
Interaction of Interleukin-17A with a Th2 Response in a Mouse Model of Allergic Airway Inflammation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
2.2. Whole Body Plethysmography
2.3. In Vitro Cytokine Production of Mouse Lymphocytes
2.4. Preparation of Murine Lungs
2.5. ELISA
2.6. Bronchoalveolar Lavage (BAL)
2.7. Histology
2.8. RNA Isolation and cDNA Synthesis
2.9. Real Time PCR
| Protein | Forward Primer | Reverse Primer |
| mIl17RA | AGTTCCAGTTTCTGTCCATGC | TGGATTTGTGGTTTGGGTC |
| mIl17RC | GCAGAGCCTGAAGAAGCTG | CCCAAGACTAGCCTCGAAAC |
| mRplp0 | CGTCCTCGTTGGAGTGACAT | TAGTTGGACTTCCAGGTCGC |
| mMUC5AC | CCATGCAGAGTCCTCAGAACAA | TTACTGGAAAGGCCCAAGCA |
| mTGFβ1 | CTCCCGTGGCTTCTAGTGC | GCCTTAGTTTGGACAGGATCTG |
| hIL-17RA | GCTTCACCCTGTGGAACGAAT | TATGTGGTGCATGTGCTCAAA |
| hIL-17RC | GATGGTGACAACGTGCATCTG | CAAGGTAATGATCTGCGGTCC |
| hIL-8 | CTTGGCAGCCTTCCTGATTT | TTCTTTAGCACTCCTTGGCAAAA |
| hRPLP0 | CTGGAAGTCCAACTACTTCCT | CATCATGGTGTTCTTGCCCAT |
| hTGFβ1 | GGCGTGCTAATGGTGGAAAA | TGTGTGTACTCTGCTTGAACTTGTCA |
2.10. Immunofluorescence Staining and Detection
2.11. Cell Culture
2.12. Statistical Analysis
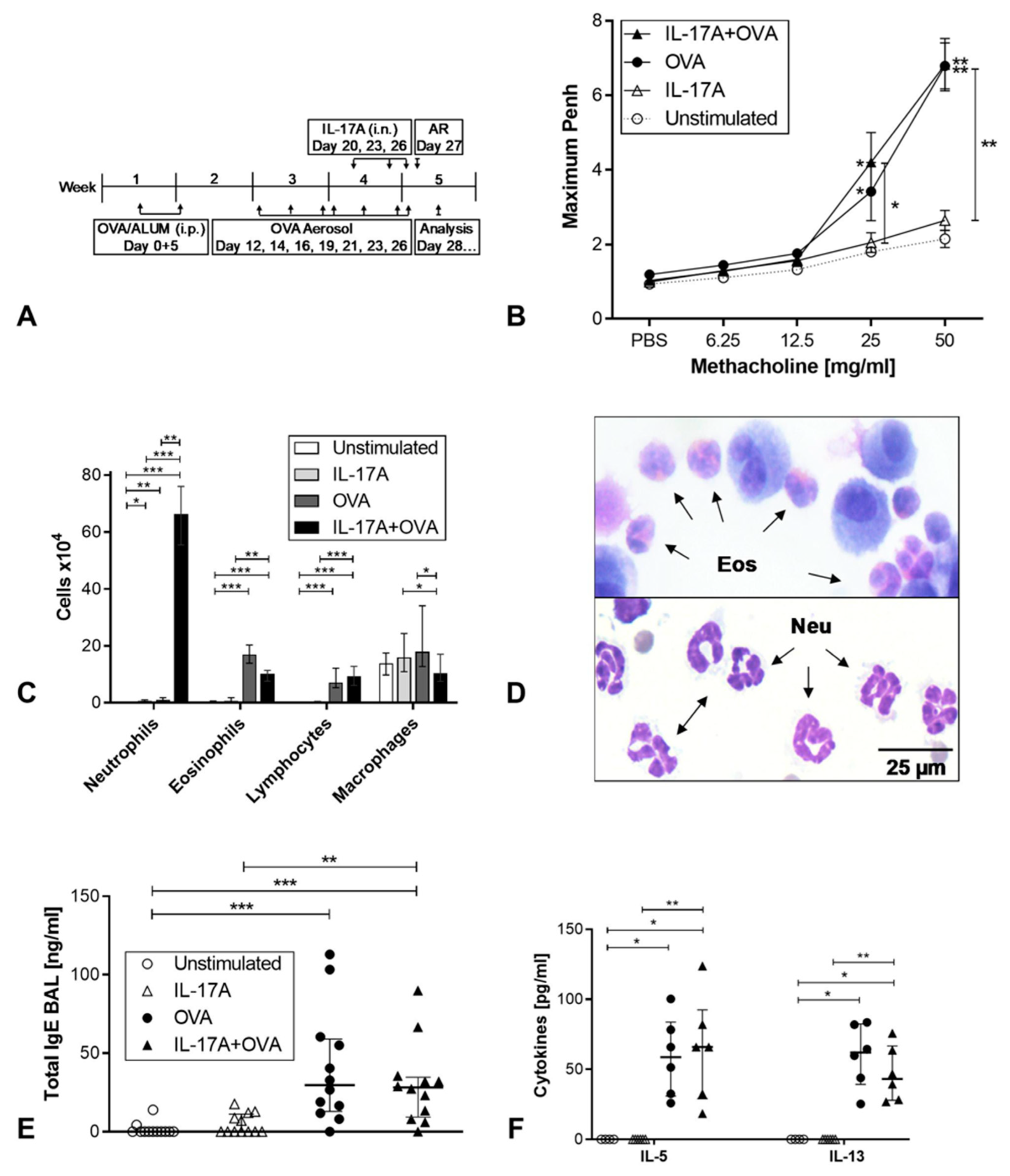
3. Results
3.1. Impact of IL-17A on Allergic Inflammation in a Mouse Model of Asthma
3.2. Analysis of Airway Remodeling
3.3. Investigation of the IL-17 Receptor Expression In Vivo and In Vitro
3.4. Investigation of IL-8 Expression by Stimulation with IL-17A and IL-13 of Human Cell Lines
4. Discussion
4.1. Influence of IL-17A on Th2-Mediated Inflammation in the OVA Mouse Model
4.2. Influence of IL-17A on Airway Remodeling in the OVA Mouse Model
4.3. Mechanisms of Action of IL-17A in the OVA Mouse Model
4.4. Significance of IL-17A for Human Lung Fibroblasts and Epithelial Cells
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fehrenbach, H.; Wagner, C.; Wegmann, M. Airway remodeling in asthma: What really matters. Cell Tissue Res. 2017, 367, 551–569. [Google Scholar] [CrossRef] [Green Version]
- Trevor, J.L.; Deshane, J.S. Refractory asthma: Mechanisms, targets, and therapy. Allergy 2014, 69, 817–827. [Google Scholar] [CrossRef]
