
SNARE Protein Snc1 Is Essential for Vesicle Trafficking, Membrane Fusion and Protein Secretion in Fungi

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Structure of Snc1 Protein
3. Regulatory Role of Snc1 in Endocytosis and Exocytosis
3.1. Exocytosis
3.2. Endocytosis
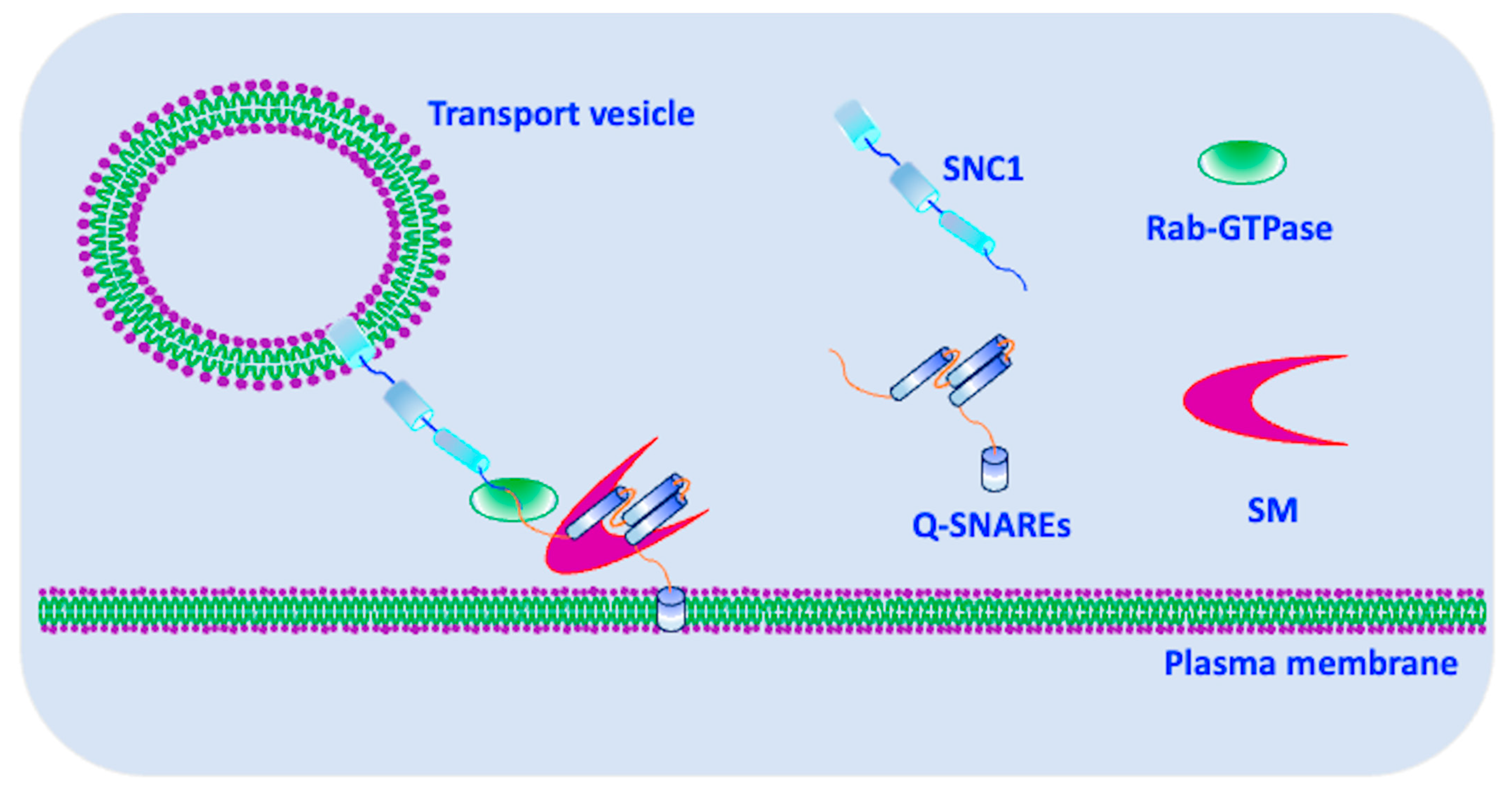
4. Assembly and Disassembly of Snc1 SNARE Complex
5. Impact of Ubiquitination on the Localization and Recycling of Snc1
6. Various Snc1 Interacting Proteins and Their Role in Cellular Trafficking in Fungi
7. Role of Snc1 in Protein Secretion and Development of Fungi
8. Strategies for Enhancing Fungal Protein Production through SNC1 Genetic Manipulation
- Overexpression of SNC1: As mentioned earlier, the overexpression of SNC1 was shown to increase fungal protein secretion. Thus, one potential strategy by which to enhance protein production would be to overexpress SNC1 in the fungal hosts. This could be achieved through various methods, such as promoter engineering, gene modification, or increasing the gene copy number. The overexpression of SNC1 can be achieved using strong promoters such as Ppdc1, Pcdna1 or Pcbh1. Genetically engineered promoters, which include the transcriptional factors of both the constitutive and inducible promoters, remain active even under different media. For more details, please visit the following article [78].
