
In Vitro Effects of Methylprednisolone over Oligodendroglial Cells: Foresight to Future Cell Therapies

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. HOG Cell Culture
2.2. Analysis of HOG Cell Proliferation and Maturation
2.3. Western Blot
2.4. Statistical Analysis
3. Results
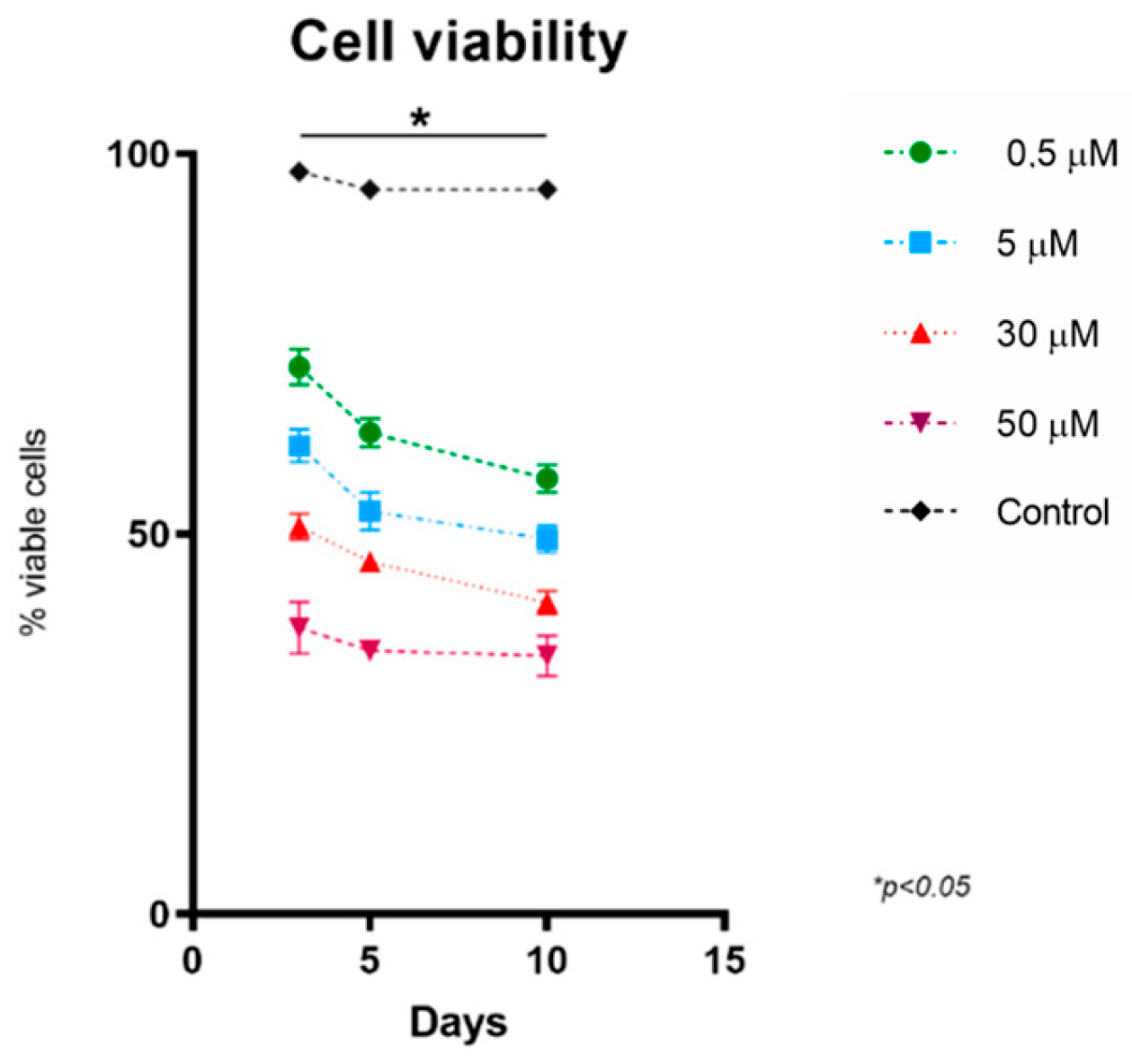
3.1. Methylprednisolone Reduces the Proliferation of HOG Cells by Reducing Their Cell Viability in a Dose-Dependent Manner
3.2. Methylprednisolone Induces Oligodendrocyte Cell Death in a Dose-Dependent Manner
3.3. Methylprednisolone Reduces Oligodendrocyte Differentiation in a Dose-Dependent Manner
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Czepiel, M.; Balasubramaniyan, V.; Schaafsma, W.; Stancic, M.; Mikkers, H.; Huisman, C.; Boddeke, E.; Copray, S. Differentiation of induced pluripotent stem cells into functional oligodendrocytes. Glia 2011, 59, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B.; Lee, H.; Arauzo-Bravo, M.J.; Hwang, K.; Nam, D.; Park, M.R.; Zaehres, H.; Park, K.I.; Lee, S.J. Oct4-induced oligodendrocyte progenitor cells enhance functional recovery in spinal cord injury model. EMBO J. 2015, 34, 2971–2983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazm Bojnordi, M.; Ghasemi, H.H.; Akbari, E. Remyelination after Lysophosphatidyl Choline-Induced Demyelination Is Stimulated by Bone Marrow Stromal Cell-Derived Oligoprogenitor Cell Transplantation. Cells Tissues Organs 2015, 200, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Sim, F.J.; McClain, C.R.; Schanz, S.J.; Protack, T.L.; Windrem, M.S.; Goldman, S.A. CD140a identifies a population of highly myelinogenic, migration-competent and efficiently engrafting human oligodendrocyte progenitor cells. Nat. Biotechnol. 2011, 29, 934–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Bates, J.; Li, X.; Schanz, S.; Chandler-Militello, D.; Levine, C.; Maherali, N.; Studer, L.; Hochedlinger, K.; Windrem, M.; et al. Human iPSC-derived oligodendrocyte progenitor cells can myelinate and rescue a mouse model of congenital hypomyelination. Cell Stem Cell 2013, 12, 252–264. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.; Zuchero, J.B.; Ahlenius, H.; Marro, S.; Ng, Y.H.; Vierbuchen, T.; Hawkins, J.S.; Geissler, R.; Barres, B.A.; Wernig, M. Generation of oligodendroglial cells by direct lineage conversion. Nat. Biotechnol. 2013, 31, 434–439. [Google Scholar] [CrossRef] [Green Version]
