

Modulation of NRF2/KEAP1 Signaling in Preeclampsia
 , , , and
, , , and
Abstract
:1. Introduction
2. NRF2/KEAP1 Signaling in PE
3. NRF2 Cellular Modulators
4. NRF2 Modulation by Natural Compounds
5. NRF2 Modulation by Synthetic Compounds
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Costa, M.A. The endocrine function of human placenta: An overview. Reprod. Biomed. Online 2016, 32, 14–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tossetta, G.; Avellini, C.; Licini, C.; Giannubilo, S.R.; Castellucci, M.; Marzioni, D. High temperature requirement A1 and fibronectin: Two possible players in placental tissue remodelling. Eur. J. Histochem. 2016, 60, 2724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, K.J.; Seow, K.M.; Chen, K.H. Preeclampsia: Recent Advances in Predicting, Preventing, and Managing the Maternal and Fetal Life-Threatening Condition. Int. J. Environ. Res. Public. Health 2023, 20, 2994. [Google Scholar] [CrossRef] [PubMed]
- Tossetta, G.; Fantone, S.; Delli Muti, N.; Balercia, G.; Ciavattini, A.; Giannubilo, S.R.; Marzioni, D. Preeclampsia and severe acute respiratory syndrome coronavirus 2 infection: A systematic review. J. Hypertens. 2022, 40, 1629–1638. [Google Scholar] [CrossRef]
- Gesuita, R.; Licini, C.; Picchiassi, E.; Tarquini, F.; Coata, G.; Fantone, S.; Tossetta, G.; Ciavattini, A.; Castellucci, M.; Di Renzo, G.C.; et al. Association between first trimester plasma htra1 level and subsequent preeclampsia: A possible early marker? Pregnancy Hypertens. 2019, 18, 58–62. [Google Scholar] [CrossRef]
- Burton, G.J.; Redman, C.W.; Roberts, J.M.; Moffett, A. Pre-eclampsia: Pathophysiology and clinical implications. BMJ 2019, 366, l2381. [Google Scholar] [CrossRef] [Green Version]
- Huppertz, B. The Critical Role of Abnormal Trophoblast Development in the Etiology of Preeclampsia. Curr. Pharm. Biotechnol. 2018, 19, 771–780. [Google Scholar] [CrossRef]
- Fantone, S.; Mazzucchelli, R.; Giannubilo, S.R.; Ciavattini, A.; Marzioni, D.; Tossetta, G. AT-rich interactive domain 1A protein expression in normal and pathological pregnancies complicated by preeclampsia. Histochem. Cell Biol. 2020, 154, 339–346. [Google Scholar] [CrossRef]
- Deshpande, J.S.; Sundrani, D.P.; Sahay, A.S.; Gupte, S.A.; Joshi, S.R. Unravelling the potential of angiogenic factors for the early prediction of preeclampsia. Hypertens. Res. 2021, 44, 756–769. [Google Scholar] [CrossRef]
- Tenorio, M.B.; Ferreira, R.C.; Moura, F.A.; Bueno, N.B.; de Oliveira, A.C.M.; Goulart, M.O.F. Cross-Talk between Oxidative Stress and Inflammation in Preeclampsia. Oxidative Med. Cell. Longev. 2019, 2019, 8238727. [Google Scholar] [CrossRef] [Green Version]
- Cecati, M.; Sartini, D.; Campagna, R.; Biagini, A.; Ciavattini, A.; Emanuelli, M.; Giannubilo, S.R. Molecular analysis of endometrial inflammation in preterm birth. Cell. Mol. Biol. 2017, 63, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Licini, C.; Tossetta, G.; Avellini, C.; Ciarmela, P.; Lorenzi, T.; Toti, P.; Gesuita, R.; Voltolini, C.; Petraglia, F.; Castellucci, M.; et al. Analysis of cell-cell junctions in human amnion and chorionic plate affected by chorioamnionitis. Histol. Histopathol. 2016, 31, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Tossetta, G.; Fantone, S.; Gesuita, R.; Di Renzo, G.C.; Meyyazhagan, A.; Tersigni, C.; Scambia, G.; Di Simone, N.; Marzioni, D. HtrA1 in Gestational Diabetes Mellitus: A Possible Biomarker? Diagnostics 2022, 12, 2705. [Google Scholar] [CrossRef]
- delli Muti, N.; Salvio, G.; Ciarloni, A.; Perrone, M.; Tossetta, G.; Lazzarini, R.; Bracci, M.; Balercia, G. Can Extremely Low Frequency Magnetic Field Affect Human Sperm Parameters and Male Fertility? Tissue Cell 2023, 82, 102045. [Google Scholar] [CrossRef] [PubMed]
- Didziokaite, G.; Biliute, G.; Gudaite, J.; Kvedariene, V. Oxidative Stress as a Potential Underlying Cause of Minimal and Mild Endometriosis-Related Infertility. Int. J. Mol. Sci. 2023, 24, 3809. [Google Scholar] [CrossRef] [PubMed]
- Shu, D.Y.; Chaudhary, S.; Cho, K.S.; Lennikov, A.; Miller, W.P.; Thorn, D.C.; Yang, M.; McKay, T.B. Role of Oxidative Stress in Ocular Diseases: A Balancing Act. Metabolites 2023, 13, 187. [Google Scholar] [CrossRef] [PubMed]
- Tossetta, G.; Fantone, S.; Giannubilo, S.R.; Marzioni, D. The Multifaced Actions of Curcumin in Pregnancy Outcome. Antioxidants 2021, 10, 126. [Google Scholar] [CrossRef] [PubMed]
- Campagna, R.; Mateuszuk, L.; Wojnar-Lason, K.; Kaczara, P.; Tworzydlo, A.; Kij, A.; Bujok, R.; Mlynarski, J.; Wang, Y.; Sartini, D.; et al. Nicotinamide N-methyltransferase in endothelium protects against oxidant stress-induced endothelial injury. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 119082. [Google Scholar] [CrossRef]
- Zapotoczny, B.; Braet, F.; Kus, E.; Ginda-Makela, K.; Klejevskaja, B.; Campagna, R.; Chlopicki, S.; Szymonski, M. Actin-spectrin scaffold supports open fenestrae in liver sinusoidal endothelial cells. Traffic 2019, 20, 932–942. [Google Scholar] [CrossRef]
- Iantomasi, T.; Romagnoli, C.; Palmini, G.; Donati, S.; Falsetti, I.; Miglietta, F.; Aurilia, C.; Marini, F.; Giusti, F.; Brandi, M.L. Oxidative Stress and Inflammation in Osteoporosis: Molecular Mechanisms Involved and the Relationship with microRNAs. Int. J. Mol. Sci. 2023, 24, 3772. [Google Scholar] [CrossRef]
- Moratilla-Rivera, I.; Sanchez, M.; Valdes-Gonzalez, J.A.; Gomez-Serranillos, M.P. Natural Products as Modulators of Nrf2 Signaling Pathway in Neuroprotection. Int. J. Mol. Sci. 2023, 24, 3748. [Google Scholar] [CrossRef] [PubMed]
- Tossetta, G.; Marzioni, D. Natural and synthetic compounds in Ovarian Cancer: A focus on NRF2/KEAP1 pathway. Pharmacol. Res. 2022, 183, 106365. [Google Scholar] [CrossRef] [PubMed]
- Marzioni, D.; Mazzucchelli, R.; Fantone, S.; Tossetta, G. NRF2 modulation in TRAMP mice: An in vivo model of prostate cancer. Mol. Biol. Rep. 2023, 50, 873–881. [Google Scholar] [CrossRef]
- Tossetta, G.; Marzioni, D. Targeting the NRF2/KEAP1 pathway in cervical and endometrial cancers. Eur. J. Pharmacol. 2023, 941, 175503. [Google Scholar] [CrossRef] [PubMed]
- Tossetta, G.; Fantone, S.; Montanari, E.; Marzioni, D.; Goteri, G. Role of NRF2 in Ovarian Cancer. Antioxidants 2022, 11, 663. [Google Scholar] [CrossRef] [PubMed]
