
A Novel NCI-H69AR Drug-Resistant Small-Cell Lung Cancer Mini-Tumor Model for Anti-Cancer Treatment Screening

Abstract
:
1. Introduction
2. Materials and Methods
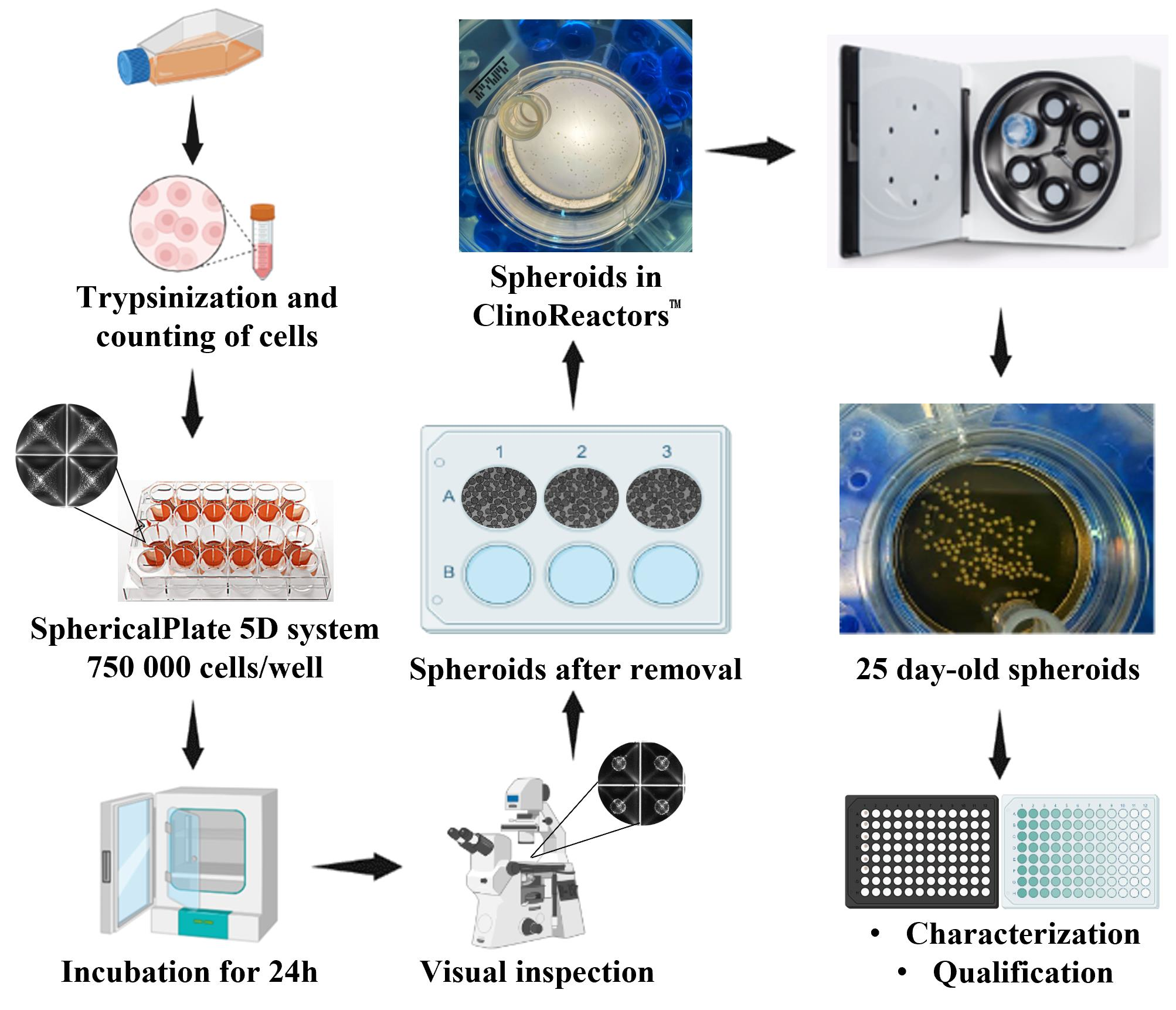
2.1. Two-Dimensional Culturing of NCI-H69AR Cells
2.2. Spheroid Formation and Maintenance
2.3. Characterization of the NCI-H69AR Spheroid Model
2.3.1. Planimetry
2.3.2. The Bradford Soluble Protein Assay
2.3.3. Intracellular Adenosine Triphosphate Cell Viability Assay
2.3.4. Extracellular Adenylate Kinase Cell Death Assay
2.3.5. Approximate Glucose Consumption
2.3.6. qRT-PCR Analysis of the Samples
2.3.7. pH Measurements
2.3.8. Histological Analysis
2.4. Evaluation of the NHI-H69AR Spheroid Model for Anticancer Treatment Screening
2.5. Statistical Data Analysis
3. Results
3.1. Characterization of the NCI-H69AR Spheroid Model
3.1.1. Planimetry
3.1.2. Soluble Protein Content
3.1.3. Intracellular ATP Content
3.1.4. Extracellular Adenylate Kinase Content
3.1.5. Approximate Glucose Consumption
3.1.6. Efflux Transporter Relative Gene Expression
3.1.7. Histology
3.2. Evaluation of the NHI-H69AR Spheroid Model Treatment Reactivity
3.2.1. Irinotecan Treatment Reactivity
3.2.2. Paclitaxel Treatment Reactivity
3.2.3. Cisplatin Treatment Reactivity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Deo, S.V.S.; Sharma, J.; Kumar, S. GLOBOCAN 2020 Report on Global Cancer Burden: Challenges and Opportunities for Surgical Oncologists. Ann. Surg. Oncol. 2022, 29, 6497–6500. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Barta, J.A.; Powell, C.A.; Wisnivesky, J.P. Global Epidemiology of Lung Cancer. Ann. Glob. Health 2019, 85, 8. [Google Scholar] [CrossRef] [Green Version]
- Oser, M.G.; Niederst, M.J.; Sequist, L.V.; Engelman, J.A. Transformation from non-small-cell lung cancer to small-cell lung cancer: Molecular drivers and cells of origin. Lancet Oncol. 2015, 16, e165–e172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, X.; Li, Y.; Wang, H.; Jia, T.; Wang, E.; Luo, Y.; Wei, Y.; Qin, Z.; Ma, X. Small cell lung cancer transformation: From pathogenesis to treatment. Semin. Cancer Biol. 2022, 86, 595–606. [Google Scholar] [CrossRef]
- Mamdani, H.; Induru, R.; Jalal, S.I. Novel therapies in small cell lung cancer. Transl. Lung Cancer Res. 2015, 4, 533–544. [Google Scholar] [CrossRef]
- Khanna, P.; Blais, N.; Gaudreau, P.-O.; Corrales-Rodriguez, L. Immunotherapy Comes of Age in Lung Cancer. Clin. Lung Cancer 2017, 18, 13–22. [Google Scholar] [CrossRef]
- Oronsky, B.; Reid, T.R.; Oronsky, A.; Carter, C.A. What’s new in SCLC? A review. Neoplasia 2017, 19, 842–847. [Google Scholar] [CrossRef]
- Corral, J.M.; del Puerto-Nevado, L.; Cedeño, M.; Río-Vilariño, A.; Mahillo-Fernández, I.; Galeano, C.; Baños, N.; García-Foncillas, J.; Dómine, M.; Cebrián, A. Galectin-1, a novel promising target for outcome prediction and treatment in SCLC. Biomed. Pharmacother. 2022, 156, 113987. [Google Scholar] [CrossRef]
- Arriola, E.; Trigo, J.M.; Sánchez-Gastaldo, A.; Navarro, A.; Perez, C.; Crama, L.; Ponce-Aix, S. Prognostic Value of Clinical Staging According to TNM in Patients With SCLC: A Real-World Surveillance Epidemiology and End-Results Database Analysis. JTO Clin. Res. Rep. 2022, 3, 1–5. [Google Scholar] [CrossRef]