- The co-expression of SNC1 along with key regulatory genes: SNC1 is just one of many genes that play a significant role in the protein secretion of filamentous fungi. Thus, another strategy by which to enhance protein production would be to co-express SNC1 with other genes involved in protein secretion, such as chaperones, transporters or related SNAREs (please consult [18] for reference). This could potentially enhance the efficiency and fidelity of the protein secretion pathway.
- Targeted deletion of SNC1 inhibitors: Several proteins have been identified that can inhibit the function of SNC1 and other vesicular trafficking proteins. The activation of the glycogen synthase kinase (GSK-3β) retards the exocytosis of the synaptic vesicle in response to membrane depolarization [79]. Generally, the HOPS complex works synergistically with Sec17/18 during SANRE assembly and disassembly, respectively. Strikingly, HOPS inhibits the disassembly of SNARE complexes in the trans-, but not in the cis-, configuration [80]. Similarly, elevated levels of Sec17 can inhibit vacuole fusion through the recapture of primed SNAREs [81]. MED (myristoylated alanine-rich C kinase substrate effector domain) may generally inhibit vacuolar lipid rearrangements or may interfere with the essential interactions of SNAREs with lipids [81]. Pobbati et al. suggested that tomosyn can act as a negative regulator of exocytosis by inhibiting the binding of the vesicular synaptobrevin 2 to its plasma membrane acceptors [82]. Thus, a strategy by which to enhance protein production would be to genetically manipulate these inhibitors to increase the activity of Snc1.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pimentel, P.S.S.-R.; de Oliveira, J.B.; Astolfi-Filho, S.; Pereira, N., Jr. Enzymatic Hydrolysis of Lignocellulosic Biomass Using an Optimized Enzymatic Cocktail Prepared from Secretomes of Filamentous Fungi Isolated from Amazonian Biodiversity. Appl. Biochem. Biotechnol. 2021, 193, 3915–3935. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zheng, X.; Li, G.; Wang, X. TORC1 Signaling in Fungi: From Yeasts to Filamentous Fungi. Microorganisms 2023, 11, 218. [Google Scholar] [CrossRef]
- Staples, M.I.; Frazer, C.; Fawzi, N.L.; Bennett, R.J. Phase Separation in Fungi. Nat. Microbiol. 2023, 8, 375–386. [Google Scholar] [CrossRef]
- Fitz, E.; Wanka, F.; Seiboth, B. The Promoter Toolbox for Recombinant Gene Expression in Trichoderma Reesei. Front. Bioeng. Biotechnol. 2018, 6, 135. [Google Scholar] [CrossRef]
- Choi, U.B.; Dunleavy, K.; Matlock, H.; Gething, C.; Howells, G.; Misra, B.; White, K.I.; Brunger, A. Conformational Dynamics of SNARE Recycling Mediated by NSF. FASEB J. 2022, 36. [Google Scholar] [CrossRef]
- Adnan, M.; Islam, W.; Zhang, J.; Zheng, W.; Lu, G.-D. Diverse Role of SNARE Protein Sec22 in Vesicle Trafficking, Membrane Fusion, and Autophagy. Cells 2019, 8, 337. [Google Scholar] [CrossRef] [Green Version]
- Adnan, M.; Fang, W.; Sun, P.; Zheng, Y.; Abubakar, Y.S.; Zhang, J.; Lou, Y.; Zheng, W.; Lu, G. R-SNARE FgSec22 Is Essential for Growth, Pathogenicity and DON Production of Fusarium Graminearum. Curr. Genet. 2020, 66, 421–435. [Google Scholar] [CrossRef]
- Adnan, M.; Islam, W.; Noman, A.; Hussain, A.; Anwar, M.; Khan, M.U.; Akram, W.; Ashraf, M.F.; Raza, M.F. Q-SNARE Protein FgSyn8 Plays Important Role in Growth, DON Production and Pathogenicity of Fusarium Graminearum. Microb. Pathog. 2020, 140, 103948. [Google Scholar] [CrossRef]