- Emanuelli, M.; Sartini, D.; Molinelli, E.; Campagna, R.; Pozzi, V.; Salvolini, E.; Simonetti, O.; Campanati, A.; Offidani, A. The Double-Edged Sword of Oxidative Stress in Skin Damage and Melanoma: From Physiopathology to Therapeutical Approaches. Antioxidants 2022, 11, 612. [Google Scholar] [CrossRef]
- Sartini, D.; Campagna, R.; Lucarini, G.; Pompei, V.; Salvolini, E.; Mattioli-Belmonte, M.; Molinelli, E.; Brisigotti, V.; Campanati, A.; Bacchetti, T.; et al. Differential immunohistochemical expression of paraoxonase-2 in actinic keratosis and squamous cell carcinoma. Hum. Cell 2021, 34, 1929–1931. [Google Scholar] [CrossRef]
- Torres-Cuevas, I.; Parra-Llorca, A.; Sanchez-Illana, A.; Nunez-Ramiro, A.; Kuligowski, J.; Chafer-Pericas, C.; Cernada, M.; Escobar, J.; Vento, M. Oxygen and oxidative stress in the perinatal period. Redox Biol. 2017, 12, 674–681. [Google Scholar] [CrossRef]
- Taysi, S.; Tascan, A.S.; Ugur, M.G.; Demir, M. Radicals, Oxidative/Nitrosative Stress and Preeclampsia. Mini Rev. Med. Chem. 2019, 19, 178–193. [Google Scholar] [CrossRef]
- Teimoori, B.; Moradi-Shahrebabak, M.; Razavi, M.; Rezaei, M.; Harati-Sadegh, M.; Salimi, S. The effect of GPx-1 rs1050450 and MnSOD rs4880 polymorphisms on PE susceptibility: A case-control study. Mol. Biol. Rep. 2019, 46, 6099–6104. [Google Scholar] [CrossRef]
- Luo, Z.C.; Julien, P.; Wei, S.Q.; Audibert, F.; Fraser, W.D.; Maternal and Infant Research on Oxidative Stress study group. Association of pre-eclampsia with SOD2 Ala16Val polymorphism among mother-father-infant triads. Int. J. Gynaecol. Obstet. 2018, 142, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Szczesny-Malysiak, E.; Stojak, M.; Campagna, R.; Grosicki, M.; Jamrozik, M.; Kaczara, P.; Chlopicki, S. Bardoxolone Methyl Displays Detrimental Effects on Endothelial Bioenergetics, Suppresses Endothelial ET-1 Release, and Increases Endothelial Permeability in Human Microvascular Endothelium. Oxidative Med. Cell. Longev. 2020, 2020, 4678252. [Google Scholar] [CrossRef] [PubMed]
- Baird, L.; Yamamoto, M. The Molecular Mechanisms Regulating the KEAP1-NRF2 Pathway. Mol. Cell. Biol. 2020, 40, e00099-20. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.M.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Shahcheraghi, S.H.; Salemi, F.; Small, S.; Syed, S.; Salari, F.; Alam, W.; Cheang, W.S.; Saso, L.; Khan, H. Resveratrol regulates inflammation and improves oxidative stress via Nrf2 signaling pathway: Therapeutic and biotechnological prospects. Phytother. Res. 2023, 37, 1590–1605. [Google Scholar] [CrossRef]
- Catanzaro, E.; Calcabrini, C.; Turrini, E.; Sestili, P.; Fimognari, C. Nrf2: A potential therapeutic target for naturally occurring anticancer drugs? Expert. Opin. Ther. Targets 2017, 21, 781–793. [Google Scholar] [CrossRef]
- Hamzawy, M.A.; Abo-Youssef, A.M.; Malak, M.N.; Khalaf, M.M. Multiple targets of Nrf 2 inhibitor; trigonelline in combating urethane-induced lung cancer by caspase-executioner apoptosis, cGMP and limitation of cyclin D1 and Bcl2. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 9393–9408. [Google Scholar] [CrossRef]
- Ni, C.; Ye, Q.; Mi, X.; Jiao, D.; Zhang, S.; Cheng, R.; Fang, Z.; Fang, M.; Ye, X. Resveratrol inhibits ferroptosis via activating NRF2/GPX4 pathway in mice with spinal cord injury. Microsc. Res. Tech. 2023. [Google Scholar] [CrossRef]
- Martin, K.R.; Barrett, J.C. Reactive oxygen species as double-edged swords in cellular processes: Low-dose cell signaling versus high-dose toxicity. Hum. Exp. Toxicol. 2002, 21, 71–75. [Google Scholar] [CrossRef]
- Sauer, H.; Wartenberg, M.; Hescheler, J. Reactive oxygen species as intracellular messengers during cell growth and differentiation. Cell Physiol. Biochem. 2001, 11, 173–186. [Google Scholar] [CrossRef]
- Fantone, S.; Tossetta, G.; Di Simone, N.; Tersigni, C.; Scambia, G.; Marcheggiani, F.; Giannubilo, S.R.; Marzioni, D. CD93 a potential player in cytotrophoblast and endothelial cell migration. Cell Tissue Res. 2022, 387, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Margioula-Siarkou, G.; Margioula-Siarkou, C.; Petousis, S.; Margaritis, K.; Vavoulidis, E.; Gullo, G.; Alexandratou, M.; Dinas, K.; Sotiriadis, A.; Mavromatidis, G. The role of endoglin and its soluble form in pathogenesis of preeclampsia. Mol. Cell. Biochem. 2022, 477, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Kweider, N.; Huppertz, B.; Kadyrov, M.; Rath, W.; Pufe, T.; Wruck, C.J. A possible protective role of Nrf2 in preeclampsia. Ann. Anat. 2014, 196, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.C.; Guilbert, L.J.; Yui, J.; Baker, P.N.; Davidge, S.T. The role of reactive nitrogen/oxygen intermediates in cytokine-induced trophoblast apoptosis. Placenta 1999, 20, 309–315. [Google Scholar] [CrossRef]
- Mam, V.; Tanbe, A.F.; Vitali, S.H.; Arons, E.; Christou, H.A.; Khalil, R.A. Impaired vasoconstriction and nitric oxide-mediated relaxation in pulmonary arteries of hypoxia- and monocrotaline-induced pulmonary hypertensive rats. J. Pharmacol. Exp. Ther. 2010, 332, 455–462. [Google Scholar] [CrossRef] [Green Version]
- Mansego, M.L.; Solar Gde, M.; Alonso, M.P.; Martinez, F.; Saez, G.T.; Escudero, J.C.; Redon, J.; Chaves, F.J. Polymorphisms of antioxidant enzymes, blood pressure and risk of hypertension. J. Hypertens. 2011, 29, 492–500. [Google Scholar] [CrossRef]
- George, E.M.; Granger, J.P. Heme oxygenase in pregnancy and preeclampsia. Curr. Opin. Nephrol. Hypertens. 2013, 22, 156–162. [Google Scholar] [CrossRef]
- Li, Y.; Yu, X.J.; Xiao, T.; Chi, H.L.; Zhu, G.Q.; Kang, Y.M. Nrf1 Knock-Down in the Hypothalamic Paraventricular Nucleus Alleviates Hypertension Through Intervention of Superoxide Production-Removal Balance and Mitochondrial Function. Cardiovasc. Toxicol. 2021, 21, 472–489. [Google Scholar] [CrossRef]
- Gomes, P.; Simao, S.; Lemos, V.; Amaral, J.S.; Soares-da-Silva, P. Loss of oxidative stress tolerance in hypertension is linked to reduced catalase activity and increased c-Jun NH2-terminal kinase activation. Free Radic. Biol. Med. 2013, 56, 112–122. [Google Scholar] [CrossRef]
- Li, J.; Zhang, C.; Xing, Y.; Janicki, J.S.; Yamamoto, M.; Wang, X.L.; Tang, D.Q.; Cui, T. Up-regulation of p27(kip1) contributes to Nrf2-mediated protection against angiotensin II-induced cardiac hypertrophy. Cardiovasc. Res. 2011, 90, 315–324. [Google Scholar] [CrossRef]
