
Retinal Injury Activates Complement Expression in Müller Cells Leading to Neuroinflammation and Photoreceptor Cell Death
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Retinal Detachment Mice Model
2.3. TUNEL Labeling and Quantitation of Photoreceptors in the Outer Nuclear Layer (ONL)
2.4. Immunostaining and Subretinal Immune Cell Counts
2.5. Flow Cytometric Analyzes of Complement Receptors
2.6. Statistical Analysis
3. Results
3.1. Müller Glial Cells Express C3 during Retinal Detachment, and Loss of C3 Protected Photoreceptor from Cell Death and Suppressed Sub-Retinal Immune Cell Infiltration

3.2. C3aR Is Upregulated in Microglia/Macrophages during RD, and Loss of C3aR Protected Photoreceptors from Cell Death and Suppressed Immune Cell Infiltration
3.3. Loss of C5-Reduced Photoreceptor Cell Death and Suppressed Immune Cell Infiltration
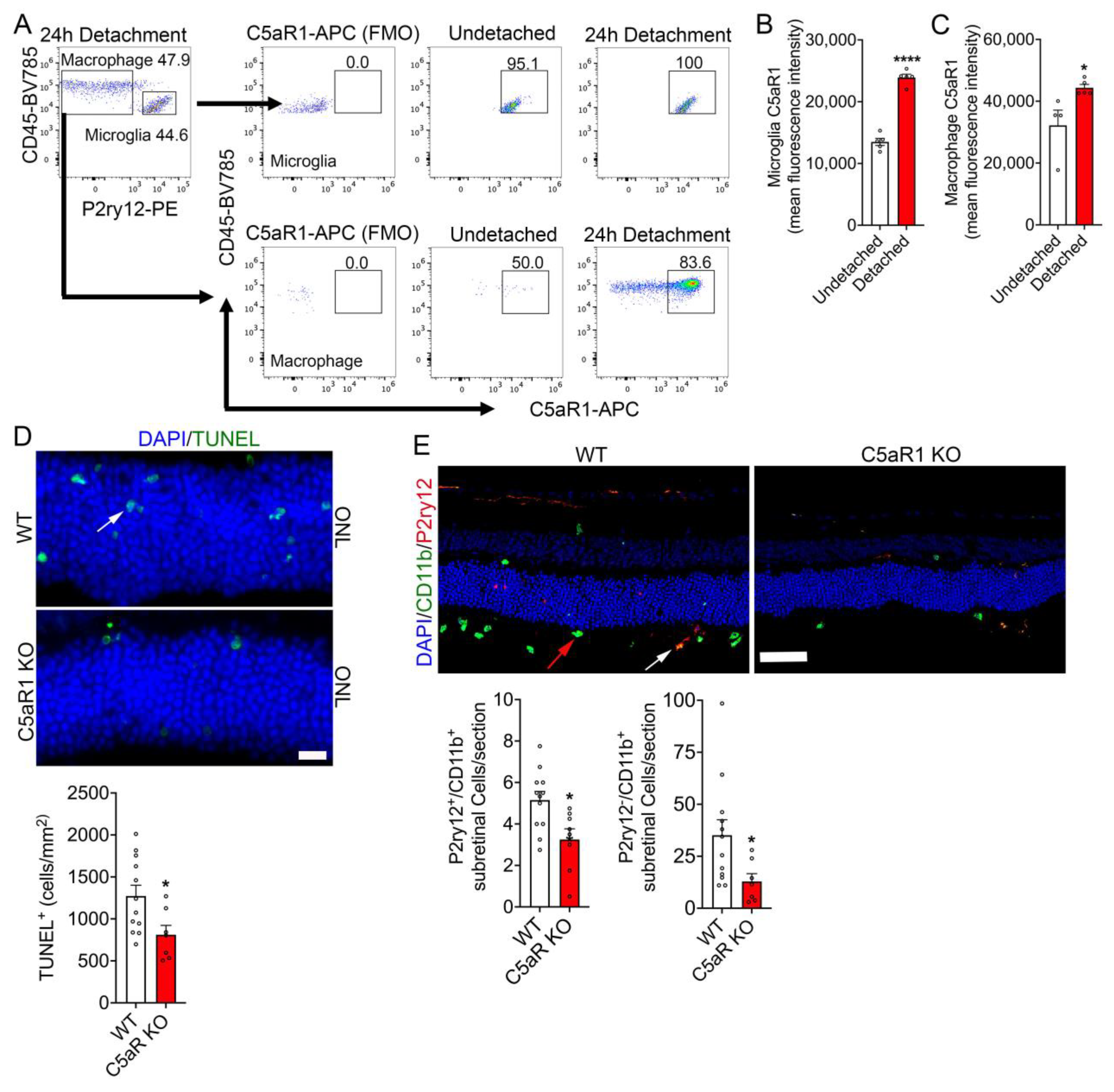
3.4. C5aR1 Is Upregulated in Microglia/Macrophages during RD, and the Loss of C5aR1 Protected Photoreceptor from Cell Death and Suppressed Immune Cell Infiltration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bird, A.C.; Bressler, N.M.; Bressler, S.B.; Chisholm, I.H.; Coscas, G.; Davis, M.D.; de Jong, P.T.; Klaver, C.C.; Klein, B.E.; Klein, R.; et al. An international classification and grading system for age-related maculopathy and age-related macular degeneration. The International ARM Epidemiological Study Group. Surv. Ophthalmol. 1995, 39, 367–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.Q.; Choudhry, N. Tractional Retinal Detachment Secondary to Diabetic Retinopathy. JAMA Ophthalmol. 2018, 136, e183507. [Google Scholar] [CrossRef] [PubMed]
- Mason, R.H.; Minaker, S.A.; Marafon, S.B.; Figueiredo, N.; Hillier, R.J.; Muni, R.H. Retinal displacement following rhegmatogenous retinal detachment: A systematic review and meta-analysis. Surv. Ophthalmol. 2022, 67, 950–964. [Google Scholar] [CrossRef] [PubMed]
- Sweigard, J.H.; Matsumoto, H.; Smith, K.E.; Kim, L.A.; Paschalis, E.I.; Okonuki, Y.; Castillejos, A.; Kataoka, K.; Hasegawa, E.; Yanai, R.; et al. Inhibition of the alternative complement pathway preserves photoreceptors after retinal injury. Sci. Transl. Med. 2015, 7, 297ra116. [Google Scholar] [CrossRef] [Green Version]
- Newton, F.; Megaw, R. Mechanisms of Photoreceptor Death in Retinitis Pigmentosa. Genes 2020, 11, 1120. [Google Scholar] [CrossRef]
- Wenzel, A.; Grimm, C.; Samardzija, M.; Reme, C.E. Molecular mechanisms of light-induced photoreceptor apoptosis and neuroprotection for retinal degeneration. Prog. Retin. Eye Res. 2005, 24, 275–306. [Google Scholar] [CrossRef]
- Manoharan, N.; Patnaik, J.L.; Olson, J.L. Increased Complement Levels in Human Vitreous Aspirates of Proliferative Diabetic Retinopathy and Retinal Detachment Eyes. Retina 2019, 39, 2212–2218. [Google Scholar] [CrossRef]
