
Pharmacometabolic Effects of Pteryxin and Valproate on Pentylenetetrazole-Induced Seizures in Zebrafish Reveal Vagus Nerve Stimulation

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish
2.2. Animal Treatment
2.3. Toxicological Evaluation
2.4. Locomotor Tracking
2.5. Local Field Potential
2.6. LC-MS/MS Quantification and Estimation of Bioavailability of VPN (as VPA) and PTX in Larvae
2.6.1. Sample Extraction
2.6.2. LC-ESI-MS/MS Measurements
2.7. Amino Acids and Neurotransmitters Quantification
2.7.1. Sample Preparation and Extraction
2.7.2. LC-MS/MS Conditions
2.8. Statistical Analysis
3. Results and Discussion
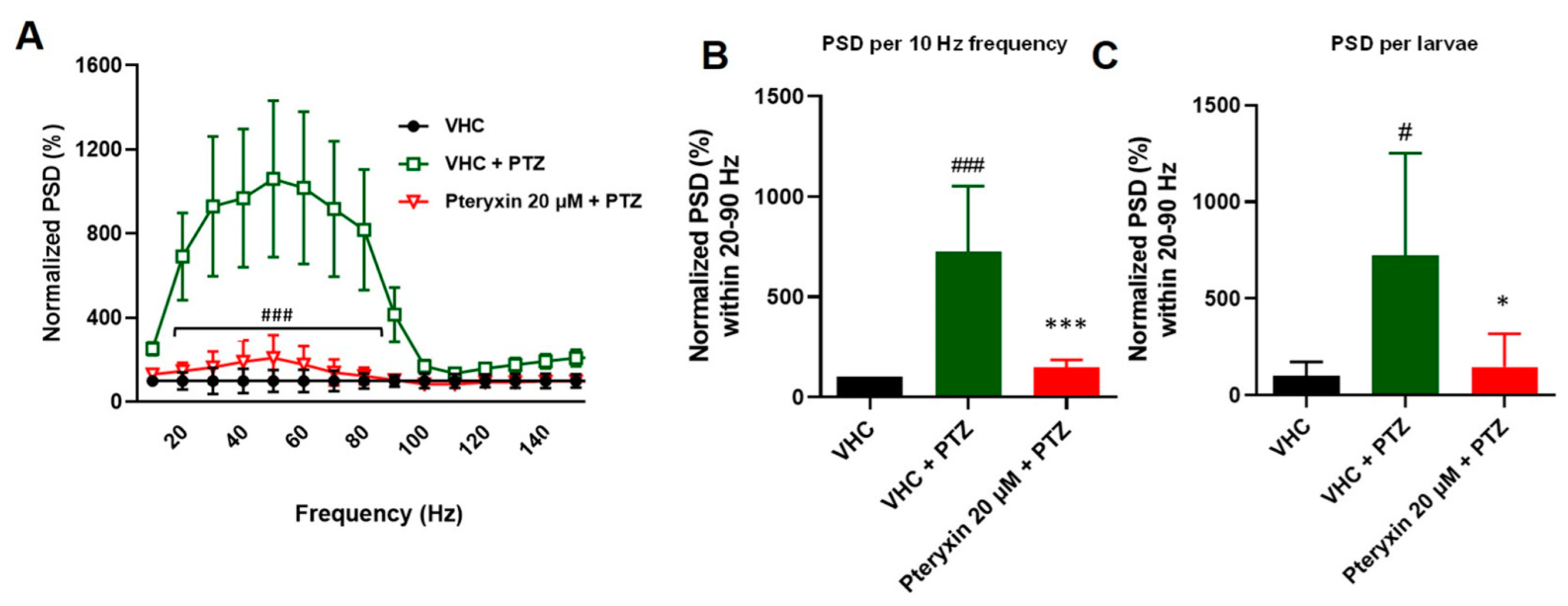
3.1. Determination of Anticonvulsant Activity of VPN and PTX in the PTZ Epilepsy-like Zebrafish Model
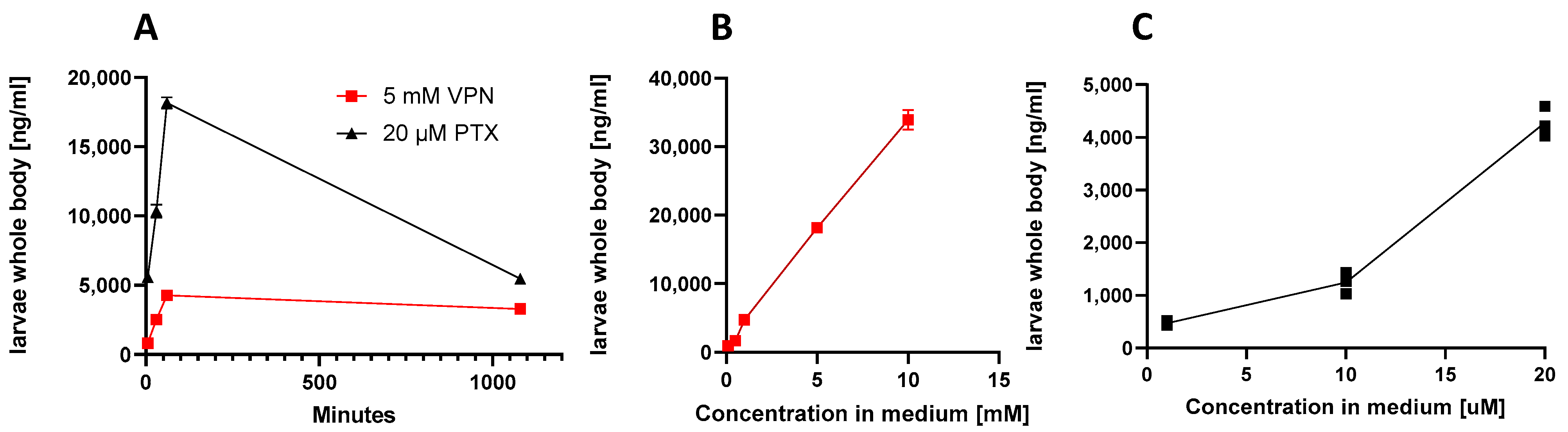
3.2. Measurment of Compound Concentration in Larvae Using LC-MS/MS Quantification
3.3. Pharmacometabolic Changes on Amino Acid and Neurotransmitter Levels in Larvae Measured by LC-MS/MS
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Horzmann, K.A.; Freeman, J.L. Making Waves: New Developments in Toxicology with the Zebrafish. Toxicol. Sci. 2018, 163, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Brox, S.; Ritter, A.P.; Küster, E.; Reemtsma, T. A Quantitative HPLC-MS/MS Method for Studying Internal Concentrations and Toxicokinetics of 34 Polar Analytes in Zebrafish (Danio Rerio) Embryos. Anal. Bioanal. Chem. 2014, 406, 4831–4840. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Heylen, L.; Partoens, M.; Mills, J.D.; Kaminski, R.M.; Godard, P.; Gillard, M.; de Witte, P.A.M.; Siekierska, A. Connectivity Mapping Using a Novel Sv2a Loss-of-Function Zebrafish Epilepsy Model as a Powerful Strategy for Anti-Epileptic Drug Discovery. Front. Mol. Neurosci. 2022, 15, 881933. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A.V.; Stewart, A.M.; Gerlai, R. Zebrafish as an Emerging Model for Studying Complex Brain Disorders. Trends Pharmacol. Sci. 2014, 35, 63–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basnet, R.M.; Zizioli, D.; Taweedet, S.; Finazzi, D.; Memo, M. Zebrafish Larvae as a Behavioral Model in Neuropharmacology. Biomedicines 2019, 7, 23. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, F.; Noldus, L.P.J.J.; Tegelenbosch, R.A.J.; Richardson, M.K. Zebrafish Embryos and Larvae in Behavioural Assays. Behaviour 2012, 149, 1241–1281. [Google Scholar] [CrossRef]
