

Rlip Reduction Induces Oxidative Stress and Mitochondrial Dysfunction in Mutant Tau-Expressed Immortalized Hippocampal Neurons: Mechanistic Insights
 , , , , and
, , , , and
Abstract
:1. Introduction
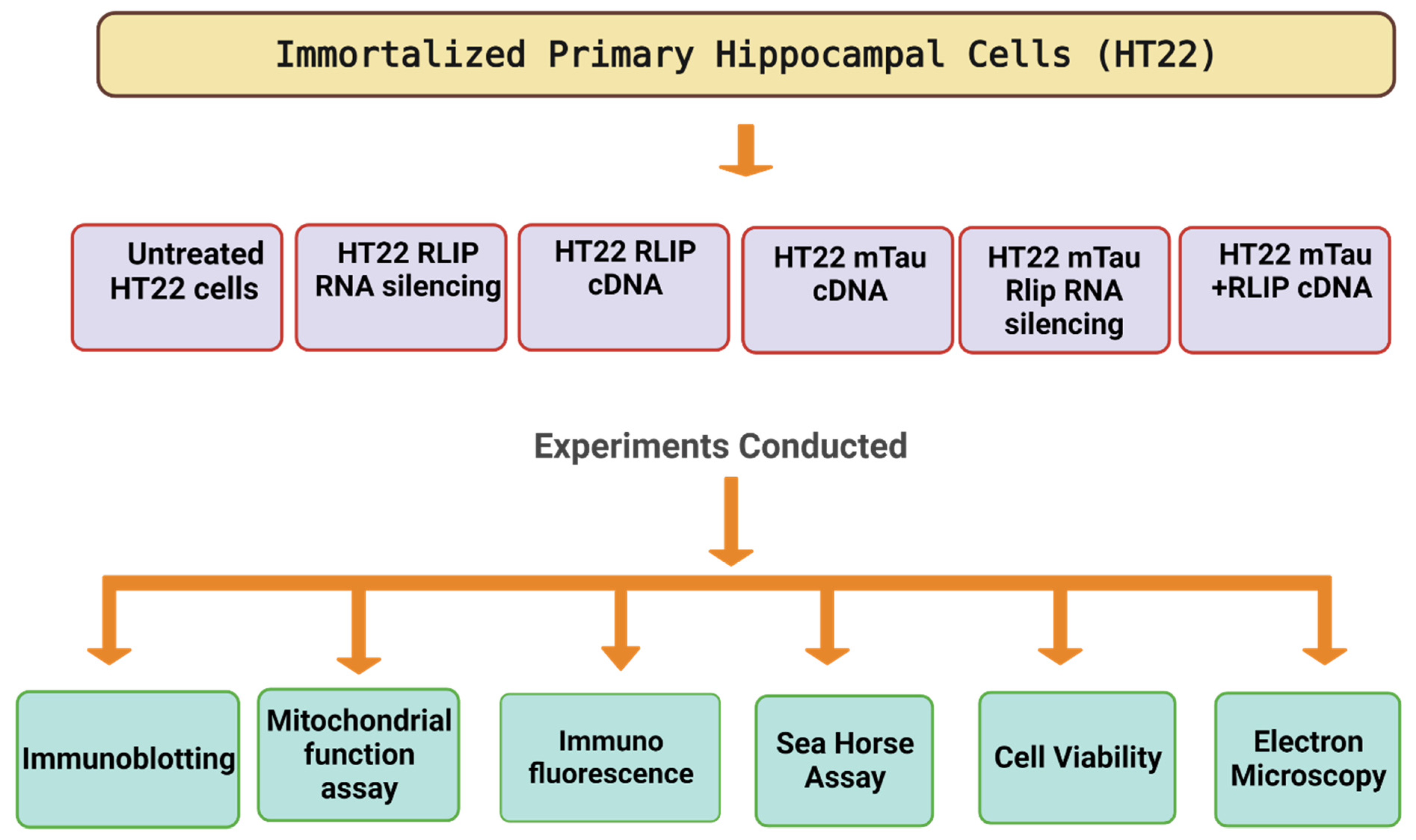
2. Material and Methods
2.1. Mutant Tau cDNA Constructs
2.2. Tissue Culture Work
2.3. mTau Overexpression and Rlip Depletion by siRNA
2.4. Cell Survival Assay
2.5. Mitochondrial Respiration Using Seahorse XFe96 Extracellular Flux Analyzer
2.6. Mitochondrial Function Analysis
2.7. Western Blot Analysis
2.8. Immunofluorescence Analysis
2.9. Transmission Electron Microscopy
2.10. Statistical Analysis
3. Results
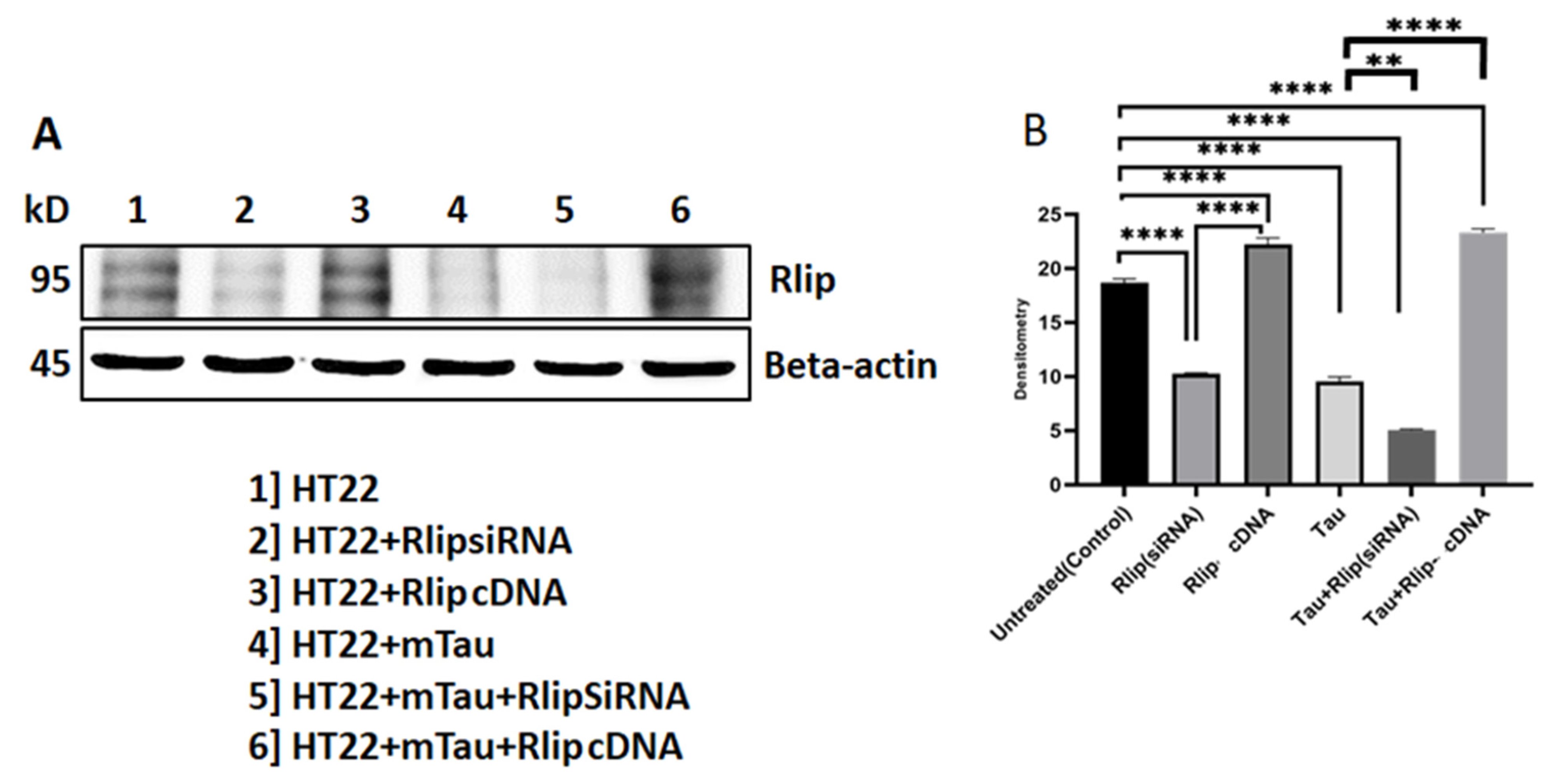
3.1. Immunoblotting Analysis of Rlip Protein
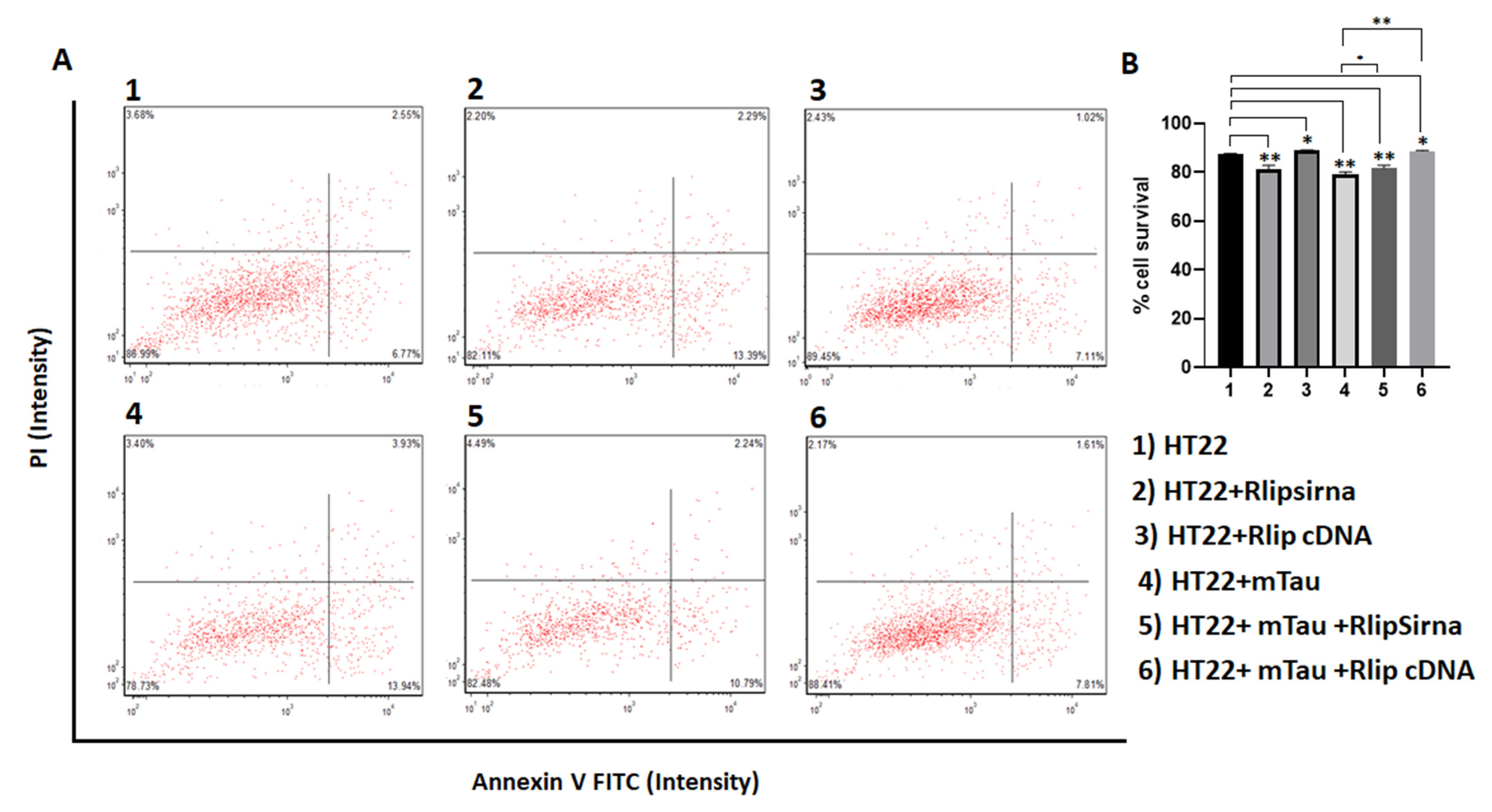
3.2. Cell Survival Assays in HT22 Cells and HT22 Cells Transfected with Mutant Tau cDNA and Rlip Overexpression and Silencing
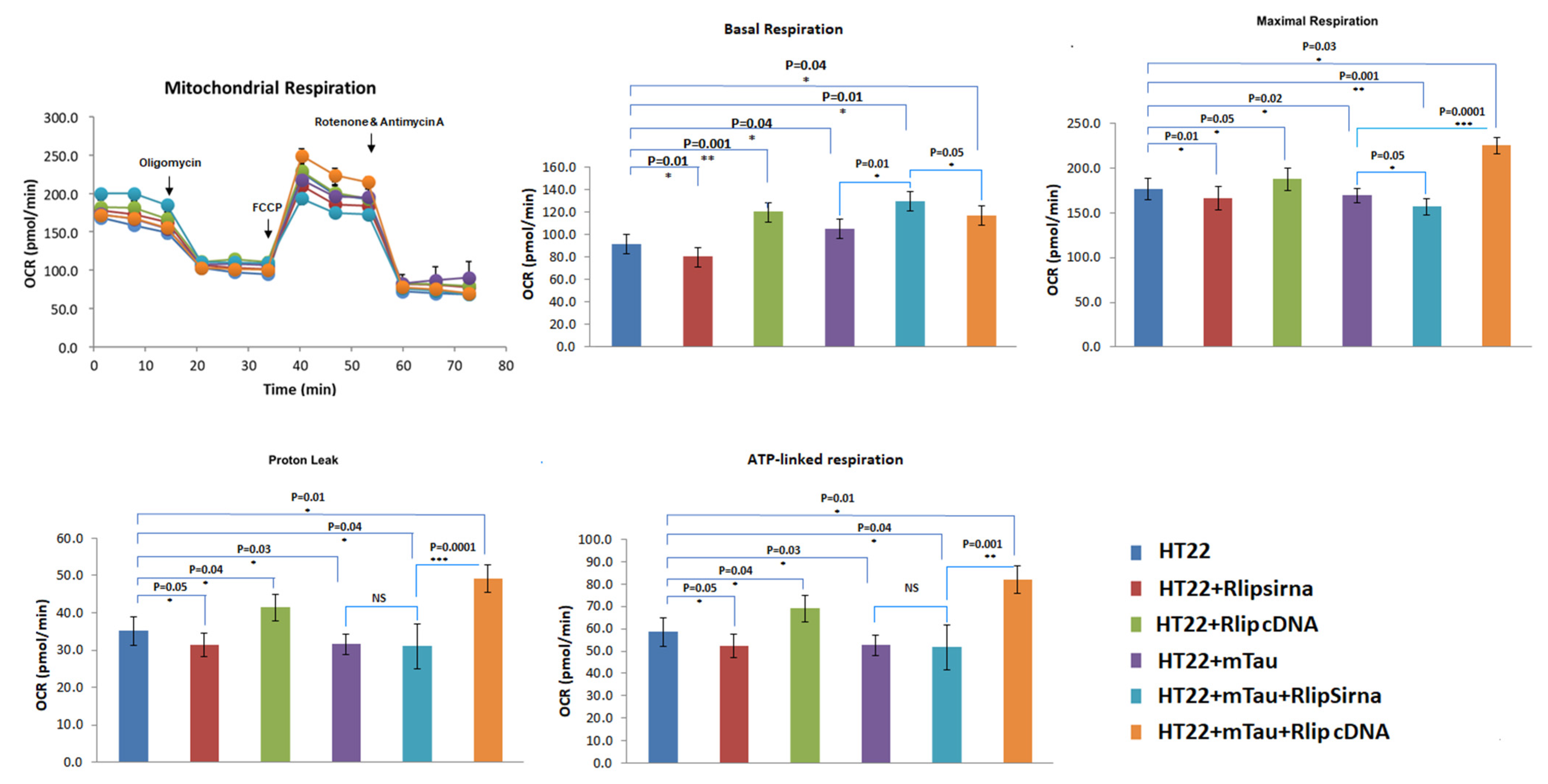
3.3. Mitochondrial Respiration
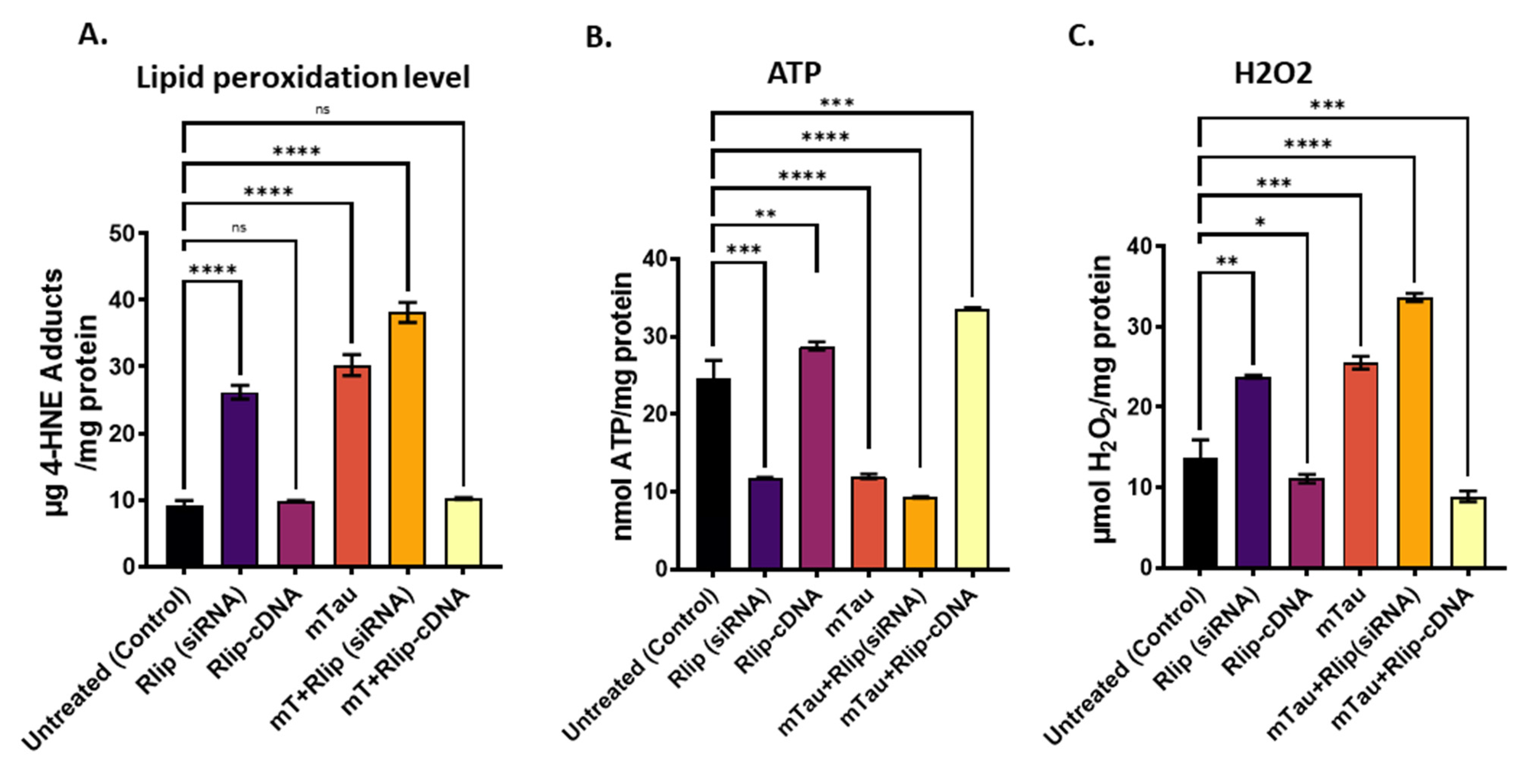
3.4. Mitochondrial Functional Analysis in HT22 Cells
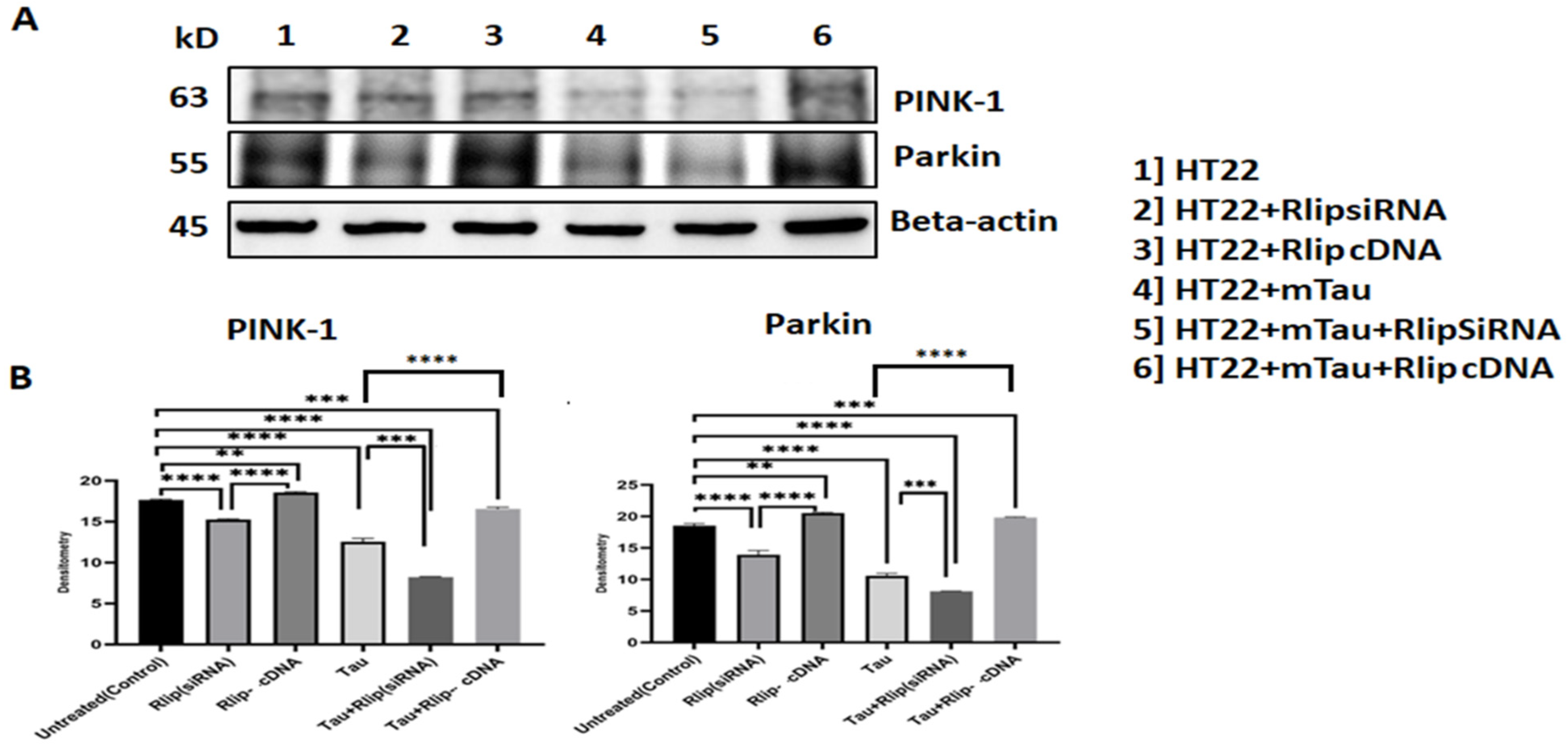
3.5. Immunoblotting Analysis of Mitophagy Proteins
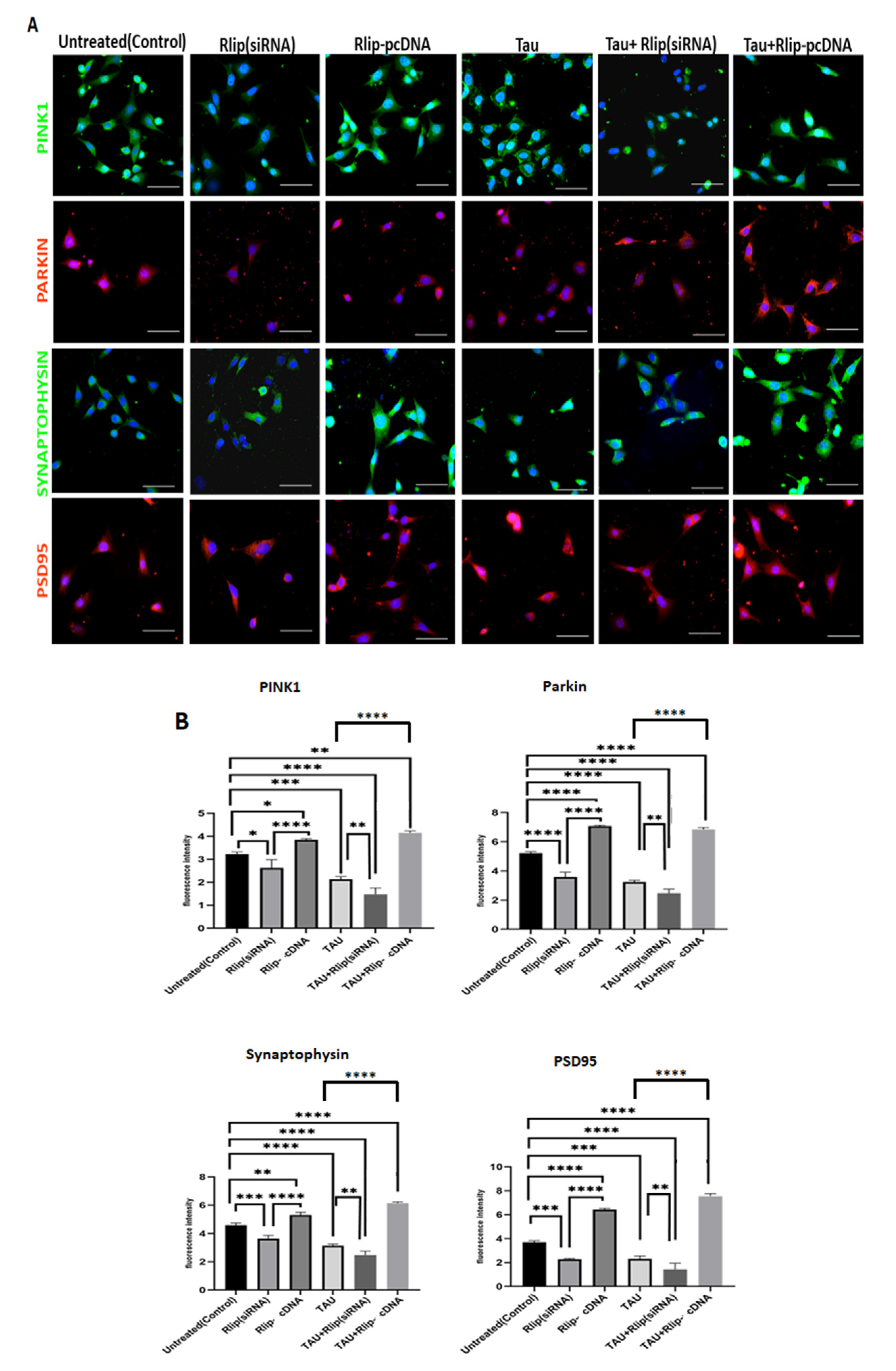
3.6. Immunofluorescence Analysis of Mitophagy and Synaptic Proteins
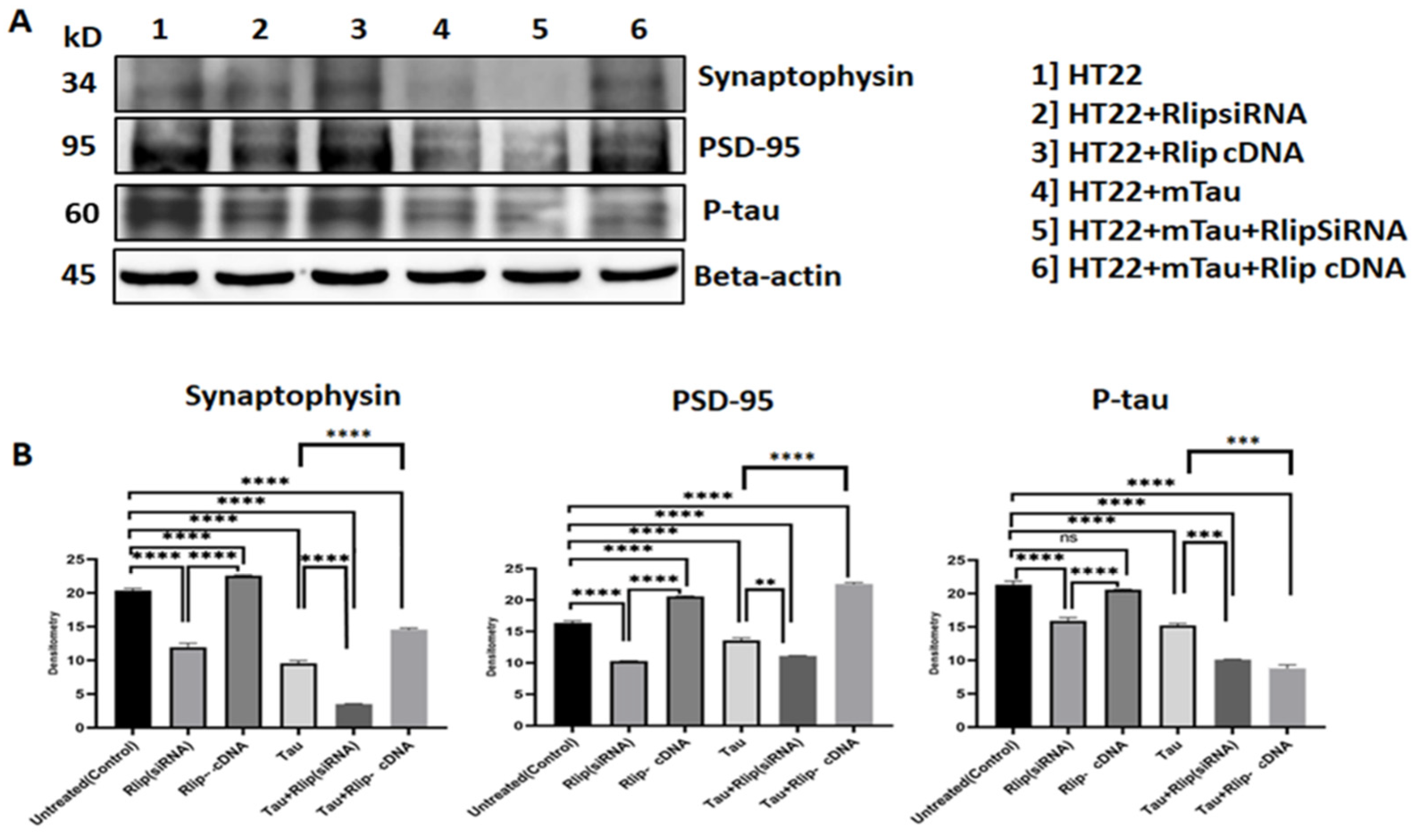
3.7. Immunoblotting Analysis of Synaptic Proteins
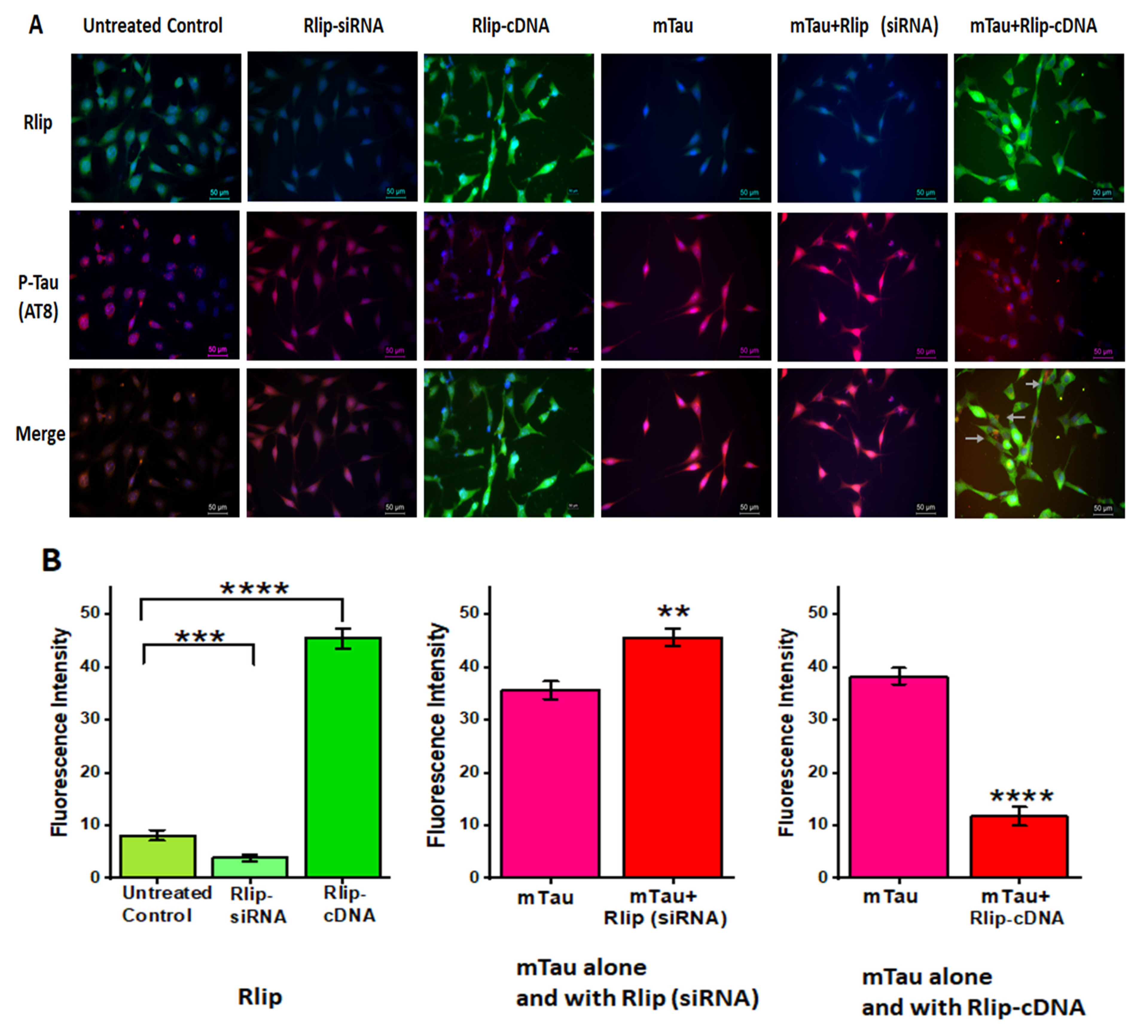
3.8. Immunofluorescence Analysis and Colocalization of RLIP and Mutant Tau Proteins
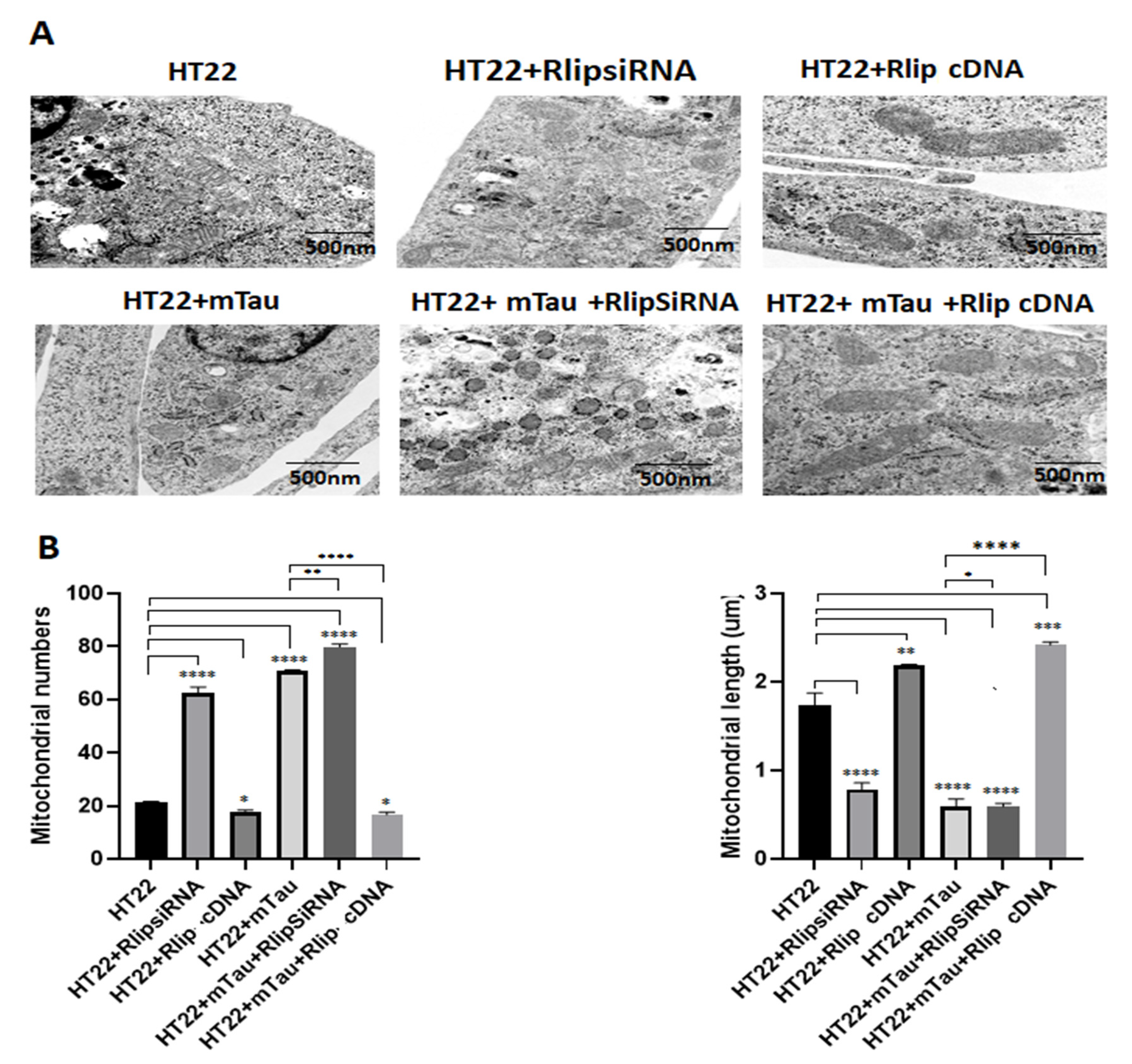
3.9. Transmission Electron Microscopy Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- John, A.; Reddy, P.H. Synaptic basis of Alzheimer’s disease: Focus on synaptic amyloid beta, P-tau and mitochondria. Ageing Res. Rev. 2021, 65, 101208. [Google Scholar] [CrossRef]