- Shanker, M.; Willcutts, D.; Roth, J.A.; Ramesh, R. Drug resistance in lung cancer. Lung Cancer (Auckl) 2010, 1, 23–36. [Google Scholar] [PubMed]
- Triller, N.; Korošec, P.; Kern, I.; Košnik, M.; Debeljak, A. Multidrug resistance in small cell lung cancer: Expression of P-glycoprotein, multidrug resistance protein 1 and lung resistance protein in chemo-naive patients and in relapsed disease. Lung Cancer 2006, 54, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Tan, Y.; Na, D.E.Y.M.; Bai, Y.; Gao, K. Effect of expression levels of multidrug resistance gene-related protein 1, P-glycoprotein and topoisomerase II on paclitaxel, gemcitabine and vinorelbine sensitivity in pulmonary cancer. Trop. J. Pharm. Res. 2019, 18, 1597–1601. [Google Scholar]
- Chen, Y.-T.; Feng, B.; Chen, L.-B. Update of Research on Drug Resistance in Small Cell Lung Cancer Chemotherapy. Asian Pac. J. Cancer Prev. 2012, 13, 3577–3581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailly, C. Irinotecan: 25 years of cancer treatment. Pharmacol. Res. 2019, 148, 104398. [Google Scholar] [CrossRef]
- Huang, X.; Wang, M.; You, Q.; Kong, J.; Zhang, H.; Yu, C.; Wang, Y.; Wang, H.; Huang, R. Synthesis, mechanisms of action, and toxicity of novel aminophosphonates derivatives conjugated irinotecan in vitro and in vivo as potent antitumor agents. Eur. J. Med. Chem. 2020, 189, 112067. [Google Scholar] [CrossRef]
- Xu, Y.; Villalona-Calero, M.A. Irinotecan: Mechanisms of tumor resistance and novel strategies for modulating its activity. Ann. Oncol. 2002, 13, 1841–1851. [Google Scholar] [CrossRef]
- de Jong, F.A.; de Jonge, M.J.A.; Verweij, J.; Mathijssen, R.H.J. Role of pharmacogenetics in irinotecan therapy. Cancer Lett. 2006, 234, 90–106. [Google Scholar] [CrossRef]
- Chen, Z.; Shi, T.; Zhang, L.; Zhu, P.; Deng, M.; Huang, C.; Hu, T.; Jiang, L.; Li, J. Mammalian drug efflux transporters of the ATP binding cassette (ABC) family in multidrug resistance: A review of the past decade. Cancer Lett. 2016, 370, 153–164. [Google Scholar] [CrossRef]
- Ren, X.; Zhao, B.; Chang, H.; Xiao, M.; Wu, Y.; Liu, Y. Paclitaxel suppresses proliferation and induces apoptosis through regulation of ROS and the AKT/MAPK signaling pathway in canine mammary gland tumor cells. Mol. Med. Rep. 2018, 17, 8289–8299. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.R.; Wang, J.-Q.; Yang, D.-H.; Xu, X.-X. Paclitaxel resistance related to nuclear envelope structural sturdiness. Drug Resist. Updat. 2022, 65, 100881. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.R.; Xu, X.-X. Breaking malignant nuclei as a non-mitotic mechanism of taxol-paclitaxel. J. Cancer Biol. 2021, 2, 86–93. [Google Scholar] [CrossRef]
- Gottesman, M.M. Mechanisms of Cancer Drug Resistance. Annu. Rev. Med. 2002, 53, 615–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maloney, S.M.; Hoover, C.A.; Morejon-Lasso, L.V.; Prosperi, J.R. Mechanisms of Taxane Resistance. Cancers 2020, 12, 3323. [Google Scholar] [CrossRef]