- Acharya, K.R.; Ackerman, S.J. Eosinophil Granule Proteins: Form and Function. J. Biol. Chem. 2014, 289, 17406–17415. [Google Scholar] [CrossRef]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2018, 18, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Radermecker, C.; Sabatel, C.; Vanwinge, C.; Ruscitti, C.; Maréchal, P.; Perin, F.; Schyns, J.; Rocks, N.; Toussaint, M.; Cataldo, D.; et al. Locally instructed CXCR4hi neutrophils trigger environment-driven allergic asthma through the release of neutrophil extracellular traps. Nat. Immunol. 2019, 20, 1444–1455. [Google Scholar] [CrossRef] [PubMed]
- Newcomb, D.C.; Peebles, R.S. Th17-mediated inflammation in asthma. Curr. Opin. Immunol. 2013, 25, 755–760. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Ying, S.; Dai, Y. Pathological Roles of Neutrophil-Mediated Inflammation in Asthma and Its Potential for Therapy as a Target. J. Immunol. Res. 2017, 2017, 3743048. [Google Scholar] [CrossRef] [Green Version]
- Chesné, J.; Braza, F.; Mahay, G.; Brouard, S.; Aronica, M.; Magnan, A. IL-17 in severe asthma. Where do we stand? Am. J. Respir. Crit. Care Med. 2014, 190, 1094–1101. [Google Scholar] [CrossRef]
- Evasovic, J.M.; Singer, C.A. Regulation of IL-17A and implications for TGF-β1 comodulation of airway smooth muscle remodeling in severe asthma. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 316, L843–L868. [Google Scholar] [CrossRef]
- Kerzel, S.; Dehne, J.; Rogosch, T.; Schaub, B.; Maier, R.F.; Zemlin, M. Th17 cell frequency in peripheral blood from children with allergic asthma correlates with the level of asthma control. J. Pediatr. 2012, 161, 1172–1174. [Google Scholar] [CrossRef]
- Nieminen, K.; Valovirta, E.; Savolainen, J. Clinical outcome and IL-17, IL-23, IL-27 and FOXP3 expression in peripheral blood mononuclear cells of pollen-allergic children during sublingual immunotherapy. Pediatr. Allergy Immunol. 2010, 21, e174–e184. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, J.; Gao, Y.-D.; Guo, W. Th17 immunity in patients with allergic asthma. Int. Arch. Allergy Immunol. 2010, 151, 297–307. [Google Scholar] [CrossRef]
- Cosmi, L.; Liotta, F.; Maggi, E.; Romagnani, S.; Annunziato, F. Th17 cells: New players in asthma pathogenesis. Allergy 2011, 66, 989–998. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-H.; Voo, K.S.; Liu, B.; Chen, C.-Y.; Uygungil, B.; Spoede, W.; Bernstein, J.A.; Huston, D.P.; Liu, Y.-J. A novel subset of CD4(+) T(H)2 memory/effector cells that produce inflammatory IL-17 cytokine and promote the exacerbation of chronic allergic asthma. J. Exp. Med. 2010, 207, 2479–2491. [Google Scholar] [CrossRef] [Green Version]