- Mao, P.; Manczak, M.; Shirendeb, U.P.; Reddy, P.H. MitoQ, a mitochondria-targeted antioxidant, delays disease progression and alleviates pathogenesis in an experimental autoimmune encephalomyelitis mouse model of multiple sclerosis. Biochim. Biophys. Acta 2013, 1832, 2322–2331. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J. Alzheimer’s disease: Genes, proteins, and therapy. Physiol. Rev. 2001, 81, 741–766. [Google Scholar] [CrossRef]
- Mattson, M.P. Pathways towards and away from Alzheimer’s disease. Nature 2004, 430, 631–639. [Google Scholar] [CrossRef]
- Swerdlow, R.H.; Burns, J.M.; Khan, S.M. The Alzheimer’s disease mitochondrial cascade hypothesis. J. Alzheimer’s Dis. 2010, 2, S265–S279. [Google Scholar] [CrossRef] [PubMed]
- Wyss-Coray, T.; Mucke, L. Inflammation in Neurodegenerative Disease—A Double-Edged Sword. Neuron 2002, 35, 419–432. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.H.; Beal, M.F. Amyloid beta, mitochondrial dysfunction and synaptic damage: Implications for cognitive decline in aging and Alzheimer’s disease. Trends Mol. Med. 2008, 2, 45–53. [Google Scholar] [CrossRef]
- Rawat, P.; Sehar, U.; Bisht, J.; Selman, A.; Culberson, J.; Reddy, P.H. Phosphorylated Tau in Alzheimer’s Disease and Other Tauopathies. Int. J. Mol. Sci. 2022, 23, 12841. [Google Scholar] [CrossRef]
- Kumar, S.; Reddy, P.H. Are circulating microRNAs peripheral biomarkers for Alzheimer’s disease? Biochim. Biophys. Acta 2016, 1862, 1617–1627. [Google Scholar] [CrossRef]
- Reddy, P.H.; Tonk, S.; Kumar, S.; Vijayan, M.; Kandimalla, R.; Kuruva, C.S.; Reddy, A.P. A critical evaluation of neuroprotective and neurodegenerative MicroRNAs in Alzheimer’s disease. Biochem. Biophys. Res. Commun. 2017, 483, 1156–1165. [Google Scholar] [CrossRef]
- Vijayan, M.; Reddy, P.H. Stroke, Vascular Dementia, and Alzheimer’s Disease: Molecular Links. J. Alzheimer’s Dis. 2016, 54, 427–443. [Google Scholar] [CrossRef]
- Wang, W.; Zhao, F.; Ma, X.; Perry, G.; Zhu, X. Mitochondria dysfunction in the pathogenesis of Alzheimer’s disease: Recent advances. Mol. Neurodegener. 2020, 15, 30. [Google Scholar] [CrossRef] [PubMed]