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gualdani, R.; de Clippele, M.; Ratbi, I.; Gailly, P.; Tajeddine, N. Store-Operated Calcium Entry Contributes to Cisplatin-Induced Cell Death in Non-Small Cell Lung Carcinoma. Cancers 2019, 11, 430. [Google Scholar] [CrossRef] [Green Version]
- Tchounwou, P.B.; Dasari, S.; Noubissi, F.K.; Ray, P.; Kumar, S. Advances in Our Understanding of the Molecular Mechanisms of Action of Cisplatin in Cancer Therapy. J. Exp. Pharmacol. 2021, 13, 303–328. [Google Scholar] [CrossRef]
- Aldossary, S.A. Review on Pharmacology of Cisplatin: Clinical Use, Toxicity and Mechanism of Resistance of Cisplatin. Biomed. Pharmacol. J. 2019, 12, 7–15. [Google Scholar] [CrossRef]
- Santo, V.E.; Estrada, M.F.; Rebelo, S.P.; Abreu, S.; Silva, I.; Pinto, C.; Veloso, S.C.; Serra, A.T.; Boghaert, E.; Alves, P.M.; et al. Adaptable stirred-tank culture strategies for large scale production of multicellular spheroid-based tumor cell models. J. Biotechnol. 2016, 221, 118–129. [Google Scholar] [CrossRef]
- Carvalho, M.R.; Lima, D.; Reis, R.L.; Oliveira, J.M.; Correlo, V.M. Anti-Cancer Drug Validation: The Contribution of Tissue Engineered Models. Stem Cell Rev. Rep. 2017, 13, 347–363. [Google Scholar] [CrossRef] [Green Version]
- Barzegari, A.; Saei, A.A. An Update to Space Biomedical Research: Tissue Engineering in Microgravity Bioreactors. BioImpacts 2012, 2, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Madsen, N.H.; Nielsen, B.S.; Larsen, J.; Gad, M. In vitro 2D and 3D cancer models to evaluate compounds that modulate macrophage polarization. Cell. Immunol. 2022, 378, 104574. [Google Scholar] [CrossRef]
- Ferreira, L.; Gaspar, V.; Mano, J. Design of spherically structured 3D in vitro tumor models -Advances and prospects. Acta Biomater. 2018, 75, 11–34. [Google Scholar] [CrossRef] [PubMed]
- Wanigasekara, J.; Cullen, P.J.; Bourke, P.; Tiwari, B.; Curtin, J.F. Advances in 3D culture systems for therapeutic discovery and development in brain cancer. Drug Discov. Today 2023, 28, 103426. [Google Scholar] [CrossRef]
- Antoni, D.; Burckel, H.; Josset, E.; Noel, G. Three-Dimensional Cell Culture: A Breakthrough in Vivo. Int. J. Mol. Sci. 2015, 16, 5517–5527. [Google Scholar] [CrossRef]
- Breslin, S.; O’driscoll, L. Three-dimensional cell culture: The missing link in drug discovery. Drug Discov. Today 2013, 18, 240–249. [Google Scholar] [CrossRef]
- Pozzi, S.; Scomparin, A.; Dangoor, S.I.; Ajamil, D.R.; Ofek, P.; Neufeld, L.; Krivitsky, A.; Vaskovich-Koubi, D.; Kleiner, R.; Dey, P.; et al. Meet me halfway: Are in vitro 3D cancer models on the way to replace in vivo models for nanomedicine development? Adv. Drug Deliv. Rev. 2021, 175, 113760. [Google Scholar] [CrossRef] [PubMed]
- Achilli, T.-M.; Meyer, J.; Morgan, J.R. Advances in the formation, use and understanding of multi-cellular spheroids. Expert Opin. Biol. Ther. 2012, 12, 1347–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Loggenberg, S.; Willers, C.; van der Kooy, F.; Gouws, C.; Hamman, J.H.; Steyn, J.D. Evaluating in vitro cytotoxic effects of Artemisia afra and Artemisia annua infusions against selected lung cancer cell lines. S. Afr. J. Bot. 2022, 150, 404–411. [Google Scholar] [CrossRef]