- Nezu, M.; Souma, T.; Yu, L.; Sekine, H.; Takahashi, N.; Wei, A.Z.; Ito, S.; Fukamizu, A.; Zsengeller, Z.K.; Nakamura, T.; et al. Nrf2 inactivation enhances placental angiogenesis in a preeclampsia mouse model and improves maternal and fetal outcomes. Sci. Signal. 2017, 10, eaam5711. [Google Scholar] [CrossRef] [PubMed]
- Giretti, I.; D’Ascenzo, R.; Correani, A.; Antognoli, L.; Monachesi, C.; Biagetti, C.; Pompilio, A.; Marinelli, L.; Burattini, I.; Cogo, P.; et al. Hypertriglyceridemia and lipid tolerance in preterm infants with a birth weight of less than 1250 g on routine parenteral nutrition. Clin. Nutr. 2021, 40, 4444–4448. [Google Scholar] [CrossRef] [PubMed]
- Nobile, S.; Marchionni, P.; Gidiucci, C.; Correani, A.; Palazzi, M.L.; Spagnoli, C.; Rondina, C.; Marche Neonatal, N.; Carnielli, V.P. Oxygen saturation/FIO2 ratio at 36 weeks’ PMA in 1005 preterm infants: Effect of gestational age and early respiratory disease patterns. Pediatr. Pulmonol. 2019, 54, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Mundal, S.B.; Rakner, J.J.; Silva, G.B.; Gierman, L.M.; Austdal, M.; Basnet, P.; Elschot, M.; Bakke, S.S.; Ostrop, J.; Thomsen, L.C.V.; et al. Divergent Regulation of Decidual Oxidative-Stress Response by NRF2 and KEAP1 in Preeclampsia with and without Fetal Growth Restriction. Int. J. Mol. Sci. 2022, 23, 1966. [Google Scholar] [CrossRef]
- Loset, M.; Mundal, S.B.; Johnson, M.P.; Fenstad, M.H.; Freed, K.A.; Lian, I.A.; Eide, I.P.; Bjorge, L.; Blangero, J.; Moses, E.K.; et al. A transcriptional profile of the decidua in preeclampsia. Am. J. Obstet. Gynecol. 2011, 204, 84.e1–84.e27. [Google Scholar] [CrossRef] [Green Version]
- Kweider, N.; Huppertz, B.; Wruck, C.J.; Beckmann, R.; Rath, W.; Pufe, T.; Kadyrov, M. A role for Nrf2 in redox signalling of the invasive extravillous trophoblast in severe early onset IUGR associated with preeclampsia. PLoS ONE 2012, 7, e47055. [Google Scholar] [CrossRef]
- Acar, N.; Soylu, H.; Edizer, I.; Ozbey, O.; Er, H.; Akkoyunlu, G.; Gemici, B.; Ustunel, I. Expression of nuclear factor erythroid 2-related factor 2 (Nrf2) and peroxiredoxin 6 (Prdx6) proteins in healthy and pathologic placentas of human and rat. Acta Histochem. 2014, 116, 1289–1300. [Google Scholar] [CrossRef]
- Wruck, C.J.; Huppertz, B.; Bose, P.; Brandenburg, L.O.; Pufe, T.; Kadyrov, M. Role of a fetal defence mechanism against oxidative stress in the aetiology of preeclampsia. Histopathology 2009, 55, 102–106. [Google Scholar] [CrossRef]
- Correani, A.; Visentin, S.; Cosmi, E.; Ponchia, E.; D’Aronco, S.; Simonato, M.; Vedovelli, L.; Cogo, P.; Carnielli, V.P. The maternal-fetal gradient of free and esterified phytosterols at the time of delivery in humans. Clin. Nutr. 2018, 37, 2107–2112. [Google Scholar] [CrossRef]
- Borst, P.; Evers, R.; Kool, M.; Wijnholds, J. A family of drug transporters: The multidrug resistance-associated proteins. J. Natl. Cancer Inst. 2000, 92, 1295–1302. [Google Scholar] [CrossRef]
- Ji, L.; Li, H.; Gao, P.; Shang, G.; Zhang, D.D.; Zhang, N.; Jiang, T. Nrf2 pathway regulates multidrug-resistance-associated protein 1 in small cell lung cancer. PLoS ONE 2013, 8, e63404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Wang, T.; Que, R.; Yang, J.; Wang, Z.; Jiang, X.; Wang, L. The potentially protective role of ATP-binding cassette transporters in preeclampsia via Nrf2. Pregnancy Hypertens. 2019, 18, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Dong, Y.; Chu, Y.; Guo, Y.; Li, L. The mechanisms of ferroptosis and its role in alzheimer’s disease. Front. Mol. Biosci. 2022, 9, 965064. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.; Berk, M.; Carvalho, A.F.; Maes, M.; Walker, A.J.; Puri, B.K. Why should neuroscientists worry about iron? The emerging role of ferroptosis in the pathophysiology of neuroprogressive diseases. Behav. Brain Res. 2018, 341, 154–175. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Chen, X.; Jiang, L.; Lu, B.; Yuan, M.; Zhu, D.; Zhu, H.; He, Q.; Yang, B.; Ying, M. DJ-1 suppresses ferroptosis through preserving the activity of S-adenosyl homocysteine hydrolase. Nat. Commun. 2020, 11, 1251. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.S.; Hwang, H.S.; Sohn, I.S.; Park, S.H. Expression of DJ-1 proteins in placentas from women with severe preeclampsia. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 168, 40–44. [Google Scholar] [CrossRef]
- Liao, T.; Xu, X.; Ye, X.; Yan, J. DJ-1 upregulates the Nrf2/GPX4 signal pathway to inhibit trophoblast ferroptosis in the pathogenesis of preeclampsia. Sci. Rep. 2022, 12, 2934. [Google Scholar] [CrossRef]
- Martinez-Casales, M.; Hernanz, R.; Alonso, M.J. Vascular and Macrophage Heme Oxygenase-1 in Hypertension: A Mini-Review. Front. Physiol. 2021, 12, 643435. [Google Scholar] [CrossRef]
- Kweider, N.; Fragoulis, A.; Rosen, C.; Pecks, U.; Rath, W.; Pufe, T.; Wruck, C.J. Interplay between vascular endothelial growth factor (VEGF) and nuclear factor erythroid 2-related factor-2 (Nrf2): Implications for preeclampsia. J. Biol. Chem. 2011, 286, 42863–42872. [Google Scholar] [CrossRef] [Green Version]
- Branch, D.W.; Mitchell, M.D.; Miller, E.; Palinski, W.; Witztum, J.L. Pre-eclampsia and serum antibodies to oxidised low-density lipoprotein. Lancet 1994, 343, 645–646. [Google Scholar] [CrossRef]
- Uzun, H.; Benian, A.; Madazli, R.; Topcuoglu, M.A.; Aydin, S.; Albayrak, M. Circulating oxidized low-density lipoprotein and paraoxonase activity in preeclampsia. Gynecol. Obstet. Investig. 2005, 60, 195–200. [Google Scholar] [CrossRef]
- Tsumita, T.; Maishi, N.; Annan, D.A.; Towfik, M.A.; Matsuda, A.; Onodera, Y.; Nam, J.M.; Hida, Y.; Hida, K. The oxidized-LDL/LOX-1 axis in tumor endothelial cells enhances metastasis by recruiting neutrophils and cancer cells. Int. J. Cancer 2022, 151, 944–956. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Itoh, K.; Ruiz, E.; Leake, D.S.; Unoki, H.; Yamamoto, M.; Mann, G.E. Role of Nrf2 in the regulation of CD36 and stress protein expression in murine macrophages: Activation by oxidatively modified LDL and 4-hydroxynonenal. Circ. Res. 2004, 94, 609–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chigusa, Y.; Tatsumi, K.; Kondoh, E.; Fujita, K.; Nishimura, F.; Mogami, H.; Konishi, I. Decreased lectin-like oxidized LDL receptor 1 (LOX-1) and low Nrf2 activation in placenta are involved in preeclampsia. J. Clin. Endocrinol. Metab. 2012, 97, E1862–E1870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Agarwal, A.; Sharma, R.K. The role of placental oxidative stress and lipid peroxidation in preeclampsia. Obstet. Gynecol. Surv. 2005, 60, 807–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serhan, C.N.; Chiang, N.; Van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Zhao, F.; Lin, F.; Xiang, H.; Wang, N.; Ye, D.; Huang, Y. Preeclampsia is associated with a deficiency of lipoxin A4, an endogenous anti-inflammatory mediator. Fertil. Steril. 2014, 102, 282–290.e284. [Google Scholar] [CrossRef]