- Morton, H.; Kshirsagar, S.; Orlov, E.; Bunquin, L.E.; Sawant, N.; Boleng, L.; George, M.; Basu, T.; Ramasubramanian, B.; Pradeepkiran, J.A.; et al. Defective mitophagy and synaptic degeneration in Alzheimer’s disease: Focus on aging, mitochondria and synapse. Free Radic. Biol. Med. 2021, 172, 652–667. [Google Scholar] [CrossRef] [PubMed]
- Pradeepkiran, J.A.; Baig, J.; Selman, A.; Reddy, P.H. Mitochondria in Aging and Alzheimer’s Disease: Focus on Mitophagy. Neuroscientist 2023. [Google Scholar] [CrossRef]
- Dewanjee, S.; Chakraborty, P.; Bhattacharya, H.; Chacko, L.; Singh, B.; Chaudhary, A.; Javvaji, K.; Pradhan, S.R.; Vallamkondu, J.; Dey, A.; et al. Altered glucose metabolism in Alzheimer’s disease: Role of mitochondrial dysfunction and oxidative stress. Free Radic. Biol. Med. 2022, 193, 134–157. [Google Scholar] [CrossRef]
- Reddy, P.H. Abnormal tau, mitochondrial dysfunction, impaired axonal transport of mitochondria, and synaptic deprivation in Alzheimer’s disease. Brain Res. 2011, 1415, 136–148. [Google Scholar] [CrossRef]
- Lee, V.M.; Goedert, M.; Trojanowski, J.Q. Neurodegenerative tauopathies. Annu. Rev. Neurosci. 2001, 24, 1121–1159. [Google Scholar] [CrossRef] [PubMed]
- Avila, J.; Pérez, M.; Lim, F.; Gómez-Ramos, A.; Hernández, F.; Lucas, J.J. Tau in neurodegenerative diseases: Tau phosphorylation and assembly. Neurotox. Res. 2004, 6, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Querfurth, H.W.; LaFerla, F.M. Alzheimer’s disease. N. Engl. J. Med. 2010, 362, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, K.; Alonso, A.d.C.; Chen, S.; Chohan, M.O.; El-Akkad, E.; Gong, C.-X.; Khatoon, S.; Li, B.; Liu, F.; Rahman, A.; et al. Tau pathology in Alzheimer disease and other tauopathies. Biochim. Biophys. Acta 2005, 1739, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Ittner, L.M.; Götz, J. Amyloid-β and tau--a toxic pas de deux in Alzheimer’s disease. Nat. Rev. Neurosci. 2011, 12, 65–72. [Google Scholar] [CrossRef]