- Stransky, S.; Cutler, R.; Aguilan, J.; Nieves, E.; Sidoli, S. Investigation of reversible histone acetylation and dynamics in gene expression regulation using 3D liver spheroid model. Epigenetics Chromatin 2022, 15, 35. [Google Scholar] [CrossRef] [PubMed]
- Merwe, L.; Svitina, H.; Willers, C.; Wrzesinski, K.; Gouws, C. A novel NCI-H69V small cell lung cancer functional mini-tumor model for future treatment screening applications. Biotechnol. Prog. 2022, 38, e3253. [Google Scholar] [CrossRef]
- Frandsen, H.S.; Vej-Nielsen, J.M.; Smith, L.E.; Sun, L.; Mikkelsen, K.L.; Thulesen, A.P.; Hagensen, C.E.; Yang, F.; Rogowska-Wrzesinska, A. Mapping Proteome and Lipidome Changes in Early-Onset Non-Alcoholic Fatty Liver Disease Using Hepatic 3D Spheroids. Cells 2022, 11, 3216. [Google Scholar] [CrossRef]
- Calitz, C.; Hamman, J.H.; Viljoen, A.M.; Fey, S.J.; Wrzesinski, K.; Gouws, C. Toxicity and anti-prolific properties of Xysmalobium undulatum water extract during short-term exposure to two-dimensional and three-dimensional spheroid cell cultures. Toxicol. Mech. Methods 2018, 28, 641–652. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ganti, A.K.P.; Loo, B.W.; Bassetti, M.; Blakely, C.; Chiang, A.; A D’Amico, T.; D’Avella, C.; Dowlati, A.; Downey, R.J.; Edelman, M.; et al. Small cell lung cancer, version 2.2022, NCCN clinical practice guidelines in oncology. J. Natl. Compr. Cancer Netw. 2021, 19, 1441–1464. [Google Scholar] [CrossRef] [PubMed]
- Duda, K.; Majerczak, J.; Nieckarz, Z.; Heymsfield, S.B.; Zoladz, J.A. Chapter 1—Human Body Composition and Muscle Mass. In Muscle and Exercise Physiology; Zoladz, J.A., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 3–26. [Google Scholar]
- Wojdyla, K.; Wrzesinski, K.; Williamson, J.; Fey, S.J.; Rogowska-Wrzesinska, A. Acetaminophen-induced S-nitrosylation and S-sulfenylation signalling in 3D cultured hepatocarcinoma cell spheroids. Toxicol. Res. 2016, 5, 905–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCoach, D.B. Hierarchical Linear Modeling. In The Reviewer’s Guide to Quantitative Methods in the Social Sciences; Hancock, G.R., Mueller, R.O., Eds.; Routledge: New York, NY, USA, 2010; pp. 123–140. [Google Scholar]
- Wrzesinski, K.; Frandsen, H.S.; Calitz, C.; Gouws, C.; Korzeniowska, B.; Fey, S.J. Clinostat 3D Cell Culture: Protocols for the Preparation and Functional Analysis of Highly Reproducible, Large, Uniform Spheroids and Organoids. Next Gener. Cult. Platf. Reliab. Vitr. Models Methods Protoc. 2021, 2273, 17–62. [Google Scholar] [CrossRef]
- Cole, S.P.; Sparks, K.E.; Fraser, K.; Loe, D.W.; Grant, C.E.; Wilson, G.M.; Deeley, R.G. Pharmacological characterization of multidrug resistant MRP-transfected human tumor cells. Cancer Res. 1994, 54, 5902–5910. [Google Scholar]
- Ji, L.; Li, H.; Gao, P.; Shang, G.; Zhang, D.D.; Zhang, N.; Jiang, T. Nrf2 Pathway Regulates Multidrug-Resistance-Associated Protein 1 in Small Cell Lung Cancer. PLoS ONE 2013, 8, e63404. [Google Scholar] [CrossRef] [Green Version]
- Henness, S.; Davey, M.W.; Harvie, R.M.; Davey, R.A. Fractionated irradiation of H69 small-cell lung cancer cells causes stable radiation and drug resistance with increased MRP1, MRP2, and topoisomerase IIα expression. Int. J. Radiat. Oncol. 2002, 54, 895–902. [Google Scholar] [CrossRef]