- Lin, F.; Zeng, P.; Xu, Z.; Ye, D.; Yu, X.; Wang, N.; Tang, J.; Zhou, Y.; Huang, Y. Treatment of Lipoxin A(4) and its analogue on low-dose endotoxin induced preeclampsia in rat and possible mechanisms. Reprod. Toxicol. 2012, 34, 677–685. [Google Scholar] [CrossRef]
- Pang, H.; Yi, P.; Wu, P.; Liu, Z.; Liu, Z.; Gong, J.; Hao, H.; Cai, L.; Ye, D.; Huang, Y. Effect of lipoxin A4 on lipopolysaccharide-induced endothelial hyperpermeability. Sci. World J. 2011, 11, 1056–1067. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.Q.; Zhang, L. Hypoxia and the integrated stress response promote pulmonary hypertension and preeclampsia: Implications in drug development. Drug Discov. Today 2021, 26, 2754–2773. [Google Scholar] [CrossRef]
- Goldman-Wohl, D.; Yagel, S. Regulation of trophoblast invasion: From normal implantation to pre-eclampsia. Mol. Cell. Endocrinol. 2002, 187, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Hou, W.; Song, X.; Yu, Y.; Huang, J.; Sun, X.; Kang, R.; Tang, D. Ferroptosis: Process and function. Cell. Death Differ. 2016, 23, 369–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhang, L.; Zhou, X. Activation of Nrf2 signaling protects hypoxia-induced HTR-8/SVneo cells against ferroptosis. J. Obstet. Gynaecol. Res. 2021, 47, 3797–3806. [Google Scholar] [CrossRef] [PubMed]
- Qiu, D.; Wu, J.; Li, M.; Wang, L.; Zhu, X.; Chen, Y. Impaction of factors associated with oxidative stress on the pathogenesis of gestational hypertension and preeclampsia: A Chinese patients based study. Medicine 2021, 100, e23666. [Google Scholar] [CrossRef]
- Feng, H.; Wang, L.; Zhang, G.; Zhang, Z.; Guo, W. Oxidative stress activated by Keap-1/Nrf2 signaling pathway in pathogenesis of preeclampsia. Int. J. Clin. Exp. Pathol. 2020, 13, 382–392. [Google Scholar]
- Bayarsaikhan, G.; Bayarsaikhan, D.; Lee, J.; Lee, B. Targeting Scavenger Receptors in Inflammatory Disorders and Oxidative Stress. Antioxidants 2022, 11, 936. [Google Scholar] [CrossRef]
- Kalousova, M.; Zima, T.; Tesar, V.; Dusilova-Sulkova, S.; Skrha, J. Advanced glycoxidation end products in chronic diseases-clinical chemistry and genetic background. Mutat. Res. 2005, 579, 37–46. [Google Scholar] [CrossRef]
- Ungurianu, A.; Zanfirescu, A.; Gradinaru, D.; Ionescu-Tirgoviste, C.; Danciulescu Miulescu, R.; Margina, D. Interleukins and redox impairment in type 2 diabetes mellitus: Mini-review and pilot study. Curr. Med. Res. Opin. 2022, 38, 511–522. [Google Scholar] [CrossRef]
- Stanek, A.; Brozyna-Tkaczyk, K.; Myslinski, W. Oxidative Stress Markers among Obstructive Sleep Apnea Patients. Oxidative Med. Cell. Longev. 2021, 2021, 9681595. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, L.; Ouyang, X.; Jiang, Z.; Xie, Z.; Fan, L.; Zhu, D.; Li, L. Advanced oxidation protein products play critical roles in liver diseases. Eur. J. Clin. Investig. 2019, 49, e13098. [Google Scholar] [CrossRef] [Green Version]
- Cristani, M.; Speciale, A.; Saija, A.; Gangemi, S.; Minciullo, P.L.; Cimino, F. Circulating Advanced Oxidation Protein Products as Oxidative Stress Biomarkers and Progression Mediators in Pathological Conditions Related to Inflammation and Immune Dysregulation. Curr. Med. Chem. 2016, 23, 3862–3882. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.; Huang, Q.T.; Zhong, M.; Yin, Q. AOPPs (advanced oxidation protein products) promote apoptosis in trophoblastic cells through interference with NADPH oxidase signaling: Implications for preeclampsia. J. Matern.-Fetal Neonatal Med. 2015, 28, 1747–1755. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yin, Q.; Hu, H.; Chen, Q.; Huang, Q.; Zhong, M. AOPPs induce HTR-8/SVneo cell apoptosis by downregulating the Nrf-2/ARE/HO-1 anti-oxidative pathway: Potential implications for preeclampsia. Placenta 2021, 112, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Avellini, C.; Licini, C.; Lazzarini, R.; Gesuita, R.; Guerra, E.; Tossetta, G.; Castellucci, C.; Giannubilo, S.R.; Procopio, A.; Alberti, S.; et al. The trophoblast cell surface antigen 2 and miR-125b axis in urothelial bladder cancer. Oncotarget 2017, 8, 58642–58653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimi Dermani, F.; Datta, I.; Gholamzadeh Khoei, S. MicroRNA-452: A double-edged sword in multiple human cancers. Clin. Transl. Oncol. 2023, 25, 1189–1206. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xun, C.; Yu, C.H. Role of microRNA-regulated cancer stem cells in recurrent hepatocellular carcinoma. World J. Hepatol. 2022, 14, 1985–1996. [Google Scholar] [CrossRef] [PubMed]
- Poniewierska-Baran, A.; Sluczanowska-Glabowska, S.; Malkowska, P.; Sierawska, O.; Zadroga, L.; Pawlik, A.; Niedzwiedzka-Rystwej, P. Role of miRNA in Melanoma Development and Progression. Int. J. Mol. Sci. 2022, 24, 201. [Google Scholar] [CrossRef]
- Licini, C.; Avellini, C.; Picchiassi, E.; Mensa, E.; Fantone, S.; Ramini, D.; Tersigni, C.; Tossetta, G.; Castellucci, C.; Tarquini, F.; et al. Pre-eclampsia predictive ability of maternal miR-125b: A clinical and experimental study. Transl. Res. 2021, 228, 13–27. [Google Scholar] [CrossRef]
- Atic, A.I.; Thiele, M.; Munk, A.; Dalgaard, L.T. Circulating microRNAs associated with non-alcoholic fatty liver disease. Am. J. Physiol. Cell Physiol. 2023, 324, C588–C602. [Google Scholar] [CrossRef]
- Elkhawaga, S.Y.; Ismail, A.; Elsakka, E.G.E.; Doghish, A.S.; Elkady, M.A.; El-Mahdy, H.A. miRNAs as cornerstones in adipogenesis and obesity. Life Sci. 2023, 315, 121382. [Google Scholar] [CrossRef]
- Tofigh, R.; Hosseinpourfeizi, M.; Baradaran, B.; Teimourian, S.; Safaralizadeh, R. Rheumatoid arthritis and non-coding RNAs; how to trigger inflammation. Life Sci. 2023, 315, 121367. [Google Scholar] [CrossRef] [PubMed]