- Sharma, R.; Gupta, S.; Singh, S.V.; Medh, R.D.; Ahmad, H.; LaBelle, E.F.; Awasthi, Y.C. Purification and characterization of dinitrophenylglutathione ATPase of human erythrocytes and its expression in other tissues. Biochem. Biophys. Res. Commun. 1990, 171, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, S.; Cheng, J.-Z.; Singhal, S.S.; Pandya, U.; Sharma, R.; Singh, S.V.; Zimniak, P.; Awasthi, Y.C. Functional Reassembly of ATP-Dependent Xenobiotic Transport by the N- and C-Terminal Domains of RLIP76 and Identification of ATP Binding Sequences. Biochemistry 2001, 40, 4159–4168. [Google Scholar] [CrossRef] [PubMed]
- Singhal, S.S.; Singhal, J.; Cheng, J.; Pikuła, S.; Sharma, R.; Zimniak, P.; Awasthi, Y.C.; Awasthi, S. Purification and functional reconstitution of intact ral-binding Gtpase activating protein, RLIP76, in artificial liposomes. Acta Biochim. Pol. 2001, 48, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Singhal, J.; Nagaprashantha, L.; Vatsyayan, R.; Awasthi, S.; Singhal, S.S. RLIP76, a Glutathione-Conjugate Transporter, Plays a Major Role in the Pathogenesis of Metabolic Syndrome. PLoS ONE 2011, 6, e24688. [Google Scholar] [CrossRef] [PubMed]
- Singhal, S.S.; Garg, R.; Horne, D.; Singhal, S.; Awasthi, S.; Salgia, R. RLIP: A necessary transporter protein for translating oxidative stress into pro-obesity and pro-carcinogenic signaling. Biochim. Biophys. Acta Rev. Cancer 2022, 1877, 188803. [Google Scholar] [CrossRef]
- Hindle, A.; Singh, S.P.; Pradeepkiran, J.A.; Bose, C.; Vijayan, M.; Kshirsagar, S.; Sawant, N.A.; Reddy, P.H. Rlip76: An Unexplored Player in Neurodegeneration and Alzheimer’s Disease? Int. J. Mol. Sci. 2022, 23, 6098. [Google Scholar] [CrossRef]
- Awasthi, S.; Hindle, A.; Sawant, N.A.; George, M.; Vijayan, M.; Kshirsagar, S.; Morton, H.; Bunquin, L.E.; Palade, P.T.; Lawrence, J.J.; et al. RALBP1 in Oxidative Stress and Mitochondrial Dysfunction in Alzheimer’s Disease. Cells 2021, 10, 3113. [Google Scholar] [CrossRef]
