
Inducible Rbpms-CreERT2 Mouse Line for Studying Gene Function in Retinal Ganglion Cell Physiology and Disease
 , ,
, , 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Mice and Tamoxifen Treatment
2.2. Histology and Immunohistochemistry
2.3. Optomotor Response
2.4. Fluorescein Angiography
2.5. Electroretinography
2.6. Optical Coherence Tomography
2.7. RNA-Sequencing (RNA-Seq)
2.8. Quantification and Statistics Analysis
3. Results
3.1. Generation and Identification of Rbpms-CreERT2 Mice
3.2. Assessment of Retinal Feature and Transcriptome in Rbpms-CreERT2 Knock-in Mice
3.3. Assessment of Retinal Function in Rbpms-CreERT2 Knock-in Mice
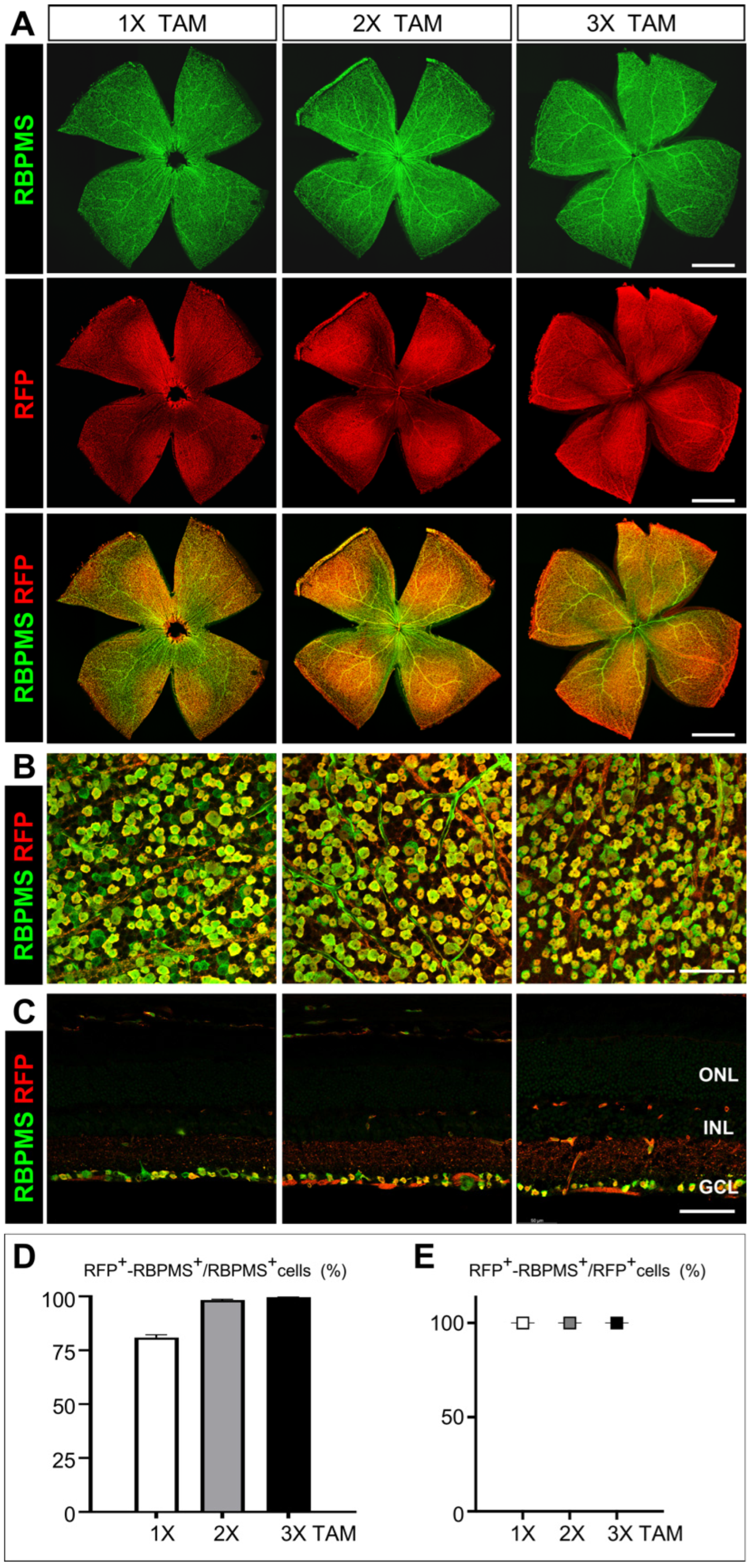
3.4. Assessment of the Cre Recombinase Activity and Efficiency in Activating Reporter Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jeon, C.J.; Strettoi, E.; Masland, R.H. The major cell populations of the mouse retina. J. Neurosci. 1998, 18, 8936–8946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masland, R.H. The Neuronal Organization of the Retina. Neuron 2012, 76, 266–280. [Google Scholar] [CrossRef] [Green Version]
- Dräger, U.C.; Olsen, J.F. Ganglion cell distribution in the retina of the mouse. Investig. Ophthalmol. Vis. Sci. 1981, 20, 285–293. [Google Scholar]
- Pang, J.-J.; Wu, S.M. Morphology and Immunoreactivity of Retrogradely Double-Labeled Ganglion Cells in the Mouse Retina. Investig. Ophthalmol. Vis. Sci. 2011, 52, 4886–4896. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.-H.; Fish, D.R.; Rockhill, R.L.; Masland, R.H. Diversity of ganglion cells in the mouse retina: Unsupervised morphological classification and its limits. J. Comp. Neurol. 2005, 489, 293–310. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Kim, I.J.; Sanes, J.R.; Meister, M. The most numerous ganglion cell type of the mouse retina is a selective feature detector. Proc. Natl. Acad. Sci. USA 2012, 109, E2391–E2398. [Google Scholar] [CrossRef]
- Sanes, J.R.; Masland, R.H. The Types of Retinal Ganglion Cells: Current Status and Implications for Neuronal Classification. Annu. Rev. Neurosci. 2015, 38, 221–246. [Google Scholar] [CrossRef]
- Rheaume, B.A.; Jereen, A.; Bolisetty, M.; Sajid, M.S.; Yang, Y.; Renna, K.; Sun, L.; Robson, P.; Trakhtenberg, E.F. Single cell transcriptome profiling of retinal ganglion cells identifies cellular subtypes. Nat. Commun. 2018, 9, 2759. [Google Scholar] [CrossRef] [Green Version]
- Tran, N.M.; Shekhar, K.; Whitney, I.E.; Jacobi, A.; Benhar, I.; Hong, G.; Yan, W.; Adiconis, X.; Arnold, M.E.; Lee, J.M.; et al. Single-Cell Profiles of Retinal Ganglion Cells Differing in Resilience to Injury Reveal Neuroprotective Genes. Neuron 2019, 104, 1039–1055.e12. [Google Scholar] [CrossRef]
- Lu, Y.; Shiau, F.; Yi, W.; Lu, S.; Wu, Q.; Pearson, J.D.; Kallman, A.; Zhong, S.; Hoang, T.; Zuo, Z.; et al. Single-Cell Analysis of Human Retina Identifies Evolutionarily Conserved and Species-Specific Mechanisms Controlling Development. Dev. Cell 2020, 53, 473–491.e9. [Google Scholar] [CrossRef]
