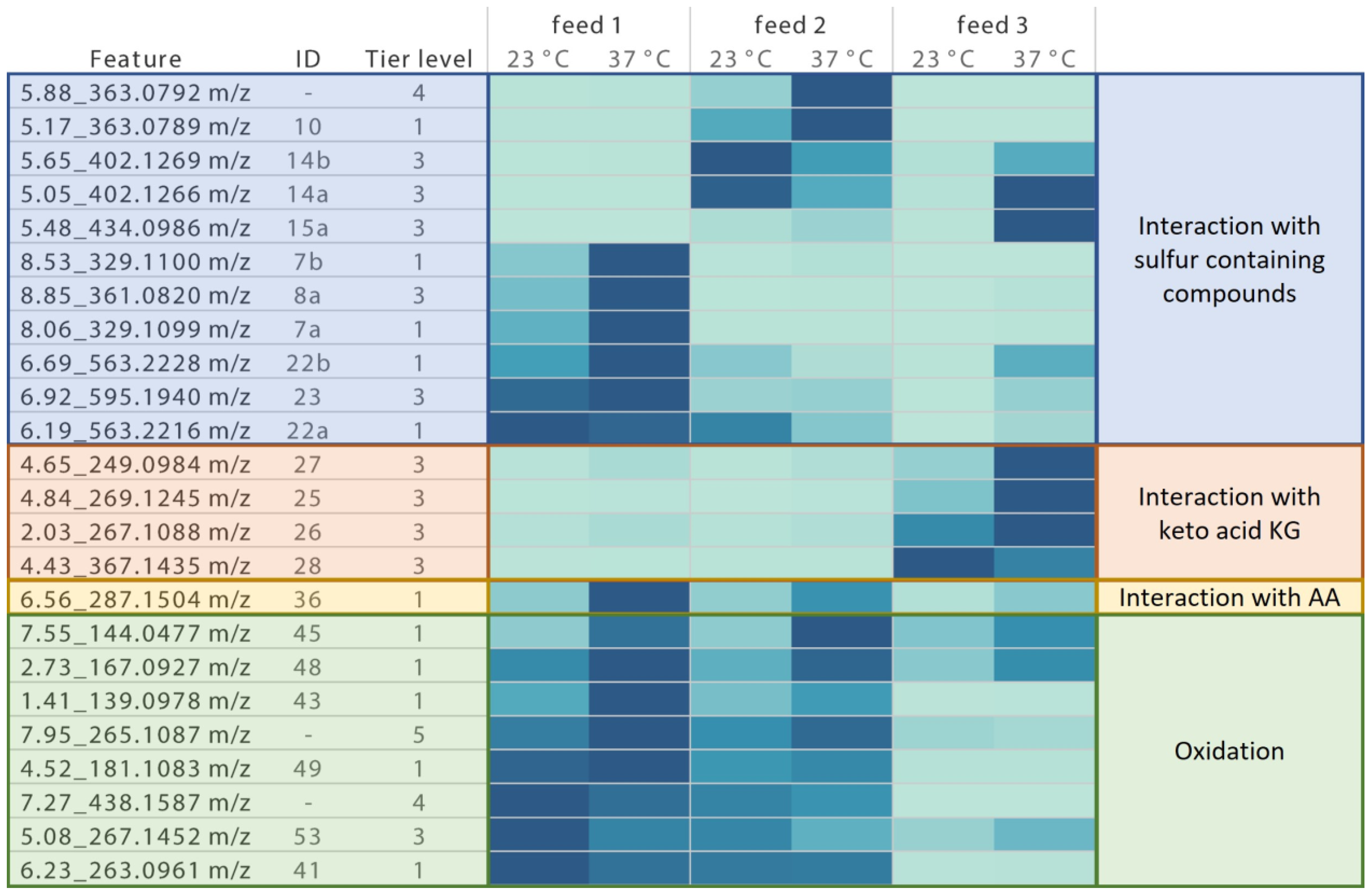
3.1.2. Thiamin Degradation and Interaction Product Identification
3.1.3. Quantification of Formed Degradation Products in Stored Feed
3.1.4. Toxicity of Thiamin-Derived Degradation Products in CHO Cells
3.2. Thiamin Necessity
3.2.1. Cell Performance and cQA Analysis
3.2.2. Thiamin Phosphate Species
3.2.3. Extracellular Metabolomics
3.2.4. Intracellular Metabolomics
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Landauer, K. Designing media for animal cell culture: CHO cells, the industrial standard. In Animal Cell Biotechnology, 2nd ed.; Pörtner, R., Ed.; Humana Press: Totowa, NJ, USA, 2014; Volume 1104, pp. 89–103. [Google Scholar]
- Bellmaine, S.; Schnellbaecher, A.; Zimmer, A. Reactivity and degradation products of tryptophan in solution and proteins. Free Radic. Biol. Med. 2020, 160, 696–718. [Google Scholar] [CrossRef] [PubMed]
- Ambrogelly, A. The Different Colors of mAbs in Solution. Antibodies 2021, 10, 21. [Google Scholar] [CrossRef]
- Zang, L.; Frenkel, R.; Simeone, J.; Lanan, M.; Byers, M.; Lyubarskaya, Y. Metabolomics profiling of cell culture media leading to the identification of riboflavin photosensitized degradation of tryptophan causing slow growth in cell culture. Anal. Chem. 2011, 83, 5422–5430. [Google Scholar] [CrossRef] [PubMed]
- Schnellbaecher, A.; Binder, D.; Bellmaine, S.; Zimmer, A. Vitamins in cell culture media: Stability and stabilization strategies. Biotechnol. Bioeng. 2019, 116, 1537–1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnellbaecher, A.; Lindig, A.; Le Mignon, M.; Hofmann, T.; Pardon, B.; Bellmaine, S.; Zimmer, A. Degradation Products of Tryptophan in Cell Culture Media: Contribution to Color and Toxicity. Int. J. Mol. Sci. 2021, 22, 6221. [Google Scholar] [CrossRef]
- Yoshimoto, S.; Kohara, N.; Sato, N.; Ando, H.; Ichihashi, M. Riboflavin Plays a Pivotal Role in the UVA-Induced Cytotoxicity of Fibroblasts as a Key Molecule in the Production of H2O2 by UVA Radiation in Collaboration with Amino Acids and Vitamins. Int. J. Mol. Sci. 2020, 21, 554. [Google Scholar] [CrossRef] [Green Version]
- McCoy, R.E.; Costa, N.A.; Morris, A.E. Factors that determine stability of highly concentrated chemically defined production media. Biotechnol. Prog. 2015, 31, 493–502. [Google Scholar] [CrossRef]
- Burdick, D. Thiamine (B1). In Kirk-Othmer Encyclopedia of Chemical Technology; Ley, C., Ed.; John Wiley & Sons: New York, NY, USA, 2000. [Google Scholar]
- Lonsdale, D. Thiamin. In Advances in Food and Nutrition Research; Eskin, N.A.M., Ed.; Academic Press: London, UK, 2018; Volume 83, pp. 1–56. [Google Scholar]
- Kochetov, G.A.; Solovjeva, O.N. Structure and functioning mechanism of transketolase. BBA Proteins Proteom. 2014, 1844, 1608–1618. [Google Scholar] [CrossRef]
- Dhir, S.; Tarasenko, M.; Napoli, E.; Giulivi, C. Neurological, Psychiatric, and Biochemical Aspects of Thiamine Deficiency in Children and Adults. Front. Psychiatry 2019, 10, 207. [Google Scholar] [CrossRef] [Green Version]