- Peigneux, P.; Laureys, S.; Fuchs, S.; Collette, F.; Perrin, F.; Reggers, J.; Phillips, C.; Degueldre, C.; Del Fiore, G.; Aerts, J.; et al. Are spatial memories strengthened in the human hippocampus during slow wave sleep? Neuron 2004, 44, 535–545. [Google Scholar] [CrossRef]
- Maguire, E.A.; Mummery, C.J.; Büchel, C. Patterns of hippocampal-cortical interaction dissociate temporal lobe memory subsystems. Hippocampus 2000, 10, 475–482. [Google Scholar] [CrossRef]
- Davis, J.B.; Maher, P. Protein kinase C activation inhibits glutamate-induced cytotoxicity in a neuronal cell line. Brain Res. 1994, 652, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, B.H.; Koshland, D.E., Jr. Excitatory amino acid uptake and N-methyl-D-aspartate-mediated secretion in a neural cell line. Proc. Natl. Acad. Sci. USA 1990, 87, 3518–3521. [Google Scholar] [CrossRef] [PubMed]
- Kshirsagar, S.; Sawant, N.; Morton, H.; Reddy, A.P.; Reddy, P.H. Mitophagy enhancers against phosphorylated Tau-induced mitochondrial and synaptic toxicities in Alzheimer disease. Pharmacol. Res. 2021, 174, 105973. [Google Scholar] [CrossRef]
- Kandimalla, R.; Manczak, M.; Yin, X.; Wang, R.; Reddy, P.H. Hippocampal phosphorylated tau induced cognitive decline, dendritic spine loss and mitochondrial abnormalities in a mouse model of Alzheimer’s disease. Hum. Mol. Genet. 2018, 27, 30–40. [Google Scholar] [CrossRef]
- Dubey, M.; Chaudhury, P.; Kabiru, H.; Shea, T.B. Tau inhibits anterograde axonal transport and perturbs stability in growing axonal neurites in part by displacing kinesin cargo: Neurofilaments attenuate tau-mediated neurite instability. Cell Motil. Cytoskelet. 2007, 65, 89–99. [Google Scholar] [CrossRef]
- Dixit, R.; Ross, J.L.; Goldman, Y.E.; Holzbaur, E.L.F. Differential Regulation of Dynein and Kinesin Motor Proteins by Tau. Science 2008, 319, 1086–1089. [Google Scholar] [CrossRef]