- Goetz, J.; Jessen, Z.F.; Jacobi, A.; Mani, A.; Cooler, S.; Greer, D.; Kadri, S.; Segal, J.; Shekhar, K.; Sanes, J.R.; et al. Unified classification of mouse retinal ganglion cells using function, morphology, and gene expression. Cell Rep. 2022, 40, 111040. [Google Scholar] [CrossRef] [PubMed]
- Whitney, I.E.; Butrus, S.; Dyer, M.A.; Rieke, F.; Sanes, J.R.; Shekhar, K. Vision-Dependent and -Independent Molecular Maturation of Mouse Retinal Ganglion Cells. Neuroscience 2023, 508, 153–173. [Google Scholar] [CrossRef]
- Newman, N.J.; Biousse, V. Hereditary optic neuropathies. Eye 2004, 18, 1144–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu-Wai-Man, P.; Griffiths, P.G.; Hudson, G.; Chinnery, P.F. Inherited mitochondrial optic neuropathies. J. Med. Genet. 2009, 46, 145–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu-Wai-Man, P.; Griffiths, P.G.; Chinnery, P.F. Mitochondrial optic neuropathies–Disease mechanisms and therapeutic strategies. Prog. Retin. Eye Res. 2011, 30, 81–114. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Struebing, F.L.; Geisert, E.E. Commonalities of optic nerve injury and glaucoma-induced neurodegeneration: Insights from transcriptome-wide studies. Exp. Eye Res. 2021, 207, 108571. [Google Scholar] [CrossRef]
- Alexander, C.; Votruba, M.; Pesch, U.E.; Thiselton, D.L.; Mayer, S.; Moore, A.; Rodriguez, M.; Kellner, U.; Leo-Kottler, B.; Auburger, G.; et al. OPA1, encoding a dynamin-related GTPase, is mutated in autosomal dominant optic atrophy linked to chromosome 3q28. Nat. Genet. 2000, 26, 211–215. [Google Scholar] [CrossRef]
- Pesch, U.E.; Leo-Kottler, B.; Mayer, S.; Jurklies, B.; Kellner, U.; Apfelstedt-Sylla, E.; Zrenner, E.; Alexander, C.; Wissinger, B. OPA1 mutations in patients with autosomal dominant optic atrophy and evidence for semi-dominant inheritance. Hum. Mol. Genet. 2001, 10, 1359–1368. [Google Scholar] [CrossRef] [Green Version]
- Wiggs, J.L. Genetic Etiologies of Glaucoma. Arch. Ophthalmol. 2007, 125, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Steinmetz, J.D.; Bourne, R.R.; Briant, P.S.; Flaxman, S.R.; Taylor, H.R.; Jonas, J.B.; Abdoli, A.A.; Abrha, W.A.; Abualhasan, A.; Abu-Gharbieh, E.G.; et al. Causes of blindness and vision impairment in 2020 and trends over 30 years, and prevalence of avoidable blindness in relation to VISION 2020: The Right to Sight: An analysis for the Global Burden of Disease Study. Lancet Glob. Health 2021, 9, e144–e160. [Google Scholar] [CrossRef]
- Wu, F.; Kaczynski, T.J.; Sethuramanujam, S.; Li, R.; Jain, V.; Slaughter, M.; Mu, X. Two transcription factors, Pou4f2 and Isl1, are sufficient to specify the retinal ganglion cell fate. Proc. Natl. Acad. Sci. USA 2015, 112, E1559–E1568. [Google Scholar] [CrossRef]
- Erkman, L.; McEvilly, R.J.; Luo, L.; Ryan, A.K.; Hooshmand, F.; O’Connell, S.M.; Keithley, E.M.; Rapaport, D.H.; Ryan, A.F.; Rosenfeld, M.G. Role of transcription factors a Brn-3.1 and Brn-3.2 in auditory and visual system development. Nature 1996, 381, 603–606. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Xiang, M.; Zhou, L.; Wagner, D.S.; Klein, W.H.; Nathans, J. POU domain factor Brn3b is required for the development of a large set of retinal ganglion cells. Proc. Natl. Acad. Sci. USA 1996, 93, 3920–3925. [Google Scholar] [CrossRef]