- Helleday, T.; Petermann, E.; Lundin, C.; Hodgson, B.; Sharma, R.A. DNA repair pathways as targets for cancer therapy. Nat. Rev. Cancer 2008, 8, 193–204. [Google Scholar] [CrossRef]
- Salehan, M.R.; Morse, H.R. DNA damage repair and tolerance: A role in chemotherapeutic drug resistance. Br. J. Biomed. Sci. 2013, 70, 31–40. [Google Scholar] [CrossRef]
- Muriithi, W.; Macharia, L.W.; Heming, E.A.C.P.; Echevarria, J.L.; Nyachieo, A.; Filho, P.N.; Neto, V.M. ABC transporters and the hallmarks of cancer: Roles in cancer aggressiveness beyond multidrug resistance. Cancer Biol. Med. 2020, 17, 253–269. [Google Scholar] [CrossRef]
- Fiorillo, M.; Ózsvári, B.; Sotgia, F.; Lisanti, M.P. High ATP Production Fuels Cancer Drug Resistance and Metastasis: Implications for Mitochondrial ATP Depletion Therapy. Front. Oncol. 2021, 11, 3875. [Google Scholar] [CrossRef]
- Synold, T.W.; Dussault, I.; Forman, B.M. The orphan nuclear receptor SXR coordinately regulates drug metabolism and efflux. Nat. Med. 2001, 7, 584–590. [Google Scholar] [CrossRef]
- Smit, T.; Calitz, C.; Willers, C.; Svitina, H.; Hamman, J.; Fey, S.J.; Gouws, C.; Wrzesinski, K. Characterization of an Alginate Encapsulated LS180 Spheroid Model for Anti-colorectal Cancer Compound Screening. ACS Med. Chem. Lett. 2020, 11, 1014–1021. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Kang, Y.; Chen, L.; Wang, H.; Liu, J.; Zeng, S.; Yu, L. The Drug-Resistance Mechanisms of Five Platinum-Based Antitumor Agents. Front. Pharmacol. 2020, 11, 343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fey, S.J.; Wrzesinski, K. Determination of Drug Toxicity Using 3D Spheroids Constructed from an Immortal Human Hepatocyte Cell Line. Toxicol. Sci. 2012, 127, 403–411. [Google Scholar] [CrossRef]
- Kijanska, M.; Kelm, J. In vitro 3D spheroids and microtissues: ATP-based cell viability and toxicity assays. In Assay Guidance Manual; Markossian, S., Sittampalam, G.C., Brimacombe, N.P., Grossman, K., Arkin, M., Eds.; Eli Lilly & Company and the Nasional Centre for Advancing Translational Sciences: Bethesda, MD, USA, 2017; pp. 355–385. [Google Scholar]
- Kamiloglu, S.; Sari, G.; Ozdal, T.; Capanoglu, E. Guidelines for cell viability assays. Food Front. 2020, 1, 332–349. [Google Scholar] [CrossRef]
- Klepinin, A.; Zhang, S.; Klepinina, L.; Rebane-Klemm, E.; Terzic, A.; Kaambre, T.; Dzeja, P. Adenylate kinase and metabolic signaling in cancer cells. Front. Oncol. 2020, 10, 660–669. [Google Scholar] [CrossRef]
- Jacobs, A.C.; DiDone, L.; Jobson, J.; Sofia, M.K.; Krysan, D.; Dunman, P.M. Adenylate Kinase Release as a High-Throughput-Screening-Compatible Reporter of Bacterial Lysis for Identification of Antibacterial Agents. Antimicrob. Agents Chemother. 2013, 57, 26–36. [Google Scholar] [CrossRef] [Green Version]
- Raso, M.G.; Bota-Rabassedas, N.; Wistuba, I. Pathology and Classification of SCLC. Cancers 2021, 13, 820. [Google Scholar] [CrossRef]