- Gubler, C.J. Thiamin. In Handbook of Vitamins, 2nd ed.; Machlin, L.J., Ed.; Marcel Dekker: New York, NY, USA, 1991; pp. 234–281. [Google Scholar]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef]
- Blaženović, I.; Kind, T.; Sa, M.R.; Ji, J.; Vaniya, A.; Wancewicz, B.; Roberts, B.S.; Torbašinović, H.; Lee, T.; Mehta, S.S.; et al. Structure Annotation of All Mass Spectra in Untargeted Metabolomics. Anal. Chem. 2019, 91, 2155–2162. [Google Scholar] [CrossRef] [PubMed]
- Weiss, C.H.; Merkel, C.; Zimmer, A. Impact of iron raw materials and their impurities on CHO metabolism and recombinant protein product quality. Biotechnol. Prog. 2021, 37, e3148. [Google Scholar] [CrossRef] [PubMed]
- Ehret, J.; Zimmermann, M.; Eichhorn, T.; Zimmer, A. Impact of cell culture media additives on IgG glycosylation produced in Chinese hamster ovary cells. Biotechnol. Bioeng. 2019, 116, 816–830. [Google Scholar] [PubMed] [Green Version]
- Matsukawa, T.; Iwatsu, T.; Yurugi, S. Studies on Vitamin B1 and Its Related Compounds. XIV Behaviour of vitamin B1 in water. J. Pharm. Soc. Jpn. 1951, 71, 369–371. [Google Scholar] [CrossRef] [Green Version]
- Lhoest, W.; Busse, L.; Baumann, C. Nonenzymatic destruction of thiamine. J. Am. Pharm. Assoc. 1958, 47, 254–257. [Google Scholar] [CrossRef]
- Watanabe, A.; Asm, Y. Physico-chemical studies of vitamin B1 and its related compounds. 5. Decomposition of thiamine in alkaline solution. J. Pharm. Soc. Jpn. 1957, 77, 153–156. [Google Scholar] [CrossRef] [Green Version]
- Matsukawa, T. On the Mechanism of Vitamin B1 Activities. II: The Oxidation of Vitamin B1 (2). J. Pharm. Soc. Jpn. 1950, 70, 28–32. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, B.K.; Arnold, R.G. Chemistry of thiamine degradation: Mechanisms of thiamine degradation in a model system. J. Food Sci. 1972, 37, 886–888. [Google Scholar] [CrossRef]
- Voelker, A.L.; Taylor, L.S.; Mauer, L.J. Effect of pH and concentration on the chemical stability and reaction kinetics of thiamine mononitrate and thiamine chloride hydrochloride in solution. BMC Chem. 2021, 15, 47. [Google Scholar] [CrossRef]
- Kuschelewski, J.; Schnellbaecher, A.; Pering, S.; Wehsling, M.; Zimmer, A. Antioxidant effect of thiazolidine molecules in cell culture media improves stability and performance. Biotechnol. Prog. 2017, 33, 759–770. [Google Scholar] [CrossRef]
- Miller, C.S.; Sprague, J.M.; Krampitz, L.O. The reaction of thiamine with carbonyl compounds. Ann. N. Y. Acad. Sci. 1962, 98, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Gigliobianco, T.; Lakaye, B.; Wins, P.; El Moualij, B.; Zorzi, W.; Bettendorff, L. Adenosine thiamine triphosphate accumulates in Escherichia coli cells in response to specific conditions of metabolic stress. BMC Microbiol. 2010, 10, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, F. Thiamin enzymology. In Comprehensive Natural Products II: Chemistry and Biology, 1st ed.; Mander, L., Liu, H.-W., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands, 2010; Volume 7, pp. 561–598. [Google Scholar]