- Kozioł, E.; Luca, S.V.; Marcourt, L.; Nour, M.; Hnawia, E.; Jakubowicz-Gil, J.; Paduch, R.; Mroczek, T.; Wolfender, J.-L.; Skalicka-Woźniak, K. Efficient Extraction and Isolation of Skimmianine from New Caledonian Plant Medicosma Leratii and Evaluation of Its Effects on Apoptosis, Necrosis, and Autophagy. Phytochem. Lett. 2019, 30, 224–230. [Google Scholar] [CrossRef]
- Maciąg, M.; Michalak, A.; Skalicka-Woźniak, K.; Zykubek, M.; Ciszewski, A.; Budzyńska, B. Zebrafish and Mouse Models for Anxiety Evaluation—A Comparative Study with Xanthotoxin as a Model Compound. Brain Res. Bull. 2020, 165, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Dang, M.; Fogley, R.; Zon, L.I. Identifying Novel Cancer Therapies Using Chemical Genetics and Zebrafish. Adv. Exp. Med. Biol. 2016, 916, 103–124. [Google Scholar] [CrossRef]
- Kislyuk, S.; Kroonen, J.; Adams, E.; Augustijns, P.; de Witte, P.; Cabooter, D. Development of a Sensitive and Quantitative UHPLC-MS/MS Method to Study the Whole-Body Uptake of Pharmaceuticals in Zebrafish. Talanta 2017, 174, 780–788. [Google Scholar] [CrossRef]
- Halbach, K.; Ulrich, N.; Goss, K.U.; Seiwert, B.; Wagner, S.; Scholz, S.; Luckenbach, T.; Bauer, C.; Schweiger, N.; Reemtsma, T. Yolk Sac of Zebrafish Embryos as Backpack for Chemicals? Environ. Sci. Technol. 2020, 54, 10159–10169. [Google Scholar] [CrossRef] [PubMed]
- Copmans, D.; Orellana-Paucar, A.M.; Steurs, G.; Zhang, Y.; Ny, A.; Foubert, K.; Exarchou, V.; Siekierska, A.; Kim, Y.; De Borggraeve, W.; et al. Methylated Flavonoids as Anti-Seizure Agents: Naringenin 4′,7-Dimethyl Ether Attenuates Epileptic Seizures in Zebrafish and Mouse Models. Neurochem. Int. 2018, 112, 124–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarin, M.; Faelens, R.; Giusti, A.; De Croze, N.; Léonard, M.; Cabooter, D.; Annaert, P.; de Witte, P.; Ny, A. Spatiotemporal Imaging and Pharmacokinetics of Fluorescent Compounds in Zebrafish Eleuthero-Embryos after Different Routes of Administration. Sci. Rep. 2021, 11, 12229. [Google Scholar] [CrossRef] [PubMed]
- Afrikanova, T.; Serruys, A.S.K.; Buenafe, O.E.M.; Clinckers, R.; Smolders, I.; de Witte, P.A.M.; Crawford, A.D.; Esguerra, C.V. Validation of the Zebrafish Pentylenetetrazol Seizure Model: Locomotor versus Electrographic Responses to Antiepileptic Drugs. PLoS ONE 2013, 8, e54166. [Google Scholar] [CrossRef] [Green Version]
- Orhan, I.E.; Senol Deniz, F.S.; Trædal-Henden, S.; Cerón-Carrasco, J.P.; den Haan, H.; Peña-García, J.; Pérez-Sánchez, H.; Emerce, E.; Skalicka-Wozniak, K. Profiling Auspicious Butyrylcholinesterase Inhibitory Activity of Two Herbal Molecules: Hyperforin and Hyuganin C. Chem. Biodivers. 2019, 16, e1900017. [Google Scholar] [CrossRef]