- Valkonen, M.; Kalkman, E.R.; Saloheimo, M.; Penttilaö, M.; Read, N.D.; Duncan, R.R. Spatially Segregated SNARE Protein Interactions in Living Fungal Cells. J. Biol. Chem. 2007, 282, 22775–22785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, B.; Guo, W. The Exocyst Complex in Polarized Exocytosis. Curr. Opin. Cell Biol. 2009, 21, 537–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aalto, M.K.; Ronne, H.; Keränen, S. Yeast Syntaxins Sso1p and Sso2p Belong to a Family of Related Membrane Proteins That Function in Vesicular Transport. EMBO J. 1993, 12, 4095–4104. [Google Scholar] [CrossRef]
- Hong, W.; Lev, S. Tethering the Assembly of SNARE Complexes. Trends Cell Biol. 2014, 24, 35–43. [Google Scholar] [CrossRef]
- Rizo, J.; Sari, L.; Qi, Y.; Im, W.; Lin, M.M. All-Atom Molecular Dynamics Simulations of Synaptotagmin-SNARE-Complexin Complexes Bridging a Vesicle and a Flat Lipid Bilayer. Elife 2022, 11, e76356. [Google Scholar] [CrossRef]
- Wang, S.; Ma, C. Neuronal SNARE Complex Assembly Guided by Munc18-1 and Munc13-1. FEBS Open Bio 2022, 12, 1939–1957. [Google Scholar] [CrossRef]
- Fischer, R.; Zekert, N.; Takeshita, N. Polarized Growth in Fungi–Interplay between the Cytoskeleton, Positional Markers and Membrane Domains. Mol. Microbiol. 2008, 68, 813–826. [Google Scholar] [CrossRef]
- Best, J.T.; Xu, P.; McGuire, J.G.; Leahy, S.N.; Graham, T.R. Yeast Synaptobrevin, Snc1, Engages Distinct Routes of Postendocytic Recycling Mediated by a Sorting Nexin, Rcy1-COPI, and Retromer. Mol. Biol. Cell 2020, 31, 944–962. [Google Scholar] [CrossRef]
- Fiedler, M.R.; Barthel, L.; Kubisch, C.; Nai, C.; Meyer, V. Construction of an Improved Aspergillus Niger Platform for Enhanced Glucoamylase Secretion. Microb. Cell Factories 2018, 17, 1–12. [Google Scholar] [CrossRef]
- Wu, Y.; Sun, X.; Xue, X.; Luo, H.; Yao, B.; Xie, X.; Su, X. Overexpressing Key Component Genes of the Secretion Pathway for Enhanced Secretion of an Aspergillus Niger Glucose Oxidase in Trichoderma Reesei. Enzym. Microb. Technol. 2017, 106, 83–87. [Google Scholar] [CrossRef]
- Yang, S.; Zhou, X.; Guo, P.; Lin, Y.; Fan, Q.; Zuriegat, Q.; Lu, S.; Yang, J.; Yu, W.; Liu, H. The Exocyst Regulates Hydrolytic Enzyme Secretion at Hyphal Tips and Septa in the Banana Fusarium Wilt Fungus Fusarium Odoratissimum. Appl. Environ. Microbiol. 2021, 87, e03088-20. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Starr, T.L.; Glass, N.L. Plant Cell Wall–Degrading Enzymes and Their Secretion in Plant-Pathogenic Fungi. Annu. Rev. Phytopathol. 2014, 52, 427–451. [Google Scholar] [CrossRef]
- Rizo, J.; Südhof, T.C. The Membrane Fusion Enigma: SNAREs, Sec1/Munc18 Proteins, and Their Accomplices—Guilty as Charged? Annu. Rev. Cell Dev. Biol. 2012, 28, 279–308. [Google Scholar] [CrossRef] [PubMed]
- Grissom, J.H.; Segarra, V.A.; Chi, R.J. New Perspectives on Snare Function in the Yeast Minimal Endomembrane System. Genes 2020, 11, 899. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.; Lepore, D.; Kenner, L.; Czuchra, A.B.; Plooster, M.; Frost, A.; Munson, M.; Brennwald, P. Exocyst Structural Changes Associated with Activation of Tethering Downstream of Rho/Cdc42 GTPases. J. Cell Biol. 2020, 219. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; McFarlane, H.E. Regulation of Cellulose Synthesis via Exocytosis and Endocytosis. Curr. Opin. Plant Biol. 2022, 69, 102273. [Google Scholar] [CrossRef]
- Fasshauer, D.; Jahn, R. Budding Insights on Cell Polarity. Nat. Struct. Mol. Biol. 2007, 14, 360–362. [Google Scholar] [CrossRef] [Green Version]