- Nohata, N.; Hanazawa, T.; Enokida, H.; Seki, N. microRNA-1/133a and microRNA-206/133b clusters: Dysregulation and functional roles in human cancers. Oncotarget 2012, 3, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Lu, Y.; Li, Z.; Wang, Q. microRNA-133: Expression, function and therapeutic potential in muscle diseases and cancer. Curr. Drug Targets 2014, 15, 817–828. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhao, J.; Tuazon, J.P.; Borlongan, C.V.; Yu, G. MicroRNA-133a and Myocardial Infarction. Cell. Transplant. 2019, 28, 831–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mafra, D.; Alvarenga, L.; Cardozo, L.; Stockler-Pinto, M.B.; Nakao, L.S.; Stenvinkel, P.; Shiels, P.G. Inhibiting BTB domain and CNC homolog 1 (Bach1) as an alternative to increase Nrf2 activation in chronic diseases. Biochim. Biophys. Acta Gen. Subj. 2022, 1866, 130129. [Google Scholar] [CrossRef]
- Guo, H.; Wang, Y.; Jia, W.; Liu, L. MiR-133a-3p relieves the oxidative stress induced trophoblast cell apoptosis through the BACH1/Nrf2/HO-1 signaling pathway. Physiol. Res. 2021, 70, 67–78. [Google Scholar] [CrossRef]
- Takeshita, N.; Hoshino, I.; Mori, M.; Akutsu, Y.; Hanari, N.; Yoneyama, Y.; Ikeda, N.; Isozaki, Y.; Maruyama, T.; Akanuma, N.; et al. Serum microRNA expression profile: miR-1246 as a novel diagnostic and prognostic biomarker for oesophageal squamous cell carcinoma. Br. J. Cancer 2013, 108, 644–652. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, X.; Zhang, L.; Shi, Y.; Wang, J.; Yan, H. Wnt/beta-catenin signaling pathway in trophoblasts and abnormal activation in preeclampsia (Review). Mol. Med. Rep. 2017, 16, 1007–1013. [Google Scholar] [CrossRef] [Green Version]
- Fournet-Dulguerov, N.; MacLusky, N.J.; Leranth, C.Z.; Todd, R.; Mendelson, C.R.; Simpson, E.R.; Naftolin, F. Immunohistochemical localization of aromatase cytochrome P-450 and estradiol dehydrogenase in the syncytiotrophoblast of the human placenta. J. Clin. Endocrinol. Metab. 1987, 65, 757–764. [Google Scholar] [CrossRef]
- Shen, X.; Kim, W.; Fujiwara, Y.; Simon, M.D.; Liu, Y.; Mysliwiec, M.R.; Yuan, G.C.; Lee, Y.; Orkin, S.H. Jumonji modulates polycomb activity and self-renewal versus differentiation of stem cells. Cell 2009, 139, 1303–1314. [Google Scholar] [CrossRef] [Green Version]
- Muralimanoharan, S.; Kwak, Y.T.; Mendelson, C.R. Redox-Sensitive Transcription Factor NRF2 Enhances Trophoblast Differentiation via Induction of miR-1246 and Aromatase. Endocrinology 2018, 159, 2022–2033. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.; Liu, J.; Chen, J.; Li, J.; Qiu, S.; Ma, J.; Lin, X.; Zhang, L.; Wu, J. Peptides of tetraspanin oncoprotein CD151 trigger active immunity against primary tumour and experimental lung metastasis. eBioMedicine 2019, 49, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Randhawa, P.K.; Rajakumar, A.; Futuro de Lima, I.B.; Gupta, M.K. Eugenol attenuates ischemia-mediated oxidative stress in cardiomyocytes via acetylation of histone at H3K27. Free Radic. Biol. Med. 2023, 194, 326–336. [Google Scholar] [CrossRef]
- Wang, Z.; Cai, B.; Cao, C.; Lv, H.; Dai, Y.; Zheng, M.; Zhao, G.; Peng, Y.; Gou, W.; Wang, J.; et al. Downregulation of CD151 induces oxidative stress and apoptosis in trophoblast cells via inhibiting ERK/Nrf2 signaling pathway in preeclampsia. Free Radic. Biol. Med. 2021, 164, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Hussong, M.; Borno, S.T.; Kerick, M.; Wunderlich, A.; Franz, A.; Sultmann, H.; Timmermann, B.; Lehrach, H.; Hirsch-Kauffmann, M.; Schweiger, M.R. The bromodomain protein BRD4 regulates the KEAP1/NRF2-dependent oxidative stress response. Cell Death Dis. 2014, 5, e1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Mi, Y.; Zhang, F.; Cheng, Y.; Wu, X. Suppression of bromodomain-containing protein 4 protects trophoblast cells from oxidative stress injury by enhancing Nrf2 activation. Hum. Exp. Toxicol. 2021, 40, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Tang, W.; Zhang, Z.; Yang, C.; Qian, L.; Xie, X.; Qiang, E.; Zhao, J.; Zhao, W.; Xiao, L.; et al. Procyanidin B2 mitigates endothelial endoplasmic reticulum stress through a PPARdelta-Dependent mechanism. Redox Biol. 2020, 37, 101728. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, R.; Xu, R.; Chu, Y.; Gu, W. Procyanidin B2 ameliorates endothelial dysfunction and impaired angiogenesis via the Nrf2/PPARgamma/sFlt-1 axis in preeclampsia. Pharmacol. Res. 2022, 177, 106127. [Google Scholar] [CrossRef]
- Ungvari, Z.; Bagi, Z.; Feher, A.; Recchia, F.A.; Sonntag, W.E.; Pearson, K.; de Cabo, R.; Csiszar, A. Resveratrol confers endothelial protection via activation of the antioxidant transcription factor Nrf2. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H18–H24. [Google Scholar] [CrossRef] [Green Version]
- Caldeira-Dias, M.; Viana-Mattioli, S.; de Souza Rangel Machado, J.; Carlstrom, M.; de Carvalho Cavalli, R.; Sandrim, V.C. Resveratrol and grape juice: Effects on redox status and nitric oxide production of endothelial cells in in vitro preeclampsia model. Pregnancy Hypertens. 2021, 23, 205–210. [Google Scholar] [CrossRef]
- George, E.M.; Colson, D.; Dixon, J.; Palei, A.C.; Granger, J.P. Heme Oxygenase-1 Attenuates Hypoxia-Induced sFlt-1 and Oxidative Stress in Placental Villi through Its Metabolic Products CO and Bilirubin. Int. J. Hypertens. 2012, 2012, 486053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, R.; Acharya, R.; Delpachitra, P.; Hobson, S.; Sobey, C.G.; Drummond, G.R.; Wallace, E.M. Activin and NADPH-oxidase in preeclampsia: Insights from in vitro and murine studies. Am. J. Obstet. Gynecol. 2015, 212, 86.e1–86.e12. [Google Scholar] [CrossRef] [PubMed]
- Gurusinghe, S.; Cox, A.G.; Rahman, R.; Chan, S.T.; Muljadi, R.; Singh, H.; Leaw, B.; Mockler, J.C.; Marshall, S.A.; Murthi, P.; et al. Resveratrol mitigates trophoblast and endothelial dysfunction partly via activation of nuclear factor erythroid 2-related factor-2. Placenta 2017, 60, 74–85. [Google Scholar] [CrossRef]