- Baas, P.W.; Qiang, L. Neuronal microtubules: When the MAP is the roadblock. Trends Cell Biol. 2005, 15, 183–187. [Google Scholar] [CrossRef]
- Pinto-Almazan, R.; Segura-Uribe, J.J.; A Soriano-Ursúa, M.; Farfán-García, E.D.; Gallardo, J.M.; Guerra-Araiza, C. Effect of tibolone pretreatment on kinases and phosphatases that regulate the expression and phosphorylation of Tau in the hippocampus of rats exposed to ozone. Neural. Regen. Res. 2018, 13, 440–448. [Google Scholar] [CrossRef]
- Reddy, P.H.; Oliver, D.M. Amyloid Beta and Phosphorylated Tau-Induced Defective Autophagy and Mitophagy in Alzheimer’s Disease. Cells 2019, 8, 488. [Google Scholar] [CrossRef]
- dos Santos, A.; Teixeira, F.C.; da Silva, D.S.; Veleda, T.A.; de Mello, J.E.; Luduvico, K.P.; Tavares, R.G.; Stefanello, F.M.; Cunico, W.; Spanevello, R.M. Thiazolidin-4-one prevents against memory deficits, increase in phosphorylated tau protein, oxidative damage and cholinergic dysfunction in Alzheimer disease model: Comparison with donepezil drug. Brain Res. Bull. 2023, 193, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Manczak, M.; Reddy, P.H. Abnormal interaction of VDAC1 with amyloid beta and phosphorylated tau causes mitochondrial dysfunction in Alzheimer’s disease. Hum. Mol. Genet. 2012, 21, 5131–5146. [Google Scholar] [CrossRef] [PubMed]
- Trease, A.J.; George, J.W.; Roland, N.J.; Lichter, E.Z.; Emanuel, K.; Totusek, S.; Fox, H.S.; Stauch, K.L. Hyperphosphorylated Human Tau Accumulates at the Synapse, Localizing on Synaptic Mitochondrial Outer Membranes and Disrupting Respiration in a Mouse Model of Tauopathy. Front. Mol. Neurosci. 2022, 15, 852368. [Google Scholar] [CrossRef] [PubMed]