- Muzyka, V.V.; Badea, T.C. Genetic interplay between transcription factor Pou4f1/Brn3a and neurotrophin receptor Ret in retinal ganglion cell type specification. Neural Dev. 2021, 16, 5. [Google Scholar] [CrossRef]
- Sajgo, S.; Ghinia, M.G.; Brooks, M.; Kretschmer, F.; Chuang, K.; Hiriyanna, S.; Wu, Z.; Popescu, O.; Badea, T.C. Molecular codes for cell type specification in Brn3 retinal ganglion cells. Proc. Natl. Acad. Sci. USA 2017, 114, E3974–E3983. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Ding, Q.; Xie, X.; Libby, R.T.; Lefebvre, V.; Gan, L. Transcription Factors SOX4 and SOX11 Function Redundantly to Regulate the Development of Mouse Retinal Ganglion Cells. J. Biol. Chem. 2013, 288, 18429–18438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuwajima, T.; Soares, C.A.; Sitko, A.A.; Lefebvre, V.; Mason, C. SoxC Transcription Factors Promote Contralateral Retinal Ganglion Cell Differentiation and Axon Guidance in the Mouse Visual System. Neuron 2017, 93, 1110–1125.e5. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Bard, J.E.; Kann, J.; Yergeau, D.; Sapkota, D.; Ge, Y.; Hu, Z.; Wang, J.; Liu, T.; Mu, X. Single cell transcriptomics reveals lineage trajectory of retinal ganglion cells in wild-type and Atoh7-null retinas. Nat. Commun. 2021, 12, 1465. [Google Scholar] [CrossRef]
- Brown, N.L.; Patel, S.; Brzezinski, J.; Glaser, T. Math5 is required for retinal ganglion cell and optic nerve formation. Development 2001, 128, 2497–2508. [Google Scholar] [CrossRef]
- Miesfeld, J.B.; Ghiasvand, N.M.; Marsh-Armstrong, B.; Marsh-Armstrong, N.; Miller, E.B.; Zhang, P.; Manna, S.K.; Zawadzki, R.J.; Brown, N.L.; Glaser, T. The Atoh7 remote enhancer provides transcriptional robustness during retinal ganglion cell development. Proc. Natl. Acad. Sci. USA 2020, 117, 21690–21700. [Google Scholar] [CrossRef]
- Hegazy, K.A.; Dunn, M.W.; Sharma, S.C. Functional human heme oxygenase has a neuroprotective effect on adult rat ganglion cells after pressure-induced ischemia. NeuroReport 2000, 11, 1185–1189. [Google Scholar] [CrossRef]
- Bond, W.S.; Hines-Beard, J.; GoldenMerry, Y.L.; Davis, M.; Farooque, A.; Sappington, R.M.; Calkins, D.J.; Rex, T.S. Virus-mediated EpoR76E Therapy Slows Optic Nerve Axonopathy in Experimental Glaucoma. Mol. Ther. 2016, 24, 230–239. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Shiga, Y.; Nakagawa, Y.; Fujita, K.; Nishiguchi, K.M.; Tawarayama, H.; Murayama, N.; Maekawa, S.; Yabana, T.; Omodaka, K.; et al. Ecel1 Knockdown With an AAV2-Mediated CRISPR/Cas9 System Promotes Optic Nerve Damage-Induced RGC Death in the Mouse Retina. Investig. Ophthalmol. Vis. Sci. 2018, 59, 3943–3951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Josh Huang, Z.; Zeng, H. Genetic Approaches to Neural Circuits in the Mouse. Annu. Rev. Neurosci. 2013, 36, 183–215. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.A.; Hirokawa, K.E.; Sorensen, S.A.; Gu, H.; Mills, M.; Ng, L.L.; Bohn, P.; Mortrud, M.; Ouellette, B.; Kidney, J.; et al. Anatomical characterization of Cre driver mice for neural circuit mapping and manipulation. Front. Neural Circuits 2014, 8, 76. [Google Scholar] [CrossRef] [Green Version]