- Travis, W.D. Update on small cell carcinoma and its differentiation from squamous cell carcinoma and other non-small cell carcinomas. Mod. Pathol. 2012, 25, S18–S30. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Qiao, T. Unraveling tumor microenvironment of small-cell lung cancer: Implications for immunotherapy. Semin. Cancer Biol. 2022, 86, 117–125. [Google Scholar] [CrossRef]
- Munoz, M.; Henderson, M.; Haber, M.; Norris, M. Role of the MRP1/ABCC1 multidrug transporter protein in cancer. IUBMB Life 2007, 59, 752–757. [Google Scholar] [CrossRef]
- Pote, M.S.; Gacche, R.N. ATP-binding cassette efflux transporters and MDR in cancer. Drug Discov. Today 2023, 28, 103537. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Chen, W.; Yuan, Z.; Liu, X.; Jiang, H. LncRNA-MALAT1 contributes to the cisplatin-resistance of lung cancer by upregulating MRP1 and MDR1 via STAT3 activation. Biomed. Pharmacother. 2018, 101, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.H.W.; Kuo, M.T. Role of Glutathione in the Regulation of Cisplatin Resistance in Cancer Chemotherapy. Met. Based Drugs 2010, 2010, 430939. [Google Scholar] [CrossRef]
- Ullah, E.; Ali, U.; Ah, N.; Naseem, N. Mucin Histochemistry in Tumours of Colon, Ovaries and Lung. J. Cytol. Histol. 2012, 3, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Lakshmanan, I.; Ponnusamy, M.P.; Macha, M.A.; Haridas, D.; Majhi, P.D.; Kaur, S.; Jain, M.; Batra, S.K.; Ganti, A.K. Mucins in Lung Cancer: Diagnostic, Prognostic, and Therapeutic Implications. J. Thorac. Oncol. 2015, 10, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Jonckheere, N.; Skrypek, N.; Van Seuningen, I. Mucins and tumor resistance to chemotherapeutic drugs. Biochim. et Biophys. Acta (BBA)—Rev. Cancer 2014, 1846, 142–151. [Google Scholar] [CrossRef]
- Ning, Y.; Zheng, H.; Zhan, Y.; Liu, S.; Yang, Y.; Zang, H.; Luo, J.; Wen, Q.; Fan, S. Comprehensive analysis of the mechanism and treatment significance of Mucins in lung cancer. J. Exp. Clin. Cancer Res. 2020, 39, 1–10. [Google Scholar] [CrossRef]
- Ham, S.Y.; Kwon, T.; Bak, Y.; Yu, J.-H.; Hong, J.; Lee, S.K.; Yu, D.-Y.; Yoon, D.-Y. Mucin 1-mediated chemo-resistance in lung cancer cells. Oncogenesis 2016, 5, e185. [Google Scholar] [CrossRef] [PubMed]
- Wrzesinski, K.; Alnøe, S.; Jochumsen, H.H.; Mikkelsen, K.; Bryld, T.D.; Vistisen, J.S.; Alnøe, P.W.; Fey, S.J. A Purpose-Built System for Culturing Cells as In Vivo Mimetic 3D Structures, in Biomechanics and Functional Tissue Engineering; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Park, Y.; Huh, K.M.; Kang, S.-W. Applications of Biomaterials in 3D Cell Culture and Contributions of 3D Cell Culture to Drug Development and Basic Biomedical Research. Int. J. Mol. Sci. 2021, 22, 2491. [Google Scholar] [CrossRef]
- Alzeeb, G.; Metges, J.-P.; Corcos, L.; Le Jossic-Corcos, C. Three-Dimensional Culture Systems in Gastric Cancer Research. Cancers 2020, 12, 2800. [Google Scholar] [CrossRef] [PubMed]
- Fontana, F.; Raimondi, M.; Marzagalli, M.; Sommariva, M.; Gagliano, N.; Limonta, P. Three-Dimensional Cell Cultures as an In Vitro Tool for Prostate Cancer Modeling and Drug Discovery. Int. J. Mol. Sci. 2020, 21, 6806. [Google Scholar] [CrossRef]
- Lv, D.; Hu, Z.; Lu, L.; Lu, H.; Xu, X. Three-dimensional cell culture: A powerful tool in tumor research and drug discovery. Oncol. Lett. 2017, 14, 6999–7010. [Google Scholar] [CrossRef] [Green Version]