- Ma, M.; Burd, C.G. Retrograde Trafficking and Quality Control of Yeast Synaptobrevin, Snc1, Are Conferred by Its Transmembrane Domain. Mol. Biol. Cell 2019, 30, 1729–1742. [Google Scholar] [CrossRef]
- Fiedler, M.R.; Cairns, T.C.; Koch, O.; Kubisch, C.; Meyer, V. Conditional Expression of the Small GTPase ArfA Impacts Secretion, Morphology, Growth, and Actin Ring Position in Aspergillus Niger. Front. Microbiol. 2018, 9, 878. [Google Scholar] [CrossRef]
- Burston, H.E.; Maldonado-Báez, L.; Davey, M.; Montpetit, B.; Schluter, C.; Wendland, B.; Conibear, E. Regulators of Yeast Endocytosis Identified by Systematic Quantitative Analysis. J. Cell Biol. 2009, 185, 1097–1110. [Google Scholar] [CrossRef] [Green Version]
- Kashikuma, R.; Nagano, M.; Shimamura, H.; Nukaga, K.; Katsumata, I.; Toshima, J.Y.; Toshima, J. Role of Phosphatidylserine in the Localization of Cell Surface Membrane Proteins in Yeast. Cell Struct. Funct. 2023, 48, 19–30. [Google Scholar] [CrossRef]
- Oberhofer, E. Ubiquitin Modifikation Der Synaptobervine SNC1 Und SNC2 in Saccharomyces Cerevisiae. Bachelor’s Thesis, LMU Munich, Munich, Germany, 2004. [Google Scholar]
- Burri, L.; Lithgow, T. A Complete Set of SNAREs in Yeast. Traffic 2004, 5, 45–52. [Google Scholar] [CrossRef]
- Lewis, M.J.; Pelham, H.R. A New Yeast Endosomal SNARE Related to Mammalian Syntaxin 8. Traffic 2002, 3, 922–929. [Google Scholar] [CrossRef]
- Shanks, S.G.; Carpp, L.N.; Struthers, M.S.; McCann, R.K.; Bryant, N.J. The Sec1/Munc18 Protein Vps45 Regulates Cellular Levels of Its SNARE Binding Partners Tlg2 and Snc2 in Saccharomyces Cerevisiae. PLoS ONE 2012, 7, e49628. [Google Scholar] [CrossRef] [Green Version]
- Apel, A.R.; Hoban, K.; Chuartzman, S.; Tonikian, R.; Sidhu, S.; Schuldiner, M.; Wendland, B.; Prosser, D. Syp1 Regulates the Clathrin-Mediated and Clathrin-Independent Endocytosis of Multiple Cargo Proteins through a Novel Sorting Motif. Mol. Biol. Cell 2017, 28, 2434–2448. [Google Scholar] [CrossRef]
- Wrasman, K.M. Endocytic Regulation from Cargo to Coat: The Identification of Novel Factors and Endocytic Protein Interactions; Johns Hopkins University: Baltimore, MD, USA, 2016. [Google Scholar]
- Grote, E.; Vlacich, G.; Pypaert, M.; Novick, P.J. A Snc1 Endocytosis Mutant: Phenotypic Analysis and Suppression by Overproduction of Dihydrosphingosine Phosphate Lyase. Mol. Biol. Cell 2000, 11, 4051–4065. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Gonzalez, L.; Prekeris, R.; Steegmaier, M.; Advani, R.J.; Scheller, R.H. SNARE Interactions Are Not Selective: Implications for Membrane Fusion Specificity. J. Biol. Chem. 1999, 274, 5649–5653. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, N.; Mima, J. Multiple and Distinct Strategies of Yeast SNAREs to Confer the Specificity of Membrane Fusion. Sci. Rep. 2014, 4, 4277. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Zhang, M.; Li, J.; Yang, C.; Abubakar, Y.S.; Chen, X.; Zheng, W.; Wang, Z.; Zheng, H.; Zhou, J. The Small GTPase FgRab1 Plays Indispensable Roles in the Vegetative Growth, Vesicle Fusion, Autophagy and Pathogenicity of Fusarium Graminearum. Int. J. Mol. Sci. 2022, 23, 895. [Google Scholar] [CrossRef]
- Chen, X.; Selvaraj, P.; Lin, L.; Fang, W.; Wu, C.; Yang, P.; Zhang, J.; Abubakar, Y.S.; Yang, F.; Lu, G. Rab7/Retromer-Based Endolysosomal Trafficking Facilitates Effector Secretion and Host Invasion in Rice Blast. bioRxiv 2022. [Google Scholar] [CrossRef]