- Manni, M.L.; Mandalapu, S.; McHugh, K.J.; Elloso, M.M.; Dudas, P.L.; Alcorn, J.F. Molecular Mechanisms of Airway Hyperresponsiveness in a Murine Model of Steroid-Resistant Airway Inflammation. J. Immunol. 2016, 196, 963–977. [Google Scholar] [CrossRef] [Green Version]
- Hall, S.L.; Baker, T.; Lajoie, S.; Richgels, P.K.; Yang, Y.; McAlees, J.W.; van Lier, A.; Wills-Karp, M.; Sivaprasad, U.; Acciani, T.H.; et al. IL-17A enhances IL-13 activity by enhancing IL-13-induced signal transducer and activator of transcription 6 activation. J. Allergy Clin. Immunol. 2017, 139, 462–471.e14. [Google Scholar] [CrossRef] [Green Version]
- Kinyanjui, M.W.; Shan, J.; Nakada, E.M.; Qureshi, S.T.; Fixman, E.D. Dose-dependent effects of IL-17 on IL-13-induced airway inflammatory responses and airway hyperresponsiveness. J. Immunol. 2013, 190, 3859–3868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellings, P.W.; Kasran, A.; Liu, Z.; Vandekerckhove, P.; Wuyts, A.; Overbergh, L.; Mathieu, C.; Ceuppens, J.L. Interleukin-17 orchestrates the granulocyte influx into airways after allergen inhalation in a mouse model of allergic asthma. Am. J. Respir. Cell Mol. Biol. 2003, 28, 42–50. [Google Scholar] [CrossRef]
- Seys, S.F.; Lokwani, R.; Simpson, J.L.; Bullens, D.M.A. New insights in neutrophilic asthma. Curr. Opin. Pulm. Med. 2019, 25, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Peters, M.; Köhler-Bachmann, S.; Lenz-Habijan, T.; Bufe, A. Influence of an Allergen-Specific Th17 Response on Remodeling of the Airways. Am. J. Respir. Cell Mol. Biol. 2016, 54, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Kolls, J.K. Neutrophilic Inflammation in Asthma and Association with Disease Severity. Trends Immunol. 2017, 38, 942–954. [Google Scholar] [CrossRef]
- Camargo, L.d.N.; Righetti, R.F.; Aristóteles, L.R.d.C.R.B.; Dos Santos, T.M.; de Souza, F.C.R.; Fukuzaki, S.; Cruz, M.M.; Alonso-Vale, M.I.C.; Saraiva-Romanholo, B.M.; Prado, C.M.; et al. Effects of Anti-IL-17 on Inflammation, Remodeling, and Oxidative Stress in an Experimental Model of Asthma Exacerbated by LPS. Front. Immunol. 2017, 8, 1835. [Google Scholar] [CrossRef]