- Guo, H.; Wang, Y.; Liu, D. Silibinin ameliorats H2O2-induced cell apoptosis and oxidative stress response by activating Nrf2 signaling in trophoblast cells. Acta Histochem. 2020, 122, 151620. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Choi, K.J.; Cho, S.J.; Yun, S.M.; Jeon, J.P.; Koh, Y.H.; Song, J.; Johnson, G.V.; Jo, C. Fisetin stimulates autophagic degradation of phosphorylated tau via the activation of TFEB and Nrf2 transcription factors. Sci. Rep. 2016, 6, 24933. [Google Scholar] [CrossRef] [Green Version]
- Ehren, J.L.; Maher, P. Concurrent regulation of the transcription factors Nrf2 and ATF4 mediates the enhancement of glutathione levels by the flavonoid fisetin. Biochem. Pharmacol. 2013, 85, 1816–1826. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Chen, J.; Hu, J. Protective effect of Fisetin on the lipopolysaccharide-induced preeclampsia-like rats. Hypertens. Pregnancy 2022, 41, 23–30. [Google Scholar] [CrossRef]
- Savla, S.R.; Laddha, A.P.; Kulkarni, Y.A. Pharmacology of apocynin: A natural acetophenone. Drug. Metab. Rev. 2021, 53, 542–562. [Google Scholar] [CrossRef]
- Barbieri, S.S.; Cavalca, V.; Eligini, S.; Brambilla, M.; Caiani, A.; Tremoli, E.; Colli, S. Apocynin prevents cyclooxygenase 2 expression in human monocytes through NADPH oxidase and glutathione redox-dependent mechanisms. Free Radic. Biol. Med. 2004, 37, 156–165. [Google Scholar] [CrossRef]
- Ju, Y.; Feng, Y.; Hou, X.; Wu, L.; Yang, H.; Zhang, H.; Ma, Y. Combined apocyanin and aspirin treatment activates the PI3K/Nrf2/HO-1 signaling pathway and ameliorates preeclampsia symptoms in rats. Hypertens. Pregnancy 2022, 41, 39–50. [Google Scholar] [CrossRef]
- Mizuno, K.; Kume, T.; Muto, C.; Takada-Takatori, Y.; Izumi, Y.; Sugimoto, H.; Akaike, A. Glutathione biosynthesis via activation of the nuclear factor E2-related factor 2 (Nrf2)—Antioxidant-response element (ARE) pathway is essential for neuroprotective effects of sulforaphane and 6-(methylsulfinyl) hexyl isothiocyanate. J. Pharmacol. Sci. 2011, 115, 320–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, A.G.; Gurusinghe, S.; Abd Rahman, R.; Leaw, B.; Chan, S.T.; Mockler, J.C.; Murthi, P.; Marshall, S.A.; Lim, R.; Wallace, E.M. Sulforaphane improves endothelial function and reduces placental oxidative stress in vitro. Pregnancy Hypertens. 2019, 16, 1–10. [Google Scholar] [CrossRef]
- Mashmoul, M.; Azlan, A.; Mohtarrudin, N.; Yusof, B.N.M.; Khaza’ai, H. Saffron extract and crocin reduced biomarkers associated with obesity in rats fed a high-fat diet. Malays. J. Nutr. 2017, 23, 117–127. [Google Scholar]
- Chen, X.; Huang, J.; Lv, Y.; Chen, Y.; Rao, J. Crocin exhibits an antihypertensive effect in a rat model of gestational hypertension and activates the Nrf-2/HO-1 signaling pathway. Hypertens. Res. 2021, 44, 642–650. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Cai, X.; Qian, Q.; Zhuang, Q.; Yang, W.; Zhang, X.; Zhao, L. Astragaloside IV protects endothelial progenitor cells from the damage of ox-LDL via the LOX-1/NLRP3 inflammasome pathway. Drug Des. Devel Ther. 2019, 13, 2579–2589. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.L.; Zhang, Q.Z.; Wang, Y.R.; Fu, L.N.; Han, J.S.; Zhang, J.; Wang, B.M. Astragaloside IV Improves High-Fat Diet-Induced Hepatic Steatosis in Nonalcoholic Fatty Liver Disease Rats by Regulating Inflammatory Factors Level via TLR4/NF-kappaB Signaling Pathway. Front. Pharmacol. 2020, 11, 605064. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, R.; Xing, B.; Zhou, L.; Zhang, P.; Song, L. Astragaloside IV ameliorates preeclampsia-induced oxidative stress through the Nrf2/HO-1 pathway in a rat model. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E904–E911. [Google Scholar] [CrossRef]
- Ikemoto, K.; Mori, S.; Mukai, K. Synthesis and crystal structure of pyrroloquinoline quinol (PQQH(2)) and pyrroloquinoline quinone (PQQ). Acta Crystallogr. B Struct. Sci. Cryst. Eng. Mater. 2017, 73, 489–497. [Google Scholar] [CrossRef]
- Hwang, P.; Willoughby, D.S. Mechanisms Behind Pyrroloquinoline Quinone Supplementation on Skeletal Muscle Mitochondrial Biogenesis: Possible Synergistic Effects with Exercise. J. Am. Coll. Nutr. 2018, 37, 738–748. [Google Scholar] [CrossRef]
- Wang, H.; Li, M.; Chen, P.; Shi, X. Anti-inflammatory and Antioxidant Effects of Pyrroloquinoline Quinone in L-NAME-Induced Preeclampsia-Like Rat Model. Reprod. Sci. 2022, 29, 578–585. [Google Scholar] [CrossRef]
- Kim, J.; Shin, S.H.; Kang, J.K.; Kim, J.W. HX-1171 attenuates pancreatic beta-cell apoptosis and hyperglycemia-mediated oxidative stress via Nrf2 activation in streptozotocin-induced diabetic model. Oncotarget 2018, 9, 24260–24271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yada, H.; Hirose, M.; Tamano, S.; Kawabe, M.; Sano, M.; Takahashi, S.; Futakuchi, M.; Miki, T.; Shirai, T. Effects of antioxidant 1-O-hexyl-2,3,5-trimethylhydroquinone or ascorbic acid on carcinogenesis induced by administration of aminopyrine and sodium nitrite in a rat multi-organ carcinogenesis model. Jpn. J. Cancer Res. 2002, 93, 1299–1307. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kang, J.K.; Lee, M.K. 1-O-Hexyl-2,3,5-Trimethylhydroquinone Ameliorates l-DOPA-Induced Cytotoxicity in PC12 Cells. Molecules 2019, 24, 867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Gong, Y.; Rao, J.; Yang, Q.; Gao, N.; Li, G.; Ma, Y. 1-O-Hexyl-2,3,5-Trimethylhydroquinone Ameliorates the Development of Preeclampsia through Suppression of Oxidative Stress and Endothelial Cell Apoptosis. Oxidative Med. Cell. Longev. 2021, 2021, 8839394. [Google Scholar] [CrossRef] [PubMed]
- Hornos Carneiro, M.F.; Colaiacovo, M.P. Beneficial antioxidant effects of Coenzyme Q10 on reproduction. Vitam. Horm. 2023, 121, 143–167. [Google Scholar] [CrossRef]
- Mantle, D.; Lopez-Lluch, G.; Hargreaves, I.P. Coenzyme Q10 Metabolism: A Review of Unresolved Issues. Int. J. Mol. Sci. 2023, 24, 2585. [Google Scholar] [CrossRef]
- Witting, P.K.; Pettersson, K.; Letters, J.; Stocker, R. Anti-atherogenic effect of coenzyme Q10 in apolipoprotein E gene knockout mice. Free Radic. Biol. Med. 2000, 29, 295–305. [Google Scholar] [CrossRef]
- Singh, R.B.; Shinde, S.N.; Chopra, R.K.; Niaz, M.A.; Thakur, A.S.; Onouchi, Z. Effect of coenzyme Q10 on experimental atherosclerosis and chemical composition and quality of atheroma in rabbits. Atherosclerosis 2000, 148, 275–282. [Google Scholar] [CrossRef]