- Gerst, J.E.; Rodgers, L.; Riggs, M.; Wigler, M. SNC1, a Yeast Homolog of the Synaptic Vesicle-Associated Membrane Protein/Synaptobrevin Gene Family: Genetic Interactions with the RAS and CAP Genes. Proc. Natl. Acad. Sci. USA 1992, 89, 4338–4342. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Lin, Y.; Fang, W.; Zhao, X.; Lou, Y.; Wang, G.; Zheng, H.; Liang, Q.; Abubakar, Y.S.; Olsson, S. The Endosomal Recycling of FgSnc1 by FgSnx41–FgSnx4 Heterodimer Is Essential for Polarized Growth and Pathogenicity in Fusarium Graminearum. New Phytol. 2018, 219, 654–671. [Google Scholar] [CrossRef] [Green Version]
- Valkonen, M. Functional Studies of the Secretory Pathway of Filamentous Fungi: The Effect of Unfolded Protein Response on Protein Production; Edita Prima Oy: Helsinki, Finland, 2003. [Google Scholar]
- Protopopov, V.; Govindan, B.; Novick, P.; Gerst, J.E. Homologs of the Synaptobrevin/VAMP Family of Synaptic Vesicle Proteins Function on the Late Secretory Pathway in S. Cerevisiae. Cell 1993, 74, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.; Yuan, H.; Hutagalung, A.; Verma, A.; Kümmel, D.; Wu, X.; Reinisch, K.; McNew, J.A.; Novick, P. The Synaptobrevin Homologue Snc2p Recruits the Exocyst to Secretory Vesicles by Binding to Sec6p. J. Cell Biol. 2013, 202, 509–526. [Google Scholar] [CrossRef] [PubMed]
- Gerst, J.E. Conserved α-Helical Segments on Yeast Homologs of the Synaptobrevin/VAMP Family of v-SNAREs Mediate Exocytic Function. J. Biol. Chem. 1997, 272, 16591–16598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGuffin, L.J.; Bryson, K.; Jones, D.T. The PSIPRED Protein Structure Prediction Server. Bioinformatics 2000, 16, 404–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Chen, H.; Yuan, M.; Zhang, M.; Abubakar, Y.S.; Chen, X.; Zhong, H.; Zheng, W.; Zheng, H.; Zhou, J. FgAP1σ Is Critical for Vegetative Growth, Conidiation, Virulence, and DON Biosynthesis in Fusarium Graminearum. J. Fungi 2023, 9, 145. [Google Scholar] [CrossRef] [PubMed]
- Buwa, N.; Martínez-Núñez, L.; Munson, M. Exocytosis in Yeast: Major Players and Mechanisms. In Exocytosis: From Molecules to Cells; IOP Publishing: Bristol, England, 2022. [Google Scholar]
- Jahn, R.; Scheller, R.H. SNAREs—Engines for Membrane Fusion. Nat. Rev. Mol. Cell Biol. 2006, 7, 631–643. [Google Scholar] [CrossRef]
- Lee, C.; Lepore, D.; Munson, M.; Yoon, T.-Y. Exocyst Stimulates Each Step of Exocytic SNARE Complex Assembly and Vesicle Fusion. bioRxiv 2022. [Google Scholar] [CrossRef]
- Thattai, M. Molecular and Cellular Constraints on Vesicle Traffic Evolution. Curr. Opin. Cell Biol. 2023, 80, 102151. [Google Scholar] [CrossRef]
- Starr, T.L.; Gonçalves, A.P.; Meshgin, N.; Glass, N.L. The Major Cellulases CBH-1 and CBH-2 of Neurospora Crassa Rely on Distinct ER Cargo Adaptors for Efficient ER-exit. Mol. Microbiol. 2018, 107, 229–248. [Google Scholar] [CrossRef] [Green Version]
- Valdez-Taubas, J.; Pelham, H.R. Slow Diffusion of Proteins in the Yeast Plasma Membrane Allows Polarity to Be Maintained by Endocytic Cycling. Curr. Biol. 2003, 13, 1636–1640. [Google Scholar] [CrossRef] [Green Version]
- Bonifacino, J.S.; Rojas, R. Retrograde Transport from Endosomes to the Trans-Golgi Network. Nat. Rev. Mol. Cell Biol. 2006, 7, 568–579. [Google Scholar] [CrossRef]
- Gurunathan, S.; Marash, M.; Weinberger, A.; Gerst, J.E. T-SNARE Phosphorylation Regulates Endocytosis in Yeast. Mol. Biol. Cell 2002, 13, 1594–1607. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Hankins, H.M.; MacDonald, C.; Erlinger, S.J.; Frazier, M.N.; Diab, N.S.; Piper, R.C.; Jackson, L.P.; MacGurn, J.A.; Graham, T.R. COPI Mediates Recycling of an Exocytic SNARE by Recognition of a Ubiquitin Sorting Signal. Elife 2017, 6, e28342. [Google Scholar] [CrossRef]