- Ricklin, D.; Hajishengallis, G.; Yang, K.; Lambris, J.D. Complement: A key system for immune surveillance and homeostasis. Nat. Immunol. 2010, 11, 785–797. [Google Scholar] [CrossRef] [Green Version]
- Merle, N.S.; Church, S.E.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement System Part I—Molecular Mechanisms of Activation and Regulation. Front. Immunol. 2015, 6, 262. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Lennon, V.A.; Liu, Y.U.; Bosco, D.B.; Li, Y.; Yi, M.H.; Zhu, J.; Wei, S.; Wu, L.J. Astrocyte-microglia interaction drives evolving neuromyelitis optica lesion. J. Clin. Investig. 2020, 130, 4025–4038. [Google Scholar] [CrossRef]
- Friscic, J.; Bottcher, M.; Reinwald, C.; Bruns, H.; Wirth, B.; Popp, S.J.; Walker, K.I.; Ackermann, J.A.; Chen, X.; Turner, J.; et al. The complement system drives local inflammatory tissue priming by metabolic reprogramming of synovial fibroblasts. Immunity 2021, 54, 1002–1021 e1010. [Google Scholar] [CrossRef] [PubMed]
- Osaka, M.; Ito, S.; Honda, M.; Inomata, Y.; Egashira, K.; Yoshida, M. Critical role of the C5a-activated neutrophils in high-fat diet-induced vascular inflammation. Sci. Rep. 2016, 6, 21391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Werkhoven, M.B.; Damman, J.; van Dijk, M.; Daha, M.R.; de Jong, I.J.; Leliveld, A.; Krikke, C.; Leuvenink, H.G.; van Goor, H.; van Son, W.J.; et al. Complement mediated renal inflammation induced by donor brain death: Role of renal C5a-C5aR interaction. Am. J. Transpl. 2013, 13, 875–882. [Google Scholar] [CrossRef]
- Song, W.C. Membrane complement regulatory proteins in autoimmune and inflammatory tissue injury. Curr. Dir. Autoimmun. 2004, 7, 181–199. [Google Scholar] [CrossRef]
- Kolev, M.; West, E.E.; Kunz, N.; Chauss, D.; Moseman, E.A.; Rahman, J.; Freiwald, T.; Balmer, M.L.; Lotscher, J.; Dimeloe, S.; et al. Diapedesis-Induced Integrin Signaling via LFA-1 Facilitates Tissue Immunity by Inducing Intrinsic Complement C3 Expression in Immune Cells. Immunity 2020, 52, 513–527.e518. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Miller, J.W.; Vavvas, D.G. Retinal detachment model in rodents by subretinal injection of sodium hyaluronate. J. Vis. Exp. 2013, 79, e50660. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, H.; Kataoka, K.; Tsoka, P.; Connor, K.M.; Miller, J.W.; Vavvas, D.G. Strain difference in photoreceptor cell death after retinal detachment in mice. Invest. Ophthalmol. Vis. Sci. 2014, 55, 4165–4174. [Google Scholar] [CrossRef] [Green Version]
- Gnanaguru, G.; Tabor, S.J.; Bonilla, G.M.; Sadreyev, R.; Yuda, K.; Kohl, J.; Connor, K.M. Microglia refine developing retinal astrocytic and vascular networks through the complement C3/C3aR axis. Development 2023, 150, dev201047. [Google Scholar] [CrossRef]
- Okunuki, Y.; Tabor, S.J.; Lee, M.Y.; Connor, K.M. CD47 Deficiency Ameliorates Ocular Autoimmune Inflammation. Front. Immunol. 2021, 12, 680568. [Google Scholar] [CrossRef]