- Zheng, Y.M.; Chen, B.; Jiang, J.D.; Zhang, J.P. Syntaxin 1B Mediates Berberine’s Roles in Epilepsy-like Behavior in a Pentylenetetrazole-Induced Seizure Zebrafish Model. Front. Mol. Neurosci. 2018, 11, 378. [Google Scholar] [CrossRef] [Green Version]
- Koziol, E.; Jóźwiak, K.; Budzyńska, B.; de Witte, P.A.M.; Copmans, D.; Skalicka-woźniak, K. Comparative Antiseizure Analysis of Diverse Natural Coumarin Derivatives in Zebrafish. Int. J. Mol. Sci. 2021, 22, 11420. [Google Scholar] [CrossRef]
- Adams, M.; Schneider, S.V.; Kluge, M.; Kessler, M.; Hamburger, M. Epilepsy in the Renaissance: A Survey of Remedies from 16th and 17th Century German Herbals. J. Ethnopharmacol. 2012, 143, 1–13. [Google Scholar] [CrossRef]
- Luszczki, J.J.; Andres-Mach, M.; Cisowski, W.; Mazol, I.; Glowniak, K.; Czuczwar, S.J. Osthole Suppresses Seizures in the Mouse Maximal Electroshock Seizure Model. Eur. J. Pharmacol. 2009, 607, 107–109. [Google Scholar] [CrossRef]
- Kozioł, E.; Deniz, F.S.S.; Orhan, I.E.; Marcourt, L.; Budzyńska, B.; Wolfender, J.L.; Crawford, A.D.; Skalicka-Woźniak, K. High-Performance Counter-Current Chromatography Isolation and Initial Neuroactivity Characterization of Furanocoumarin Derivatives from Peucedanum Alsaticum L (Apiaceae). Phytomedicine 2019, 15, 259–264. [Google Scholar] [CrossRef]
- Romoli, M.; Mazzocchetti, P.; D’Alonzo, R.; Siliquini, S.; Rinaldi, V.E.; Verrotti, A.; Calabresi, P.; Costa, C. Valproic Acid and Epilepsy: From Molecular Mechanisms to Clinical Evidences. Curr. Neuropharmacol. 2018, 17, 926–946. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.D.; Dager, S.R.; Parow, A.; Hirashima, F.; Demopulos, C.; Stoll, A.L.; Lyoo, I.K.; Dunner, D.L.; Renshaw, P.F. Lithium and Valproic Acid Treatment Effects on Brain Chemistry in Bipolar Disorder. Biol. Psychiatry 2004, 56, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.A. Mechanisms of Action of Antiepileptic Drugs. Seizure 1995, 4, 267–271. [Google Scholar] [CrossRef] [Green Version]
- Al-Griw, M.A.; Alshibani, Z.O.; Alghazeer, R.; Elhensheri, M.; Tabagh, R.M.; Eskandrani, A.A.; Alansari, W.S.; Habibulla, M.M.; Shamlan, G. Histone Deacetylase 2 Inhibitor Valproic Acid Attenuates Bisphenol A-Induced Liver Pathology in Male Mice. Sci. Rep. 2022, 12, 10258. [Google Scholar] [CrossRef]
- Brotzmann, K.; Wolterbeek, A.; Kroese, D.; Braunbeck, T. Neurotoxic Effects in Zebrafish Embryos by Valproic Acid and Nine of Its Analogues: The Fish-Mouse Connection? Arch. Toxicol. 2021, 95, 641–657. [Google Scholar] [CrossRef]