- Reidy, E.; Leonard, N.A.; Treacy, O.; Ryan, A.E. A 3D View of Colorectal Cancer Models in Predicting Therapeutic Responses and Resistance. Cancers 2021, 13, 227. [Google Scholar] [CrossRef] [PubMed]
- Hoyle, H.; Stenger, C.; Przyborski, S. Design considerations of benchtop fluid flow bioreactors for bio-engineered tissue equivalents in vitro. Biomater. Biosyst. 2022, 8, 100063. [Google Scholar] [CrossRef]
- Kitaeva, K.V.; Rutland, C.S.; Rizvanov, A.A.; Solovyeva, V.V. Cell Culture Based in vitro Test Systems for Anticancer Drug Screening. Front. Bioeng. Biotechnol. 2020, 8, 322. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Oliva, R.; Domínguez-García, S.; Carrascal, L.; Abalos-Martínez, J.; Pardillo-Díaz, R.; Verástegui, C.; Castro, C.; Nunez-Abades, P.; Geribaldi-Doldán, N. Evolution of Experimental Models in the Study of Glioblastoma: Toward Finding Efficient Treatments. Front. Oncol. 2021, 10, 614295. [Google Scholar] [CrossRef] [PubMed]
- Rossouw, M.J. Evaluation of the Efficacy of Selected Anticancer Compounds in Multidrug Resistant Cell Culture Models. Master’s Thesis, North-West University (South-Africa), Potchefstroom Campus, Potchefstroom, South-Africa, 2019. Available online: https://repository.nwu.ac.za/handle/10394/32773 (accessed on 20 May 2023).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Untreated NCI-H69AR 96h Treatment Gene Expression Values | |||||
| Relative MRP1 Gene Expression | Relative> MRP2 Gene Expression | Relative P-gp Gene Expression | |||
| 1.371 | ±0.442 | 1.25 | ±0.504 | 0.151 | ±0.069 |
| Normalized Average Surface Area | Normalized Soluble Protein Content per Spheroid | Normalized ATP/Prot. Values | Normalized AK Release per µg Protein | Normalized Glucose Consumption per µg Protein | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 96 h Treatment | 96 h Recovery | 96 h Treatment | 96 h Recovery | 96 h Treatment | 96 h Recovery | 96 h Treatment | 96 h Recovery | 96 h Treatment | 96 h Recovery | |
| Untreated control | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Irinotecan | 0.79 | 1.05 | 0.78 | 0.99 | 1.19 | 1.05 | 0.69 | 1.57 | 1.37 | 1.11 |
| Paclitaxel | 0.69 | 0.98 | 0.58 | 0.85 | 1.54 | 1.34 | 2.92 | 7.99 | 1.94 | 1.26 |
| Cisplatin | 0.97 | 1.00 | 0.86 | 0.95 | 1.11 | 1.03 | 0.40 | 0.73 | 0.63 | 1.10 |
| 3D Culture Method | Advantages | Disadvantages | References | |
|---|---|---|---|---|
| Anchorage-independent methods | Low-adhesion plates | Relatively easy method Low cost Reproducible Potential to study cell–cell interactions Useful in drug screening Uniform spheroid size control | Low throughput Spheroids must be transferred for analysis Low throughput production | [78,79] |
| Hanging drop methods | Relatively easy method Uniform spheroid size Low cost Homogenous spheroids | Small size of spheroids No long-term culturing Cannot be used for drug testing Medium exchange is difficult Lack of extracellular matrix Labor intensive Impossible to conduct migration and invasion studies | [79,80,81] | |