- Martersteck, E.M.; Hirokawa, K.E.; Evarts, M.; Bernard, A.; Duan, X.; Li, Y.; Ng, L.; Oh, S.W.; Ouellette, B.; Royall, J.J.; et al. Diverse Central Projection Patterns of Retinal Ganglion Cells. Cell Rep. 2017, 18, 2058–2072. [Google Scholar] [CrossRef]
- Parmhans, N.; Fuller, A.D.; Nguyen, E.; Chuang, K.; Swygart, D.; Wienbar, S.R.; Lin, T.; Kozmik, Z.; Dong, L.; Schwartz, G.W.; et al. Identification of retinal ganglion cell types and brain nuclei expressing the transcription factor Brn3c/Pou4f3 using a Cre recombinase knock-in allele. J. Comp. Neurol. 2021, 529, 1926–1953. [Google Scholar] [CrossRef] [PubMed]
- Simmons, A.B.; Bloomsburg, S.J.; Billingslea, S.A.; Merrill, M.M.; Li, S.; Thomas, M.W.; Fuerst, P.G. Pou4f2 knock-in Cre mouse: A multifaceted genetic tool for vision researchers. Mol. Vis. 2016, 22, 705–717. [Google Scholar] [PubMed]
- Barnstable, C.J.; Drager, U.C. Thy-1 antigen: A ganglion cell specific marker in rodent retina. Neuroscience 1984, 11, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Stella, S.L., Jr.; Li, S.; Sabatini, A.; Vila, A.; Brecha, N.C. Comparison of the ontogeny of the vesicular glutamate transporter 3 (VGLUT3) with VGLUT1 and VGLUT2 in the rat retina. Brain Res. 2008, 1215, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.; Qiao, M.; Bei, F.; Kim, I.J.; He, Z.; Sanes, J.R. Subtype-specific regeneration of retinal ganglion cells following axotomy: Effects of osteopontin and mTOR signaling. Neuron 2015, 85, 1244–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, E.; Hwang, G.S.; Pan, Z.H. Characterization of transgenic mouse lines expressing Cre recombinase in the retina. Neuroscience 2010, 165, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Marquardt, T.; Ashery-Padan, R.; Andrejewski, N.; Scardigli, R.; Guillemot, F.; Gruss, P. Pax6 Is Required for the Multipotent State of Retinal Progenitor Cells. Cell 2001, 105, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Drayson, L.E.; Triplett, J.W. A Chrnb3-Cre BAC transgenic mouse line for manipulation of gene expression in retinal ganglion cells. Genesis 2019, 57, e23305. [Google Scholar] [CrossRef] [PubMed]
- Akerberg, A.A.; Burns, C.E.; Burns, C.G. Exploring the Activities of RBPMS Proteins in Myocardial Biology. Pediatr. Cardiol. 2019, 40, 1410–1418. [Google Scholar] [CrossRef] [PubMed]
- Gerber, W.V.; Vokes, S.A.; Zearfoss, N.R.; Krieg, P.A. A role for the RNA-binding protein, hermes, in the regulation of heart development. Dev. Biol. 2002, 247, 116–126. [Google Scholar] [CrossRef] [Green Version]