- Christensen, K.R.; Combs, B.; Richards, C.; Grabinski, T.; Alhadidy, M.M.; Kanaan, N.M. Phosphomimetics at Ser199/Ser202/Thr205 in Tau Impairs Axonal Transport in Rat Hippocampal Neurons. Mol. Neurobiol. 2023, 60, 3423–3438. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Bai, F. The Association of Tau with Mitochondrial Dysfunction in Alzheimer’s Disease. Front Neurosci. 2018, 12, 163. [Google Scholar] [CrossRef] [PubMed]
- Butler, T.; Bowen, R.; Atwood, C.S. Septal hypertrophy and cell cycle re-entry in, A.D. Aging 2019, 11, 297–298. [Google Scholar] [CrossRef] [PubMed]
- de la Torre, J.C. Deciphering Alzheimer’s Disease Pathogenic Pathway: Role of Chronic Brain Hypoperfusion on p-Tau and mTOR. J. Alzheimer’s Dis. 2021, 79, 1381–1396. [Google Scholar] [CrossRef]
- Wang, S.; Chen, X.; Crisman, L.; Dou, X.; Winborn, C.S.; Wan, C.; Puscher, H.; Yin, Q.; Kennedy, M.J.; Shen, J. Regulation of cargo exocytosis by a Reps1-Ralbp1-RalA module. Sci. Adv. 2023, 8, eade2540. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker Primary Antibody—Species and Dilution | Company, City, and State of Purchase | Secondary Antibody, Dilution | Company, City, and State of Purchase |
|---|---|---|---|
| RalBP1/Rlip Rabbit (polyclonal) # 3630S 1:1000 | Cell Signaling Technology Danvers, MA, USA | Donkey anti-rabbit HRP 1:10,000 | GE Healthcare Amersham, Piscataway, NJ, USA |
| Phospho-Tau (Ser202, Thr205) (AT8) Mouse (monoclonal) 1:500 #MN1020 | Invitrogen (ThermoFisher Sci.) Waltham, MA, USA | Sheep anti-mouse HRP 1:10,000 | GE Healthcare Amersham, Piscataway, NJ, USA |
| SYN Rabbit monoclonal 1:400 #Ab32127 | Abcam, Cambridge, MA, USA | Donkey anti-rabbit HRP 1:10,000 | GE Healthcare Amersham, Piscataway, NJ, USA |
| PSD-95 Mouse Monoclonal Antibody 1:500 # MA1-046 | Invitrogen, Waltham, MA, USA | Sheep anti-mouse HRP 1:10,000 | GE Healthcare Amersham, Piscataway, NJ, USA |
| PINK1 Rabbit polyclonal 1:500 #BC100-494 | Novus Biological, Littleton, CO, USA | Donkey anti-rabbit HRP 1:10,000 | GE Healthcare Amersham, Piscataway, NJ, USA |