- Delage, C.; Palayer, M.; Etain, B.; Hagenimana, M.; Blaise, N.; Smati, J.; Chouchana, M.; Bloch, V.; Besson, V.C. Valproate, Divalproex, Valpromide: Are the Differences in Indications Justified? Biomed. Pharmacother. 2023, 158, 114051. [Google Scholar] [CrossRef]
- Mendes, G.D.; Lotufo, C.C.; Bosio-Guimarães, R.A.; de Castro, H.A.; Babadopulos, T.; Ribas Freitas, A.R.; de Antunes, N.J.; Nucci, G. De Comparative Bioavailability Study with Two Sodium Valproate Tablet Formulations Administered under Fasting Conditions in Healthy Subjects. Int. J. Clin. Pharmacol. Ther. 2022, 60, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Lin, J.; Liu, X.; Li, W.; Ding, Y.; Zhang, Y.; Zhou, S.; Guo, N.; Li, Q. Characterization of the Locomotor Activities of Zebrafish Larvae under the Influence of Various Neuroactive Drugs. Ann. Transl. Med. 2018, 6, 173. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Sharma, P.; Mazumder, A.G.; Rana, A.K.; Sharma, S.; Singh, D. Development and Validation of Chemical Kindling in Adult Zebrafish: A Simple and Improved Chronic Model for Screening of Antiepileptic Agents. J. Neurosci. Methods 2020, 346, 108916. [Google Scholar] [CrossRef]
- Wallenburg, E.; Klok, B.; de Jong, K.; de Maat, M.; van Erp, N.; Stalpers-Konijnenburg, S.; Essink, G.; van Luin, M. Monitoring Protein-Unbound Valproic Acid Serum Concentrations in Clinical Practice. Ther. Drug Monit. 2017, 39, 269–272. [Google Scholar] [CrossRef]
- Gebuijs, I.G.E.; Metz, J.R.; Zethof, J.; Carels, C.E.L.; Wagener, F.A.D.T.G.; Von den Hoff, J.W. The Anti-Epileptic Drug Valproic Acid Causes Malformations in the Developing Craniofacial Skeleton of Zebrafish Larvae. Mech. Dev. 2020, 163, 103632. [Google Scholar] [CrossRef]
- Wang, J.; Ma, Y.; Li, W.; Hu, F.; Chen, T.; Shen, X.; Feng, S. Study on Pharmacokinetics and Tissue Distribution of Pteryxin in Mice by Ultra-Pressure Liquid Chromatography with Tandem Mass Spectrometry. Biomed. Chromatogr. 2012, 26, 802–807. [Google Scholar] [CrossRef]
- Jeong, J.Y.; Kwon, H.B.; Ahn, J.C.; Kang, D.; Kwon, S.H.; Park, J.A.; Kim, K.W. Functional and Developmental Analysis of the Blood-Brain Barrier in Zebrafish. Brain Res. Bull. 2008, 75, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Fleming, A.; Diekmann, H.; Goldsmith, P. Functional Characterisation of the Maturation of the Blood-Brain Barrier in Larval Zebrafish. PLoS ONE 2013, 8, e77548. [Google Scholar] [CrossRef] [Green Version]