- Bonser, L.R.; Erle, D.J. Airway Mucus and Asthma: The Role of MUC5AC and MUC5B. J. Clin. Med. 2017, 6, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seibold, M.A. Interleukin-13 Stimulation Reveals the Cellular and Functional Plasticity of the Airway Epithelium. Ann. Am. Thorac. Soc. 2018, 15, S98–S102. [Google Scholar] [CrossRef] [PubMed]
- Kudo, M.; Melton, A.C.; Chen, C.; Engler, M.B.; Huang, K.E.; Ren, X.; Wang, Y.; Bernstein, X.; Li, J.T.; Atabai, K.; et al. IL-17A produced by αβ T cells drives airway hyper-responsiveness in mice and enhances mouse and human airway smooth muscle contraction. Nat. Med. 2012, 18, 547–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, S.; Li, M.; Liu, B.; Ma, Z.; Yang, Q. Neutrophil extracellular traps and pulmonary fibrosis: An update. J. Inflamm. 2023, 20, 2. [Google Scholar] [CrossRef]
- Hong, C.-W. Current Understanding in Neutrophil Differentiation and Heterogeneity. Immune Netw. 2017, 17, 298–306. [Google Scholar] [CrossRef] [Green Version]
- Gaffen, S.L. Structure and signalling in the IL-17 receptor family. Nat. Rev. Immunol. 2009, 9, 556–567. [Google Scholar] [CrossRef] [Green Version]
- Goepfert, A.; Lehmann, S.; Wirth, E.; Rondeau, J.-M. The human IL-17A/F heterodimer: A two-faced cytokine with unique receptor recognition properties. Sci. Rep. 2017, 7, 8906. [Google Scholar] [CrossRef]
- Korn, T.; Bettelli, E.; Oukka, M.; Kuchroo, V.K. IL-17 and Th17 Cells. Annu. Rev. Immunol. 2009, 27, 485–517. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peters, K.; Ernst, S.; Peters, M. Interaction of Interleukin-17A with a Th2 Response in a Mouse Model of Allergic Airway Inflammation. Cells 2023, 12, 1774. https://doi.org/10.3390/cells12131774
Peters K, Ernst S, Peters M. Interaction of Interleukin-17A with a Th2 Response in a Mouse Model of Allergic Airway Inflammation. Cells. 2023; 12(13):1774. https://doi.org/10.3390/cells12131774
Chicago/Turabian StylePeters, Karin, Stefanie Ernst, and Marcus Peters. 2023. "Interaction of Interleukin-17A with a Th2 Response in a Mouse Model of Allergic Airway Inflammation" Cells 12, no. 13: 1774. https://doi.org/10.3390/cells12131774