| Parkin Mouse polyclonal 1:500 #NBP2-29838 | Novus Biological, Littleton, CO, USA | Sheep anti-mouse HRP 1:10,000 | GE Healthcare Amersham, Piscataway, NJ, USA |
| Antibody | Species | Dilution Used | Supplier | Catalog Number |
|---|---|---|---|---|
| RalBP1 | Mouse (monoclonal) | 1:100 | Origene Technology | TA500964 |
| SYN | Rabbit monoclonal | 1:100 | Abcam, Cambridge, MA, USA | Ab32127 |
| PSD-95 | Mouse Monoclonal Antibody | 1:100 | Invitrogen, Waltham, MA, USA | MA1-046 |
| PINK1 | Rabbit polyclonal | 1:100 | Novus Biological, Littleton, CO, USA | BC100-494 |
| Parkin | Mouse polyclonal | 1:100 | Novus Biological, Littleton, CO, USA | NBP2-29838 |
| Phospho-Tau (Ser202, Thr205) (AT8) | Mouse (monoclonal) | 1:100 | Invitrogen (ThermoFisher Sci.) | MN1020 |
| Anti-mouse IgG AlexaFluor488 | Goat | 1:100 | Invitrogen (ThermoFisher Sci.) | A-11001 |
| Anti-rabbit (H+L), (Texas-red-conjugated) | Goat | 1:100 | Novus Biologicals | NB120-6719 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reddy, P.H.; Kshirsagar, S.; Bose, C.; Pradeepkiran, J.A.; Hindle, A.; Singh, S.P.; Reddy, A.P.; Baig, J. Rlip Reduction Induces Oxidative Stress and Mitochondrial Dysfunction in Mutant Tau-Expressed Immortalized Hippocampal Neurons: Mechanistic Insights. Cells 2023, 12, 1646. https://doi.org/10.3390/cells12121646
Reddy PH, Kshirsagar S, Bose C, Pradeepkiran JA, Hindle A, Singh SP, Reddy AP, Baig J. Rlip Reduction Induces Oxidative Stress and Mitochondrial Dysfunction in Mutant Tau-Expressed Immortalized Hippocampal Neurons: Mechanistic Insights. Cells. 2023; 12(12):1646. https://doi.org/10.3390/cells12121646
Chicago/Turabian StyleReddy, P. Hemachandra, Sudhir Kshirsagar, Chhanda Bose, Jangampalli Adi Pradeepkiran, Ashly Hindle, Sharda P. Singh, Arubala P. Reddy, and Javaria Baig. 2023. "Rlip Reduction Induces Oxidative Stress and Mitochondrial Dysfunction in Mutant Tau-Expressed Immortalized Hippocampal Neurons: Mechanistic Insights" Cells 12, no. 12: 1646. https://doi.org/10.3390/cells12121646