- Gan, P.; Wang, Z.; Morales, M.; Zhang, Y.; Bassel-Duby, R.; Liu, N.; Olson, E. RBPMS is an RNA-binding protein that mediates cardiomyocyte binucleation and cardiovascular development. Dev. Cell 2022, 57, 959–973.e7. [Google Scholar] [CrossRef]
- Barnhart, M.D.; Yang, Y.; Nakagaki-Silva, E.E.; Hammond, T.H.; Pizzinga, M.; Gooding, C.; Stott, K.; Smith, C.W.J. Phosphorylation of the smooth muscle master splicing regulator RBPMS regulates its splicing activity. Nucleic Acids Res. 2022, 50, 11895–11915. [Google Scholar] [CrossRef]
- Kwong, J.M.K.; Caprioli, J.; Piri, N. RNA Binding Protein with Multiple Splicing: A New Marker for Retinal Ganglion Cells. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1052–1058. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.R.; de Sevilla Müller, L.P.; Brecha, N.C. The RNA binding protein RBPMS is a selective marker of ganglion cells in the mammalian retina. J. Comp. Neurol. 2014, 522, 1411–1443. [Google Scholar] [CrossRef] [Green Version]
- Nadal-Nicolás, F.M.; Galindo-Romero, C.; Lucas-Ruiz, F.; Marsh-Amstrong, N.; Li, W.; Vidal-Sanz, M.; Agudo-Barriuso, M. Pan-retinal ganglion cell markers in mice, rats, and rhesus macaques. Zool. Res. 2023, 44, 226–248. [Google Scholar] [CrossRef] [PubMed]
- Kwong, J.M.; Quan, A.; Kyung, H.; Piri, N.; Caprioli, J. Quantitative analysis of retinal ganglion cell survival with Rbpms immunolabeling in animal models of optic neuropathies. Investig. Ophthalmol. Vis. Sci. 2011, 52, 9694–9702. [Google Scholar] [CrossRef] [Green Version]
- Fang, F.; Zhuang, P.; Feng, X.; Liu, P.; Liu, D.; Huang, H.; Li, L.; Chen, W.; Liu, L.; Sun, Y.; et al. NMNAT2 is downregulated in glaucomatous RGCs, and RGC-specific gene therapy rescues neurodegeneration and visual function. Mol. Ther. 2022, 30, 1421–1431. [Google Scholar] [CrossRef] [PubMed]
- Jacobi, A.; Tran, N.M.; Yan, W.; Benhar, I.; Tian, F.; Schaffer, R.; He, Z.; Sanes, J.R. Overlapping transcriptional programs promote survival and axonal regeneration of injured retinal ganglion cells. Neuron 2022, 110, 2625–2645.e7. [Google Scholar] [CrossRef] [PubMed]
- Theune, W.C.; Frost, M.P.; Trakhtenberg, E.F. Transcriptomic profiling of retinal cells reveals a subpopulation of microglia/macrophages expressing Rbpms marker of retinal ganglion cells (RGCs) that confound identification of RGCs. Brain Res. 2023, 1811, 148377. [Google Scholar] [CrossRef]
- Tian, F.; Cheng, Y.; Zhou, S.; Wang, Q.; Monavarfeshani, A.; Gao, K.; Jiang, W.; Kawaguchi, R.; Wang, Q.; Tang, M.; et al. Core transcription programs controlling injury-induced neurodegeneration of retinal ganglion cells. Neuron 2022, 110, 2607–2624.e8. [Google Scholar] [CrossRef]
- Madisen, L.; Zwingman, T.A.; Sunkin, S.M.; Oh, S.W.; Zariwala, H.A.; Gu, H.; Ng, L.L.; Palmiter, R.D.; Hawrylycz, M.J.; Jones, A.R.; et al. A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat. Neurosci. 2010, 13, 133–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Hu, F.; Feng, L.; Luo, X.J.; Liang, G.; Zeng, X.Y.; Yi, J.L.; Gan, L. Bhlhb5 is required for the subtype development of retinal amacrine and bipolar cells in mice. Dev. Dyn. 2014, 243, 279–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, H.; Wang, J.; Barwick, S.R.; Yoon, Y.; Smith, S.B. Effect of long-term chronic hyperhomocysteinemia on retinal structure and function in the cystathionine-β-synthase mutant mouse. Exp. Eye Res. 2022, 214, 108894. [Google Scholar] [CrossRef]