- Luo, C.; Chen, M.; Xu, H. Complement gene expression and regulation in mouse retina and retinal pigment epithelium/choroid. Mol. Vis. 2011, 17, 1588–1597. [Google Scholar]
- Okunuki, Y.; Mukai, R.; Pearsall, E.A.; Klokman, G.; Husain, D.; Park, D.H.; Korobkina, E.; Weiner, H.L.; Butovsky, O.; Ksander, B.R.; et al. Microglia inhibit photoreceptor cell death and regulate immune cell infiltration in response to retinal detachment. Proc. Natl. Acad. Sci. USA 2018, 115, E6264–E6273. [Google Scholar] [CrossRef] [Green Version]
- Peng, Q.; Li, K.; Sacks, S.H.; Zhou, W. The role of anaphylatoxins C3a and C5a in regulating innate and adaptive immune responses. Inflamm. Allergy-Drug Targets 2009, 8, 236–246. [Google Scholar] [CrossRef]
- Wu, F.; Zou, Q.; Ding, X.; Shi, D.; Zhu, X.; Hu, W.; Liu, L.; Zhou, H. Complement component C3a plays a critical role in endothelial activation and leukocyte recruitment into the brain. J. Neuroinflamm. 2016, 13, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karasu, E.; Demmelmaier, J.; Kellermann, S.; Holzmann, K.; Kohl, J.; Schmidt, C.Q.; Kalbitz, M.; Gebhard, F.; Huber-Lang, M.S.; Halbgebauer, R. Complement C5a Induces Pro-inflammatory Microvesicle Shedding in Severely Injured Patients. Front. Immunol. 2020, 11, 1789. [Google Scholar] [CrossRef] [PubMed]
- Linsenmeier, R.A.; Padnick-Silver, L. Metabolic dependence of photoreceptors on the choroid in the normal and detached retina. Invest. Ophthalmol. Vis. Sci. 2000, 41, 3117–3123. [Google Scholar] [PubMed]
- Lewis, G.; Mervin, K.; Valter, K.; Maslim, J.; Kappel, P.J.; Stone, J.; Fisher, S. Limiting the proliferation and reactivity of retinal Muller cells during experimental retinal detachment: The value of oxygen supplementation. Am. J. Ophthalmol. 1999, 128, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Shelby, S.J.; Angadi, P.S.; Zheng, Q.D.; Yao, J.; Jia, L.; Zacks, D.N. Hypoxia inducible factor 1alpha contributes to regulation of autophagy in retinal detachment. Exp. Eye Res. 2015, 137, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Nakazawa, T.; Takeda, M.; Lewis, G.P.; Cho, K.S.; Jiao, J.; Wilhelmsson, U.; Fisher, S.K.; Pekny, M.; Chen, D.F.; Miller, J.W. Attenuated glial reactions and photoreceptor degeneration after retinal detachment in mice deficient in glial fibrillary acidic protein and vimentin. Invest. Ophthalmol. Vis. Sci. 2007, 48, 2760–2768. [Google Scholar] [CrossRef] [Green Version]
- Bringmann, A.; Pannicke, T.; Grosche, J.; Francke, M.; Wiedemann, P.; Skatchkov, S.N.; Osborne, N.N.; Reichenbach, A. Muller cells in the healthy and diseased retina. Prog. Retin. Eye Res. 2006, 25, 397–424. [Google Scholar] [CrossRef]
- Morshedian, A.; Kaylor, J.J.; Ng, S.Y.; Tsan, A.; Frederiksen, R.; Xu, T.; Yuan, L.; Sampath, A.P.; Radu, R.A.; Fain, G.L.; et al. Light-Driven Regeneration of Cone Visual Pigments through a Mechanism Involving RGR Opsin in Muller Glial Cells. Neuron 2019, 102, 1172–1183.e1175. [Google Scholar] [CrossRef]