| Magnetic levitation | High efficiency Easy method No specialized medium is required Medium can easily be exchanged | Colors the 3D constructs brown, limiting certain applications Cells adhere to the bottom of the wells Cellular behavior can be affected | [34,81] | |
| Spinner bioreactors | Relatively easy method Mass production Long-term culturing Medium composition is homogenous Can be used for high-throughput screening Not a labor-intensive method Stimulates metabolite transport | Specialized equipment is required High shear forces are present Un-uniform spheroids Medium exchange is difficult High volume of medium is required | [34,81,82] | |
| ClinoStar® system | Relatively easy method Mass production Long-term culturing Medium composition is homogenous Stimulates metabolite transport Low shear forces are present Uniform spheroids Medium exchange is easy | Specialized equipment is required Fixed volume of 10 mL | [77,83] | |
| Anchorage-dependent methods | Scaffolds | Physical strength can easily be adjusted Numerous materials can be used Mimics the in vivo environment Possible to mimic ECM presentation exactly as in the in vivo environment Possible to combine specific growth factors | Homogeneous dispersion of cells is problematic Expensive Labor intensive Possibility that scaffolds can variate | [78,79,80,81] |
| Hydrogels | Resembles tissue-like flexibility Easy to supply water-soluble factors to the cells | Mechanical resistance is low | [78,79] | |
| Other 3D methods | Microfluidic devices | Uniform spheroid formation Possible to control spheroid size Rapid spheroid formation Continuous perfusion Oxygen and nutrient distribution are uniform Constant temperature control | Expensive Labor intensive Requires specialized equipment and expertise | [80,81,84] |
| 3D bioprinters | Hypoxic conditions inside a spheroid Produces complex 3D architecture Intercellular interactions are possible Possible to reproduce paracrine Reproducing 3D tissue architecture is possible Possible to mimic the chemical environment inside a tumor High throughput screening is possible Can mimic the complex interactions between the tumor microenvironment and the ECM | Difficult to standardize the method Reproducibility of the interactions between the cells and the ECM is not possible Expensive consumables and equipment Difficult to position the cells Optimization of the printing resolution still needs to be improved | [84,85] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Niekerk, A.; Wrzesinski, K.; Steyn, D.; Gouws, C. A Novel NCI-H69AR Drug-Resistant Small-Cell Lung Cancer Mini-Tumor Model for Anti-Cancer Treatment Screening. Cells 2023, 12, 1980. https://doi.org/10.3390/cells12151980
van Niekerk A, Wrzesinski K, Steyn D, Gouws C. A Novel NCI-H69AR Drug-Resistant Small-Cell Lung Cancer Mini-Tumor Model for Anti-Cancer Treatment Screening. Cells. 2023; 12(15):1980. https://doi.org/10.3390/cells12151980
Chicago/Turabian Stylevan Niekerk, Alandi, Krzysztof Wrzesinski, Dewald Steyn, and Chrisna Gouws. 2023. "A Novel NCI-H69AR Drug-Resistant Small-Cell Lung Cancer Mini-Tumor Model for Anti-Cancer Treatment Screening" Cells 12, no. 15: 1980. https://doi.org/10.3390/cells12151980