- Dalangin, R.; Kim, A.; Campbell, R.E. The Role of Amino Acids in Neurotransmission and Fluorescent Tools for Their Detection. Int. J. Mol. Sci. 2020, 21, 6197. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Copmans, D.; Partoens, M.; Hunyadi, B.; Luyten, W.; De Witte, P. Zebrafish-Based Screening of Antiseizure Plants Used in Traditional Chinese Medicine: Magnolia officinalis Extract and Its Constituents Magnolol and Honokiol Exhibit Potent Anticonvulsant Activity in a Therapy-Resistant Epilepsy Model. ACS Chem Neurosci. 2020, 11, 730. [Google Scholar] [CrossRef] [PubMed]
- Gruenbaum, S.E.; Chen, E.C.; Sandhu, M.R.S.; Deshpande, K.; Dhaher, R.; Hersey, D.; Eid, T. Branched-Chain Amino Acids and Seizures: A Systematic Review of the Literature. CNS Drugs 2019, 33, 755–770. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Liu, C.; Chen, X.; Zheng, L.; Zou, Y.; Wen, H.; Guan, P.; Lu, F.; Luo, Y.; Tan, G.; et al. Exosomes-Loaded Electroconductive Hydrogel Synergistically Promotes Tissue Repair after Spinal Cord Injury via Immunoregulation and Enhancement of Myelinated Axon Growth. Adv. Sci. 2022, 9, e2105586. [Google Scholar] [CrossRef]
- Gao, D.; Ma, L.; Xie, Y.; Xiao, B.; Xue, S.; Xiao, W.; Zhou, Y.; Cai, X.; Yang, X. Electroacupuncture Promotes Autophagy by Regulating the AKT/MTOR Signaling Pathway in Temporal Lobe Epilepsy. Neurochem. Res. 2022, 47, 2396–2404. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, E.; Nikbakht, F.; Barati, M.; Roghani, M.; Vazifekhah, S.; Khanizadeh, A.M.; Heidari, Z. Protective Effect of N-Acetyl Cysteine on the Mitochondrial Dynamic Imbalance in Temporal Lobe Epilepsy: Possible Role of MTOR. Neuropeptides 2022, 96, 102294. [Google Scholar] [CrossRef]
- Ma, C.; Teng, L.; Lin, G.; Guo, B.; Zhuo, R.; Qian, X.; Guan, T.; Wu, R.; Liu, Y.; Liu, M. L-Leucine Promotes Axonal Outgrowth and Regeneration via MTOR Activation. FASEB J. 2021, 35, e21526. [Google Scholar] [CrossRef] [PubMed]
- Ellingsen, S.; Narawane, S.; Fjose, A.; Verri, T.; Rønnestad, I. The Zebrafish Cationic Amino Acid Transporter/Glycoprotein-Associated Family: Sequence and Spatiotemporal Distribution during Development of the Transport System b(0,+) (Slc3a1/Slc7a9). Fish Physiol. Biochem. 2021, 47, 1507–1525. [Google Scholar] [CrossRef] [PubMed]
- Broide, R.S.; Salas, R.; Ji, D.; Paylor, R.; Patrick, J.W.; Dani, J.A.; De Biasi, M. Increased Sensitivity to Nicotine-Induced Seizures in Mice Expressing the L250T Alpha 7 Nicotinic Acetylcholine Receptor Mutation. Mol. Pharmacol. 2002, 61, 695–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krahl, S.E. Vagus Nerve Stimulation for Epilepsy: A Review of the Peripheral Mechanisms. Surg. Neurol. Int. 2012, 3, S47–S52. [Google Scholar] [CrossRef]