- Gu, L.; Xu, H.; Zhang, C.; Yang, Q.; Zhang, L.; Zhang, J. Time-dependent changes in hypoxia- and gliosis-related factors in experimental diabetic retinopathy. Eye 2019, 33, 600–609. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, H.; Sugio, S.; Seghers, F.; Krizaj, D.; Akiyama, H.; Ishizaki, Y.; Gailly, P.; Shibasaki, K. Retinal Detachment-Induced Muller Glial Cell Swelling Activates TRPV4 Ion Channels and Triggers Photoreceptor Death at Body Temperature. J. Neurosci. 2018, 38, 8745–8758. [Google Scholar] [CrossRef] [Green Version]
- West, E.E.; Kemper, C. Complosome—The intracellular complement system. Nat. Rev. Nephrol. 2023, 19, 426–439. [Google Scholar] [CrossRef]
- Schartz, N.D.; Tenner, A.J. The good, the bad, and the opportunities of the complement system in neurodegenerative disease. J. Neuroinflamm. 2020, 17, 354. [Google Scholar] [CrossRef] [PubMed]
- Kastl, S.P.; Speidl, W.S.; Kaun, C.; Katsaros, K.M.; Rega, G.; Afonyushkin, T.; Bochkov, V.N.; Valent, P.; Assadian, A.; Hagmueller, G.W.; et al. In human macrophages the complement component C5a induces the expression of oncostatin M via AP-1 activation. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvelli, J.; Demaria, O.; Vely, F.; Batista, L.; Chouaki Benmansour, N.; Fares, J.; Carpentier, S.; Thibult, M.L.; Morel, A.; Remark, R.; et al. Association of COVID-19 inflammation with activation of the C5a-C5aR1 axis. Nature 2020, 588, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.Y.; Xu, F.; Yang, Y.; Sun, D.D.; Zhong, Y.; Wu, S.; Li, G.Q.; Gao, W.W.; Wang, T.; Xu, G.L.; et al. C5a/C5aR1 mediates IMQ-induced psoriasiform skin inflammation by promoting IL-17A production from gammadelta-T cells. FASEB J. 2020, 34, 10590–10604. [Google Scholar] [CrossRef]
- Carvalho, K.; Schartz, N.D.; Balderrama-Gutierrez, G.; Liang, H.Y.; Chu, S.H.; Selvan, P.; Gomez-Arboledas, A.; Petrisko, T.J.; Fonseca, M.I.; Mortazavi, A.; et al. Modulation of C5a-C5aR1 signaling alters the dynamics of AD progression. J. Neuroinflamm. 2022, 19, 178. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tabor, S.J.; Yuda, K.; Deck, J.; Gnanaguru, G.; Connor, K.M. Retinal Injury Activates Complement Expression in Müller Cells Leading to Neuroinflammation and Photoreceptor Cell Death. Cells 2023, 12, 1754. https://doi.org/10.3390/cells12131754
Tabor SJ, Yuda K, Deck J, Gnanaguru G, Connor KM. Retinal Injury Activates Complement Expression in Müller Cells Leading to Neuroinflammation and Photoreceptor Cell Death. Cells. 2023; 12(13):1754. https://doi.org/10.3390/cells12131754
Chicago/Turabian StyleTabor, Steven J., Kentaro Yuda, Jonathan Deck, Gopalan Gnanaguru, and Kip M. Connor. 2023. "Retinal Injury Activates Complement Expression in Müller Cells Leading to Neuroinflammation and Photoreceptor Cell Death" Cells 12, no. 13: 1754. https://doi.org/10.3390/cells12131754