- Zoltewicz, J.A.; Uray, G. Thiamin: A critical evaluation of recent chemistry of the pyrimidine ring. Bioorg. Chem. 1994, 22, 1–28. [Google Scholar] [CrossRef]
- Palmieri, F.; Monné, M.; Fiermonte, G.; Palmieri, L. Mitochondrial transport and metabolism of the vitamin B-derived cofactors thiamine pyrophosphate, coenzyme A, FAD and NAD+, and related diseases: A review. IUBMB Life 2022, 74, 592–617. [Google Scholar] [CrossRef]
- Tylicki, A.; Łotowski, Z.; Siemieniuk, M.; Ratkiewicz, A. Thiamine and selected thiamine antivitamins—Biological activity and methods of synthesis. Biosci. Rep. 2018, 38, BSR20171148. [Google Scholar] [CrossRef] [Green Version]
- Bettendorff, L. Thiamine. In Present Knowledge in Nutrition, 10th ed.; Erdman, J.W., Jr., MacDonald, I.A., Zeisel, S.H., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2012; pp. 171–188. [Google Scholar]
- Altamirano, C.; Illanes, A.; Canessa, R.; Becerra, S. Specific nutrient supplementation of defined serum-free medium for the improvement of CHO cells growth and t-PA production. Electron. J. Biotechnol. 2006, 9, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Eagle, H. Nutrition needs of mammalian cells in tissue culture. Science 1955, 122, 501–514. [Google Scholar] [CrossRef]
- Ritacco, F.V.; Wu, Y.; Khetan, A. Cell culture media for recombinant protein expression in Chinese hamster ovary (CHO) cells: History, key components, and optimization strategies. Biotechnol. Prog. 2018, 34, 1407–1426. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.L.; Williams, M.W.; Schenley, R.L.; Perdue, S.W.; Hayden, T.L.; Turner, J.E.; Hsie, A.W. The toxicity of sixteen metallic compounds in Chinese hamster ovary cells. Toxicol. Appl. Pharmacol. 1984, 74, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Weiss, C.H.; Caspari, J.S.; Merkel, C.; Zimmer, A. Copper impurity of iron raw material contributes to improved cell culture performance. Biotechnol. Prog. 2022, 38, e3251. [Google Scholar] [CrossRef]
- Kobayashi, K. Thermal decomposition of thiamine at the neutral range. (I) Effect of cupric ion. Vitamins 1972, 45, 239–246. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schnellbächer, A.; Zimmer, A. Stability and Requirement for Thiamin in a Cell Culture Feed Used to Produce New Biological Entities. Cells 2023, 12, 334. https://doi.org/10.3390/cells12020334
Schnellbächer A, Zimmer A. Stability and Requirement for Thiamin in a Cell Culture Feed Used to Produce New Biological Entities. Cells. 2023; 12(2):334. https://doi.org/10.3390/cells12020334
Chicago/Turabian StyleSchnellbächer, Alisa, and Aline Zimmer. 2023. "Stability and Requirement for Thiamin in a Cell Culture Feed Used to Produce New Biological Entities" Cells 12, no. 2: 334. https://doi.org/10.3390/cells12020334