- Afra, P.; Adamolekun, B.; Aydemir, S.; Watson, G.D.R. Evolution of the Vagus Nerve Stimulation (VNS) Therapy System Technology for Drug-Resistant Epilepsy. Front. Med. Technol. 2021, 3, 696543. [Google Scholar] [CrossRef]
- Manta, S.; El Mansari, M.; Blier, P. Novel Attempts to Optimize Vagus Nerve Stimulation Parameters on Serotonin Neuronal Firing Activity in the Rat Brain. Brain Stimul. 2012, 5, 422–429. [Google Scholar] [CrossRef]
- Sourbron, J.; Lagae, L. Serotonin Receptors in Epilepsy: Novel Treatment Targets? Epilepsia Open 2022, 7, 231–246. [Google Scholar] [CrossRef]
- Sourbron, J.; Schneider, H.; Kecskés, A.; Liu, Y.; Buening, E.M.; Lagae, L.; Smolders, I.; De Witte, P. Serotonergic Modulation as Effective Treatment for Dravet Syndrome in a Zebrafish Mutant Model. ACS Chem. Neurosci. 2016, 7, 588–598. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Kecskés, A.; Copmans, D.; Langlois, M.; Crawford, A.D.; Ceulemans, B.; Lagae, L.; de Witte, P.A.M.; Esguerra, C. V Pharmacological Characterization of an Antisense Knockdown Zebrafish Model of Dravet Syndrome: Inhibition of Epileptic Seizures by the Serotonin Agonist Fenfluramine. PLoS ONE 2015, 10, e0125898. [Google Scholar] [CrossRef] [Green Version]
- Cullingford, T. The Vagus Nerve–A Common Route for Epilepsy Therapies? Lancet. Neurol. 2004, 3, 518. [Google Scholar] [CrossRef]
- Pineda, R.; Beattie, C.E.; Hall, C.W. Closed-Loop Neural Stimulation for Pentylenetetrazole-Induced Seizures in Zebrafish. Dis. Model. Mech. 2013, 6, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Jackstadt, M.M.; Chamberlain, C.A.; Doonan, S.R.; Shriver, L.P.; Patti, G.J. A Multidimensional Metabolomics Workflow to Image Biodistribution and Evaluate Pharmacodynamics in Adult Zebrafish. Dis. Model. Mech. 2022, 15, dmm049550. [Google Scholar] [CrossRef] [PubMed]
- Olsson, C.; Holmberg, A.; Holmgren, S. Development of Enteric and Vagal Innervation of the Zebrafish (Danio Rerio) Gut. J. Comp. Neurol. 2008, 508, 756–770. [Google Scholar] [CrossRef] [PubMed]
- Kuil, L.E.; Chauhan, R.K.; Cheng, W.W.; Hofstra, R.M.W.; Alves, M.M. Zebrafish: A Model Organism for Studying Enteric Nervous System Development and Disease. Front. Cell Dev. Biol. 2021, 8, 629073. [Google Scholar] [CrossRef] [PubMed]
- Yabut, J.M.; Crane, J.D.; Green, A.E.; Keating, D.J.; Khan, W.I.; Steinberg, G.R. Emerging Roles for Serotonin in Regulating Metabolism: New Implications for an Ancient Molecule. Endocr. Rev. 2019, 40, 1092–1107. [Google Scholar] [CrossRef]