- Zhang, F.; Zhao, M.; Braun, D.R.; Ericksen, S.S.; Piotrowski, J.S.; Nelson, J.; Peng, J.; Ananiev, G.E.; Chanana, S.; Barns, K. A Marine Microbiome Antifungal Targets Urgent-Threat Drug-Resistant Fungi. Science 2020, 370, 974–978. [Google Scholar] [CrossRef]
- Abenza, J.F.; Pantazopoulou, A.; Rodríguez, J.M.; Galindo, A.; Penalva, M.A. Long-distance Movement of Aspergillus Nidulans Early Endosomes on Microtubule Tracks. Traffic 2009, 10, 57–75. [Google Scholar] [CrossRef]
- Hervás-Aguilar, A.; Peñalva, M.A. Endocytic Machinery Protein SlaB Is Dispensable for Polarity Establishment but Necessary for Polarity Maintenance in Hyphal Tip Cells of Aspergillus Nidulans. Eukaryot. Cell 2010, 9, 1504–1518. [Google Scholar] [CrossRef] [Green Version]
- Taheri-Talesh, N.; Horio, T.; Araujo-Bazán, L.; Dou, X.; Espeso, E.A.; Penalva, M.A.; Osmani, S.A.; Oakley, B.R. The Tip Growth Apparatus of Aspergillus Nidulans. Mol. Biol. Cell 2008, 19, 1439–1449. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, Y.; Shoji, J.; Arioka, M.; Kitamoto, K. Endocytosis Is Crucial for Cell Polarity and Apical Membrane Recycling in the Filamentous Fungus Aspergillus Oryzae. Eukaryot. Cell 2009, 8, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Orr, A.; Duan, M.; Merz, A.J.; Wickner, W. Sec17/Sec18 Act Twice, Enhancing Membrane Fusion and Then Disassembling Cis-SNARE Complexes. Elife 2017, 6, e26646. [Google Scholar] [CrossRef]
- Best, J.T.; Xu, P.; Graham, T.R. Phospholipid Flippases in Membrane Remodeling and Transport Carrier Biogenesis. Curr. Opin. Cell Biol. 2019, 59, 8–15. [Google Scholar] [CrossRef]
- Yoon, T.-Y.; Munson, M. SNARE Complex Assembly and Disassembly. Curr. Biol. 2018, 28, R397–R401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparks, R.P.; Arango, A.S.; Starr, M.L.; Aboff, Z.L.; Hurst, L.R.; Rivera-Kohr, D.A.; Zhang, C.; Harnden, K.A.; Jenkins, J.L.; Guida, W.C. A Small-Molecule Competitive Inhibitor of Phosphatidic Acid Binding by the AAA+ Protein NSF/Sec18 Blocks the SNARE-Priming Stage of Vacuole Fusion. J. Biol. Chem. 2019, 294, 17168–17185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Wu, S.; Zhou, Q.; Vivona, S.; Cipriano, D.J.; Cheng, Y.; Brunger, A.T. Mechanistic Insights into the Recycling Machine of the SNARE Complex. Nature 2015, 518, 61–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, M.J.; Nichols, B.J.; Prescianotto-Baschong, C.; Riezman, H.; Pelham, H.R. Specific Retrieval of the Exocytic SNARE Snc1p from Early Yeast Endosomes. Mol. Biol. Cell 2000, 11, 23–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.H.; Shah, A.H.; Segev, N. Ypt31/32 GTPases and Their F-Box Effector Rcy1 Regulate Ubiquitination of Recycling Proteins. Cell. Logist. 2011, 1, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Schneider, K.; Farr, T.; Pinter, N.; Schmitt, K.; Valerius, O.; Braus, G.H.; Kämper, J. The Nma1 Protein Promotes Long Distance Transport Mediated by Early Endosomes in Ustilago Maydis. Mol. Microbiol. 2022, 117, 334–352. [Google Scholar] [CrossRef]
- Kuratsu, M.; Taura, A.; Shoji, J.Y.; Kikuchi, S.; Arioka, M.; Kitamoto, K. Systematic analysis of SNARE localization in the filamentous fungus Aspergillus oryzae. Fungal Genet. Biol. 2007, 44, 1310–1323. [Google Scholar] [CrossRef]