- Palan, P.R.; Shaban, D.W.; Martino, T.; Mikhail, M.S. Lipid-soluble antioxidants and pregnancy: Maternal serum levels of coenzyme Q10, alpha-tocopherol and gamma-tocopherol in preeclampsia and normal pregnancy. Gynecol. Obstet. Investig. 2004, 58, 8–13. [Google Scholar] [CrossRef]
- Li, L.; Li, H.; Zhou, Q.; Lu, Y.; Chen, P.; Wang, X.; Zhao, H. Implication of nuclear factor-erythroid 2-like 2/heme oxygenase 1 pathway in the protective effects of coenzyme Q10 against preeclampsia-like in a rat model. Microcirculation 2020, 27, e12651. [Google Scholar] [CrossRef]
- Lanoix, D.; Beghdadi, H.; Lafond, J.; Vaillancourt, C. Human placental trophoblasts synthesize melatonin and express its receptors. J. Pineal Res. 2008, 45, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Socaciu, A.I.; Ionut, R.; Socaciu, M.A.; Ungur, A.P.; Barsan, M.; Chiorean, A.; Socaciu, C.; Rajnoveanu, A.G. Melatonin, an ubiquitous metabolic regulator: Functions, mechanisms and effects on circadian disruption and degenerative diseases. Rev. Endocr. Metab. Disord. 2020, 21, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Zeng, K.; Gao, Y.; Wan, J.; Tong, M.; Lee, A.C.; Zhao, M.; Chen, Q. The reduction in circulating levels of melatonin may be associated with the development of preeclampsia. J. Hum. Hypertens. 2016, 30, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Cai, X.; Li, J.; Huang, Y.; Liu, H.; He, J.; Fang, Z.; Feng, B.; Tang, J.; Lin, Y.; et al. Effects of Melatonin Supplementation during Pregnancy on Reproductive Performance, Maternal-Placental-Fetal Redox Status, and Placental Mitochondrial Function in a Sow Model. Antioxidants 2021, 10, 1867. [Google Scholar] [CrossRef]
- Zuo, J.; Jiang, Z. Melatonin attenuates hypertension and oxidative stress in a rat model of L-NAME-induced gestational hypertension. Vasc. Med. 2020, 25, 295–301. [Google Scholar] [CrossRef]
- Henderson, J.T.; O’Connor, E.; Whitlock, E.P. Low-dose aspirin for prevention of morbidity and mortality from preeclampsia. Ann. Intern. Med. 2014, 161, 613–614. [Google Scholar] [CrossRef]
- Hobson, S.R.; Gurusinghe, S.; Lim, R.; Alers, N.O.; Miller, S.L.; Kingdom, J.C.; Wallace, E.M. Melatonin improves endothelial function in vitro and prolongs pregnancy in women with early-onset preeclampsia. J. Pineal Res. 2018, 65, e12508. [Google Scholar] [CrossRef]
- Ye, F.; Li, X.; Li, L.; Yuan, J.; Chen, J. t-BHQ Provides Protection against Lead Neurotoxicity via Nrf2/HO-1 Pathway. Oxidative Med. Cell. Longev. 2016, 2016, 2075915. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Yang, B.; Wang, L.; Xue, P.; Deng, B.; Zhang, G.; Jiang, S.; Zhang, M.; Liu, M.; Pi, J.; et al. Curcumin protects human keratinocytes against inorganic arsenite-induced acute cytotoxicity through an NRF2-dependent mechanism. Oxidative Med. Cell. Longev. 2013, 2013, 412576. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhou, J.; Ye, Y.; Liu, Q.; Wang, X.; Zhang, N.; Wang, X. Increased Heme Oxygenase-1 and Nuclear Factor Erythroid 2-Related Factor-2 in the Placenta Have a Cooperative Action on Preeclampsia. Gynecol. Obstet. Investig. 2016, 81, 543–551. [Google Scholar] [CrossRef]
- Chartoumpekis, D.; Ziros, P.G.; Psyrogiannis, A.; Kyriazopoulou, V.; Papavassiliou, A.G.; Habeos, I.G. Simvastatin lowers reactive oxygen species level by Nrf2 activation via PI3K/Akt pathway. Biochem. Biophys. Res. Commun. 2010, 396, 463–466. [Google Scholar] [CrossRef] [PubMed]
- Chigusa, Y.; Kawasaki, K.; Kondoh, E.; Mogami, H.; Ujita, M.; Fujita, K.; Tatsumi, K.; Takeda, S.; Konishi, I. Simvastatin inhibits oxidative stress via the activation of nuclear factor erythroid 2-related factor 2 signaling in trophoblast cells. J. Obstet. Gynaecol. Res. 2016, 42, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liang, B.; Meng, F.; Li, H. Effects of Nrf-2 expression in trophoblast cells and vascular endothelial cells in preeclampsia. Am. J. Transl. Res. 2021, 13, 1006–1021. [Google Scholar] [PubMed]



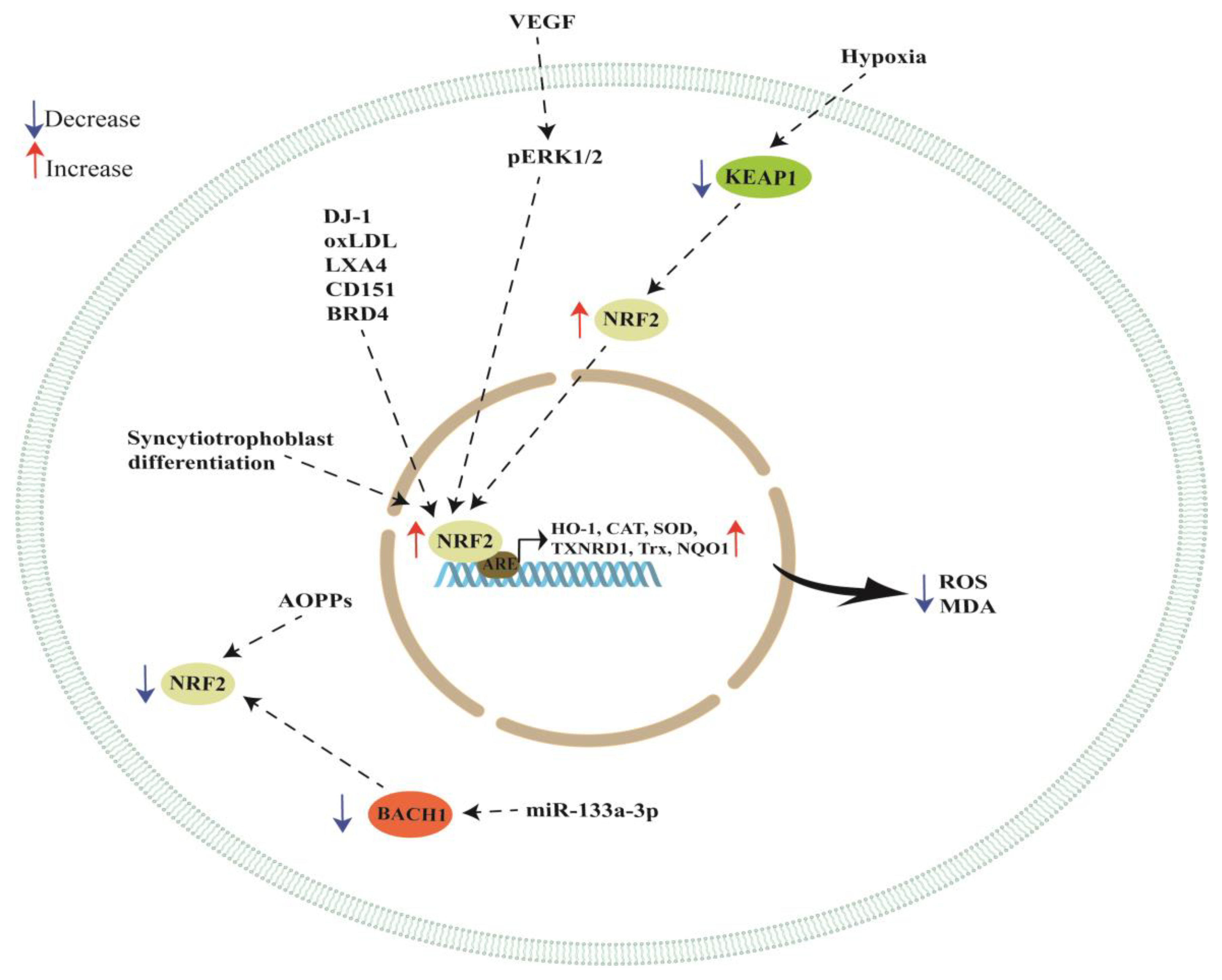{kind=link}
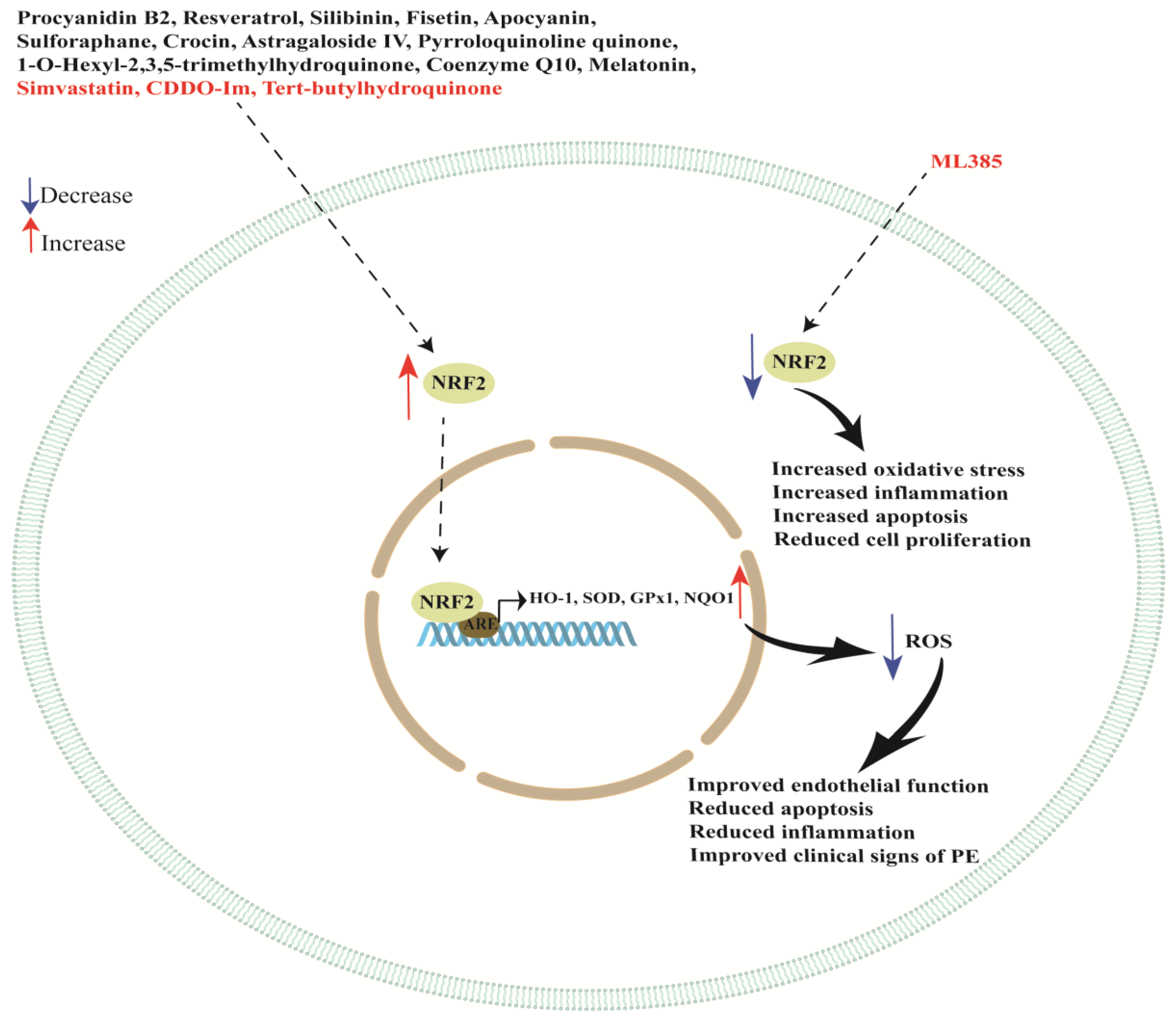{kind=link}
| Modulator | Model Studied | Results | Reference |
|---|---|---|---|
| DJ-1 | BeWo cells | NRF2 and GPX4 expression was significantly reduced when DJ-1 was knocked down in BeWo cells. Cell death was significantly increased in DJ-1-deficient cells when these cells were treated with RSL3, a ferroptosis inducer. | [67] |
| VEGF | BeWo cells | VEGF activated NRF2, increasing thioredoxin (Trx), thioredoxin reductase (TXNRD1) and heme oxygenase-1 (HO-1). VEGF activated NRF2 in an ERK1/2-dependent manner, increasing HO-1 expression then augmenting the production of carbon monoxide, which increased VEGF expression. | [69] |
| oxLDL | JAR cells and placental explants | Treatment with oxLDL increased NRF2 and HO-1 expression while the blockade of LOX-1 with TS92 inhibited the increase in HO-1 expression induced by oxLDL treatment. | [74] |
| LXA4 | HUVEC | LXA4 inhibited LPS-triggered ROS production, promoting the expression of NRF2 and improving vascular permeability under oxidant stimuli. | [79] |
| Hypoxia | HTR-8/SVneo cells | NRF2 overexpression in hypoxia-induced cells reduced the levels of MDA and ROS, and decreased ferroptosis. | [83] |
| Hypoxia | HTR-8/SVneo cells | Hypoxia reduced the activity of CAT, GSH-Px and SOD enzymes and increased NRF2 and HO-1 expression while decreasing KEAP1 expression. The activity of SOD, GSH-Px and CAT in placental tissues of patients with PE was lower compared to normal placental tissues. NRF2 and HO-1 expression in preeclamptic placentas was higher compared to normal pregnancies while KEAP1 expression was lower in PE placentas compared to the normal ones. Silencing NRF2 in HTR8/SVneo cells under hypoxic conditions reduced the activities of CAT, GSH-Px and SOD. | [84] |
| Hypoxia | PE placentas and HTR-8/SVneo cells | Lower CAT, GSH-Px and SOD activity in HTR8/SVneo cells under hypoxic conditions and in PE placentas. Increased NRF2 and HO-1 expression together with a reduced expression of KEAP1 under hypoxic conditions and in PE placentas. | [85] |
| AOPPs | HTR-8/SVneo cells | AOPPs increased apoptosis and inhibited the NRF2/ARE/HO-1 pathway. NRF2 silencing aggravated the AOPP-induced cell apoptosis, activating p53 and the caspase cascade while NRF2 overexpression showed cytoprotective effects by increasing HO-1 expression. | [93] |