- Yan, R.; Li, Y.; Müller, J.; Zhang, Y.; Singer, S.; Xia, L.; Zhong, X.; Gertsch, J.; Altmann, K.H.; Zhou, Q. Mechanism of substrate transport and inhibition of the human LAT1-4F2hc amino acid transporter. Cell Discov. 2021, 7, 16. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment/Concentration/Time | [ng/g] | [µM] | % Absorbed |
|---|---|---|---|
| Pteryxin/1 µM/1 h | 468.7 ± 44.1 | 1.2 ± 0.1 | 1.2% |
| Pteryxin/10 µM/1 h | 1243.3 ± 201.3 | 3.2 ± 0.5 | 32.2% |
| Pteryxin/20 µM/5 min | 820.0 ± 46.3 | 2.1 ± 0.1 | 10.1% |
| Pteryxin/20 µM/30 min | 2530.0 ± 105.4 | 6.5 ± 0.3 | 32.7% |
| Pteryxin/20 µM/1 h | 4280.0 ± 284.8 | 11.1 ± 0.7 | 55.4% |
| Pteryxin/20 μM/18 h | 3406.7 ± 213.6 | 8.8 ± 0.5 | 44.0% |
| Pteryxin/20 μM/without larvae | 4090.6 ± 381.6 | 20.5 ± 0.1 | - |
| VPN/VPA 100 µM 1 h | 940.7 ± 40.8 | 6.5 ± 0.3 | 6.5% |
| VPN/VPA 500 µM 1 h | 1680.0 ± 30.0 | 11.6 ± 0.2 | 2.3% |
| VPN/VPA 1 mM 1 h | 4740.0 ± 111.4 | 32.9 ± 0.8 | 3.3% |
| VPN/VPA 5 mM 5 min | 5596.6 ± 289.9 | 38.8 ± 2.0 | 0.8% |
| VPN/VPA 5 mM 30 min | 10,366.7 ± 472.6 | 71.9 ± 3.3 | 1.4% |
| VPN/VPA 5 mM 1 h | 18,166.7 ± 404.1 | 126.0 ± 2.8 | 2.5% |
| VPN/VPA 10 mM 1 h | 33,933.3 ± 1457.2 | 235.3 ± 10.1 | 9.4% |
| VPN/VPA (5 mM) 18 h | 5473.33 ± 225.02 | 37.97 ± 1.57 | 0.76% (metabolism) |
| VPN/VPA 5 mM without larvae | 734,333 ± 5859 | 5092 ± 40.6 | 100% |
| Medium control | No detected | Not detected | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skiba, A.; Pellegata, D.; Morozova, V.; Kozioł, E.; Budzyńska, B.; Lee, S.M.-Y.; Gertsch, J.; Skalicka-Woźniak, K. Pharmacometabolic Effects of Pteryxin and Valproate on Pentylenetetrazole-Induced Seizures in Zebrafish Reveal Vagus Nerve Stimulation. Cells 2023, 12, 1540. https://doi.org/10.3390/cells12111540
Skiba A, Pellegata D, Morozova V, Kozioł E, Budzyńska B, Lee SM-Y, Gertsch J, Skalicka-Woźniak K. Pharmacometabolic Effects of Pteryxin and Valproate on Pentylenetetrazole-Induced Seizures in Zebrafish Reveal Vagus Nerve Stimulation. Cells. 2023; 12(11):1540. https://doi.org/10.3390/cells12111540
Chicago/Turabian StyleSkiba, Adrianna, Daniele Pellegata, Veronika Morozova, Ewelina Kozioł, Barbara Budzyńska, Simon Ming-Yuen Lee, Jürg Gertsch, and Krystyna Skalicka-Woźniak. 2023. "Pharmacometabolic Effects of Pteryxin and Valproate on Pentylenetetrazole-Induced Seizures in Zebrafish Reveal Vagus Nerve Stimulation" Cells 12, no. 11: 1540. https://doi.org/10.3390/cells12111540