- Ji, W.; Wang, X.; Liu, X.; Wang, Y.; Liu, F.; Xu, B.; Luo, H.; Tu, T.; Zhang, W.; Xu, X. Combining Manipulation of Integration Loci and Secretory Pathway on Expression of an Aspergillus Niger Glucose Oxidase Gene in Trichoderma Reesei. Microb. Cell Factories 2023, 22, 1–11. [Google Scholar] [CrossRef]
- Van Zyl, J.H.D.; Den Haan, R.; Van Zyl, W.H. Over-Expression of Native Saccharomyces Cerevisiae Exocytic SNARE Genes Increased Heterologous Cellulase Secretion. Appl. Microbiol. Biotechnol. 2014, 98, 5567–5578. [Google Scholar] [CrossRef]
- Couve, A.; Gerst, J.E. Yeast Snc Proteins Complex with Sec9. Functional Interactions between Putative SNARE Proteins. J. Biol. Chem. 1994, 269, 23391–23394. [Google Scholar] [CrossRef]
- Pakula, T.M.; Laxell, M.; Huuskonen, A.; Uusitalo, J.; Saloheimo, M.; Penttilä, M. The Effects of Drugs Inhibiting Protein Secretion in the Filamentous Fungus Trichoderma Reesei: Evidence for down-Regulation of Genes That Encode Secreted Proteins in the Stressed Cells. J. Biol. Chem. 2003, 278, 45011–45020. [Google Scholar] [CrossRef] [Green Version]
- Gasser, B.; Saloheimo, M.; Rinas, U.; Dragosits, M.; Rodríguez-Carmona, E.; Baumann, K.; Giuliani, M.; Parrilli, E.; Branduardi, P.; Lang, C. Protein Folding and Conformational Stress in Microbial Cells Producing Recombinant Proteins: A Host Comparative Overview. Microb. Cell Factories 2008, 7, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Lipatova, Z.; Segev, N. A Role for Macro-ER-Phagy in ER Quality Control. PLoS Genet. 2015, 11, e1005390. [Google Scholar] [CrossRef]
- Adnan, M.; Ma, X.; Olsson, S.; Wang, J.; Liu, G. Promoter Regulation and Genetic Engineering Strategies for Enhanced Cellulase Expression in Trichoderma Reesei. Microbiol. Res. 2022, 259, 127011. [Google Scholar] [CrossRef]
- Zhu, L.-Q.; Liu, D.; Hu, J.; Cheng, J.; Wang, S.-H.; Wang, Q.; Wang, F.; Chen, J.-G.; Wang, J.-Z. GSK-3β Inhibits Presynaptic Vesicle Exocytosis by Phosphorylating P/Q-Type Calcium Channel and Interrupting Snare Complex Formation. J. Neurosci. 2010, 30, 3624–3633. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Jun, Y.; Thompson, J.; Yates, J.; Wickner, W. HOPS Prevents the Disassembly of Trans-SNARE Complexes by Sec17p/Sec18p during Membrane Fusion. EMBO J. 2010, 29, 1948–1960. [Google Scholar] [CrossRef] [Green Version]
- Collins, K.M.; Wickner, W.T. Trans-SNARE Complex Assembly and Yeast Vacuole Membrane Fusion. Proc. Natl. Acad. Sci. USA 2007, 104, 8755–8760. [Google Scholar] [CrossRef] [Green Version]
- Pobbati, A.V.; Razeto, A.; Böddener, M.; Becker, S.; Fasshauer, D. Structural Basis for the Inhibitory Role of Tomosyn in Exocytosis. J. Biol. Chem. 2004, 279, 47192–47200. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Function | Interaction with SNC1 | References |
|---|---|---|---|
| Sso1/Sso2 | Vesicle fusion with plasma membrane | Essential for exocytosis of secretory vesicles | [20] |
| Sec1/Munc18 | Docking of secretory vesicles and their fusion with the PM | Critical for efficient vesicle fusion with plasma membrane | [12] |
| Sec9 | Docking of cargo vesicles and their fusion | Fusion of the secretory vesicles and PM for efficient exocytosis | [20,21] |
| Sec18/NSF Sec17/SNAP | Assembly and disassembly of SNAREs | Assembly and disassembly of SNC1 mediated SNAREs | [22] |
| Exo70/ Exo84 | Important components of exocyst complex | Promotes SNC1 localization towards the exocytic sites on the PM | [23,24] |
| Sec3 | Important components of the exocyst complex | Promote exocytic complex formation at the PM | [23,24] |
| Sec5 | |||
| Sec6 | |||