- Chng, J.; Wang, T.; Nian, R.; Lau, A.; Hoi, K.M.; Ho, S.C.; Gagnon, P.; Bi, X.; Yang, Y. Cleavage efficient 2A peptides for high level monoclonal antibody expression in CHO cells. MAbs 2015, 7, 403–412. [Google Scholar] [CrossRef] [Green Version]
- Indra, A.K.; Warot, X.; Brocard, J.; Bornert, J.M.; Xiao, J.H.; Chambon, P.; Metzger, D. Temporally-controlled site-specific mutagenesis in the basal layer of the epidermis: Comparison of the recombinase activity of the tamoxifen-inducible Cre-ER(T) and Cre-ER(T2) recombinases. Nucleic Acids Res. 1999, 27, 4324–4327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, M.; Zhou, L.; Macke, J.P.; Yoshioka, T.; Hendry, S.H.; Eddy, R.L.; Shows, T.B.; Nathans, J. The Brn-3 family of POU-domain factors: Primary structure, binding specificity, and expression in subsets of retinal ganglion cells and somatosensory neurons. J. Neurosci. 1995, 15, 4762–4785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunt, A.H.; Lund, R.D. Vinblastine-induced blockage of orthograde and retrograde axonal transport of protein in retinal ganglion cells. Exp. Neurol. 1974, 45, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Perry, V.H. Evidence for an amacrine cell system in the ganglion cell layer of the rat retina. Neuroscience 1981, 6, 931–944. [Google Scholar] [CrossRef]
- Schlamp, C.L.; Montgomery, A.D.; Mac Nair, C.E.; Schuart, C.; Willmer, D.J.; Nickells, R.W. Evaluation of the percentage of ganglion cells in the ganglion cell layer of the rodent retina. Mol. Vis. 2013, 19, 1387–1396. [Google Scholar]
- Nadal-Nicolás, F.M.; Sobrado-Calvo, P.; Jiménez-López, M.; Vidal-Sanz, M.; Agudo-Barriuso, M. Long-Term Effect of Optic Nerve Axotomy on the Retinal Ganglion Cell Layer. Investig. Ophthalmol. Vis. Sci. 2015, 56, 6095–6112. [Google Scholar] [CrossRef]
- Roark, M.W.; Stringham, J.M. Visual Performance in the “Real World”: Contrast Sensitivity, Visual Acuity, and Effects of Macular Carotenoids. Mol. Nutr. Food Res. 2019, 63, 1801053. [Google Scholar] [CrossRef] [Green Version]
- Diez-Roux, G.; Banfi, S.; Sultan, M.; Geffers, L.; Anand, S.; Rozado, D.; Magen, A.; Canidio, E.; Pagani, M.; Peluso, I.; et al. A high-resolution anatomical atlas of the transcriptome in the mouse embryo. PLoS Biol. 2011, 9, e1000582. [Google Scholar] [CrossRef]
- Badea, T.C.; Cahill, H.; Ecker, J.; Hattar, S.; Nathans, J. Distinct roles of transcription factors brn3a and brn3b in controlling the development, morphology, and function of retinal ganglion cells. Neuron 2009, 61, 852–864. [Google Scholar] [CrossRef] [Green Version]