| miR-133a-3p | HTR-8/SVneo cells exposed to H2O2 | Transfecting cells with miR-133a-3p under an oxidative stress condition reduced ROS, MDA levels and apoptosis. MiR-133a-3p inhibited BACH1 (a NRF2 repressor), increasing NRF2 activation and HO-1 expression. | [106] |
| Syncytiotrophoblast differentiation | PE placentas and primary trophoblast cells | NRF2, CYP191A mRNAs and miR-1246 levels were upregulated during syncytiotrophoblast differentiation of trophoblast cells and significantly reduced by hypoxia and in PE placentas. JARID2, Axin-2 and GSK3β expression was significantly downregulated during syncytiotrophoblast differentiation. Silencing of NRF2 in cytotrophoblast cells inhibited miR-1246 and CYP19A1 due to the binding of NRF2 to the miR-1246 and CYP191A promoters. | [111] |
| CD151 | PE placentas, HTR-8/SVneo cells and mice | PE placentas showed reduced expression of CD151, HO-1, NQO1, GCLC and SOD-1. Overexpression of CD151 in HTR-8/SVneo cells enhanced HO-1, NQO1, GCLC and SOD-1 expression. Tail intravenous injection of siCD151 in pregnant mice led to a PE-like phenotype, hypertension and proteinuria. The expression of NRF2, pERK1/2, HO-1, NQO1, GCLC and SOD-1 was decreased in mice and HTR8/SVneo cells when CD151 was silenced. The beneficial effect of CD151 in HTR8/SVneo cells was inhibited when ERK and NRF2 signaling was blocked with synthetic inhibitors. | [114] |
| BRD4 | HTR8/SVneo exposed to H2O2 | BRD4 inhibition attenuated oxidative stress injury by enhancing NRF2 activation via the downregulation of KEAP1. | [116] |
| Modulator | Model Studied | Results | Reference |
|---|---|---|---|
| Procyanidin B2 | Placental explants and a PE rat model | Procyanidin B2 inhibits sFlt-1 secretion and ameliorates endothelial dysfunction and impaired angiogenesis via the NRF2/PPARγ axis. | [118] |
| Resveratrol | HUVEC incubated with plasma from PE patients | Antioxidant response element (ARE) activity was increased. The addition of resveratrol by NRF2 activation also occurred. | [120] |
| Resveratrol | Term placental explants and HUVEC treated with TNF-α and resveratrol | NRF2 knockdown abolished some of the protective effects of resveratrol on endothelial cells, but not in primary trophoblast cells. | [123] |
| Silibinin | HTR8/SVneo exposed to H2O2 | Silibinin protects the trophoblast from apoptosis, enhancing the activation of NRF2. | [124] |
| Fisetin | PE rat model | Reduction in hypertension and proteinuria; reduction in TNF-α, IL-6, IL-1β, MDA and the sFlt-1/PlGF ratio; and promoting the NRF2/HO-1 pathway in placental tissues. | [127] |
| Apocyanin | PE rat model | Reduction in preeclampsia symptoms with combined treatment of apocyanin and aspirin by activating the PI3K/NRF2/HO-1 pathway. | [130] |
| Sulforaphane | HUVEC and placental explants | In HUVEC, reduction in endothelin-1, VCAM1, ICAM1 and E-selectin. In placental explants, reduction in sFlt-1, endoglin and activin A. In HUVEC, induction of activation and nuclear translocation of NRF2, and induction of HO-1. NRF2 silencing blocked some but not all of sulforaphane’s effects and did not prevent inhibition of trophoblast secretion of sFlt-1 or activin A. | [132] |
| Crocin | PE rat model | Crocin upregulated protein levels of NRF2 and HO-1. | [134] |
| Astragaloside IV | PE rat model | Improvements in clinical signs of preeclampsia, reduction in placental cellular oxidative stress and strengthening of the NRF2/HO-1 signaling pathway in placental tissues. | [137] |
| Pyrroloquinoline quinone | PE rat model | Pyrroloquinoline quinone improved the antioxidation effect in preeclampsia models, activating the NRF2 pathway. | [140] |
| 1-O-hexyl-2,3,5-trimethylhydroquinone (HTHQ) | PE mouse model | HTHQ treatment induced NRF2 expression and nuclear translocation, increasing HO-1 expression in placentas. | [144] |
| Coenzyme Q10 | PE rat model | Coenzyme Q10 protected the rats from preeclampsia through activating the NRF2/HO-1 pathway. | [150] |
| Melatonin | Pregnant sows | Increase in mRNA levels of antioxidant-related genes involved in the NRF2/ARE pathway (NRF2, SOD, GPx1 and NQO1). | [154] |
| Melatonin | PE rat model | Increased NRF2, PlGF and HO-1 placental levels with reduction in blood pressure and urine protein content, and recovery in the fetus alive ratio, fetal weight and fetal weight/placental weight ratio. | [155] |
| Melatonin | Placental explants | Improved oxidative stress, presumably due to the potentiation of NRF2 and HO-1. | [157] |
| Tert-butylhydroquinone (tBHQ) | HTR-8/SVneo | Increased NRF2 protein and HO-1 mRNA expression after stimulation with tBHQ. HO-1 was located in the cytoplasm and NRF2 was located in both the nucleus and cytoplasm. | [160] |
| Simvastatin | JAR cells exposed to hypoxia and treated with diethyl maleate (DEM) | In hypoxia conditions, activation of NRF2 signaling depending on KEAP1 inhibition. | [162] |
| ML385 and CDDO-Im | PE rat model, HTR-8/SVneo and hESC cells | ML385 treatment reduced SBP and proteinuria in PE rats while treatment with CDDO-Im increased proteinuria and systolic blood pressure, worsening PE. HO-1 expression decreased in the PE group compared with the control group while it increased after CDDO-Im treatment compared with the PE group. ML385 did not alter HO-1 expression in placental tissue. NRF2 inhibition increased CCL2, IL-1β, TNF-α, AT1R and ROS in the embryonic tissues. NRF2 knockdown in HTR-8/SVneo and hESC cells suppressed cell proliferation, improved apoptosis and invasion and increased ROS and HO-1 expression. | [163] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tossetta, G.; Fantone, S.; Piani, F.; Crescimanno, C.; Ciavattini, A.; Giannubilo, S.R.; Marzioni, D. Modulation of NRF2/KEAP1 Signaling in Preeclampsia. Cells 2023, 12, 1545. https://doi.org/10.3390/cells12111545
Tossetta G, Fantone S, Piani F, Crescimanno C, Ciavattini A, Giannubilo SR, Marzioni D. Modulation of NRF2/KEAP1 Signaling in Preeclampsia. Cells. 2023; 12(11):1545. https://doi.org/10.3390/cells12111545
Chicago/Turabian StyleTossetta, Giovanni, Sonia Fantone, Federica Piani, Caterina Crescimanno, Andrea Ciavattini, Stefano Raffaele Giannubilo, and Daniela Marzioni. 2023. "Modulation of NRF2/KEAP1 Signaling in Preeclampsia" Cells 12, no. 11: 1545. https://doi.org/10.3390/cells12111545