| Sec8 | |||
| Sec15 | |||
| Sro7/Sro77 | Exocytosis and actin organization | Regulate trafficking and localization of SNC1 to specific membrane domains | [25] |
| Cdc42 | Subunit of exocyst complex | Fusion of secretory vesicles and the PM | [10] |
| Drs2-Cdc50 | Phospholipid flippase (Subunit of exocyst complex) | Post-endocytic recycling of SNC1 | [16,26] |
| Rcy1 | F-box protein (Subunit of exocyst complex) | ||
| Snx4-Atg20 | sorting nexin (Subunit of exocyst complex) | ||
| COPI coat complex | COPI coat complex surrounds the cargo vesicles for cellular transportation | ||
| ArfA | Recruitment of COPI to Golgi membranes | Regulate SNC1 localization and activity in the secretory pathway | [27] |
| AP180 | Functional role in endocytosis | Play a cargo-specific role in SNC1 internalization | [28] |
| Tlg1/Tlg2 | t-SNAREs localized to late Golgi and endosomes | Recycling of Snc1 protein from the PM towards Golgi | [22] |
| YPT1 | GTPase involved in vesicle trafficking | Facilitates the fusion of vesicles | [12] |
| Sla1 | Endocytosis and actin organization | SNC1 and Sla1 interaction regulates the internalization of the α-factor receptor Ste2 | [29] |
| End3/End4 | Endocytosis and actin organization | Efficient endocytosis of a subset of membrane proteins | [30] |
| Vam3/Vam7 | Vesicle fusion | Efficient fusion of vesicles with the vacuole | [22,31] |
| Pep12 | Endosomal and vacuolar trafficking | Essential for Snc1 recycling from endosomes to PM | [26,32] |
| Vps41 | Key regulator of SNC1/2 recycling | Interacts with SNC1/2 and promotes its sorting into recycling vesicles | [33] |
| Ede1 | Endocytic proteins | Mediate vesicle fusion during endocytosis | [34,35] |
| Syp1 | |||
| Pal1 | |||
| Vti1 | Trans-Golgi network to endosome transportation of vesicles | Fusion of secretory vesicles and the PM | [26,32] |
| Sec22 | ER-to-Golgi vesicular transport | Can complement SNC1 in vivo | [36,37] |
| Nyv1 | Fusion of vesicles with the vacuole | Efficient vesicle fusion with the vacuole | [22,32] |
| Syn8 | Endosomal t-SNARE involved in vesicle fusion | Exhibits promiscuous interactions with Snc1/2 | [32] |
| Sed5 | t-SNAREs of cis-Golgi | Mediate fusion of initial PM derived vesicles | [38] |
| RSP5 | Ubiquitin ligase | Efficient recycling of SNC1 | [30] |
| Rab1 | Rab-GTPase Regulate vesicular trafficking | Rab1 inactivation blocks SNC1 recycling from Golgi to PM | [39] |
| Rab7 | Rab-GTPase Regulate vesicular trafficking | Regulate SNC1 trafficking from trans-Golgi network to PM and vice versa | [40] |
| Ras2 | Ras protein required to activate adenylate cyclase pathway | Ras2-SNC1 interaction suggest a role in response to nutrient availability | [41] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adnan, M.; Islam, W.; Waheed, A.; Hussain, Q.; Shen, L.; Wang, J.; Liu, G. SNARE Protein Snc1 Is Essential for Vesicle Trafficking, Membrane Fusion and Protein Secretion in Fungi. Cells 2023, 12, 1547. https://doi.org/10.3390/cells12111547
Adnan M, Islam W, Waheed A, Hussain Q, Shen L, Wang J, Liu G. SNARE Protein Snc1 Is Essential for Vesicle Trafficking, Membrane Fusion and Protein Secretion in Fungi. Cells. 2023; 12(11):1547. https://doi.org/10.3390/cells12111547
Chicago/Turabian StyleAdnan, Muhammad, Waqar Islam, Abdul Waheed, Quaid Hussain, Ling Shen, Juan Wang, and Gang Liu. 2023. "SNARE Protein Snc1 Is Essential for Vesicle Trafficking, Membrane Fusion and Protein Secretion in Fungi" Cells 12, no. 11: 1547. https://doi.org/10.3390/cells12111547