- Hörnberg, H.; Wollerton-van Horck, F.; Maurus, D.; Zwart, M.; Svoboda, H.; Harris, W.A.; Holt, C.E. RNA-binding protein Hermes/RBPMS inversely affects synapse density and axon arbor formation in retinal ganglion cells in vivo. J. Neurosci. 2013, 33, 10384–10395. [Google Scholar] [CrossRef] [Green Version]
- Zearfoss, N.R.; Chan, A.P.; Kloc, M.; Allen, L.H.; Etkin, L.D. Identification of new Xlsirt family members in the Xenopus laevis oocyte. Mech. Dev. 2003, 120, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagaki-Silva, E.E.; Gooding, C.; Llorian, M.; Jacob, A.G.; Richards, F.; Buckroyd, A.; Sinha, S.; Smith, C.W.J. Identification of RBPMS as a mammalian smooth muscle master splicing regulator via proximity of its gene with super-enhancers. eLife 2019, 8, e46327. [Google Scholar] [CrossRef] [PubMed]
- Rowan, S.; Cepko, C.L. Genetic analysis of the homeodomain transcription factor Chx10 in the retina using a novel multifunctional BAC transgenic mouse reporter. Dev. Biol. 2004, 271, 388–402. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Ding, K.; Pan, L.; Deng, M.; Gan, L. Math5 determines the competence state of retinal ganglion cell progenitors. Dev. Biol. 2003, 264, 240–254. [Google Scholar] [CrossRef] [Green Version]
- Brzezinski, J.A.t.; Prasov, L.; Glaser, T. Math5 defines the ganglion cell competence state in a subpopulation of retinal progenitor cells exiting the cell cycle. Dev. Biol. 2012, 365, 395–413. [Google Scholar] [CrossRef] [Green Version]
- Furuta, Y.; Lagutin, O.; Hogan, B.L.; Oliver, G.C. Retina- and ventral forebrain-specific Cre recombinase activity in transgenic mice. Genesis 2000, 26, 130–132. [Google Scholar] [CrossRef]
- Chen, C.-K.J.; Chen, Y.-J.; Shay, A.; Weber, N.; Jiang, Z. Retinal ganglion cells with recombinase activity in mice with engineered Cre in the vGluT3 gene. Investig. Ophthalmol. Vis. Sci. 2020, 61, 4522. [Google Scholar]
- Warthi, G.D.; Faulkner, J.L.; Doja, J.; Ghanam, A.R.; Gao, P.; Yang, A.C.; Slivano, O.J.; Barris, C.T.; Kress, T.C.; Zawieja, S.D.; et al. An Inducible Cre Mouse with Preferential Activity in Vascular Smooth Muscle Evades a Previously Lethal Intestinal Phenotype. bioRxiv 2022. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, L.; Xie, X.; Wang, J.; Xiao, H.; Li, S.; Xu, M.; Quainoo, E.; Koppaka, R.; Zhuo, J.; Smith, S.B.; et al. Inducible Rbpms-CreERT2 Mouse Line for Studying Gene Function in Retinal Ganglion Cell Physiology and Disease. Cells 2023, 12, 1951. https://doi.org/10.3390/cells12151951
Guo L, Xie X, Wang J, Xiao H, Li S, Xu M, Quainoo E, Koppaka R, Zhuo J, Smith SB, et al. Inducible Rbpms-CreERT2 Mouse Line for Studying Gene Function in Retinal Ganglion Cell Physiology and Disease. Cells. 2023; 12(15):1951. https://doi.org/10.3390/cells12151951
Chicago/Turabian StyleGuo, Luming, Xiaoling Xie, Jing Wang, Haiyan Xiao, Shuchun Li, Mei Xu, Ebenezer Quainoo, Rithwik Koppaka, Jiaping Zhuo, Sylvia B. Smith, and et al. 2023. "Inducible Rbpms-CreERT2 Mouse Line for Studying Gene Function in Retinal Ganglion Cell Physiology and Disease" Cells 12, no. 15: 1951. https://doi.org/10.3390/cells12151951