- Zawadzka, M.; Rivers, L.E.; Fancy, S.P.; Zhao, C.; Tripathi, R.; Jamen, F.; Young, K.; Goncharevich, A.; Pohl, H.; Rizzi, M.; et al. CNS-resident glial progenitor/stem cells produce Schwann cells as well as oligodendrocytes during repair of CNS demyelination. Cell Stem Cell 2010, 6, 578–590. [Google Scholar] [CrossRef] [Green Version]
- Matias-Guiu, J.; Martinez-Vazquez, J.; Ruibal, A.; Colomer, R.; Boada, M.; Codina, A. Myelin basic protein and creatine kinase BB isoenzyme as CSF markers of intracranial tumors and stroke. Acta Neurol. Scand. 1986, 73, 461–465. [Google Scholar] [CrossRef]
- Itoh, K.; Maki, T.; Lok, J.; Arai, K. Mechanisms of cell-cell interaction in oligodendrogenesis and remyelination after stroke. Brain Res. 2015, 1623, 135–149. [Google Scholar] [CrossRef] [Green Version]
- Maki, T.; Liang, A.C.; Miyamoto, N.; Lo, E.H.; Arai, K. Mechanisms of oligodendrocyte regeneration from ventricular-subventricular zone-derived progenitor cells in white matter diseases. Front. Cell. Neurosci. 2013, 7, 275. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Chopp, M.; Zhang, Z.G. Oligodendrogenesis after cerebral ischemia. Front. Cell. Neurosci. 2013, 7, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, E.G.; Kang, S.H.; Fukaya, M.; Bergles, D.E. Oligodendrocyte progenitors balance growth with self-repulsion to achieve homeostasis in the adult brain. Nat. Neurosci. 2013, 16, 668–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, R.J.; ffrench-Constant, C.; Edgar, J.M.; Smith, K.J. Neuroprotection and repair in multiple sclerosis. Nat. Rev. Neurol. 2012, 8, 624–634. [Google Scholar] [CrossRef] [PubMed]
- All, A.H.; Gharibani, P.; Gupta, S.; Bazley, F.A.; Pashai, N.; Chou, B.K.; Shah, S.; Resar, L.M.; Cheng, L.; Gearhart, J.D.; et al. Early intervention for spinal cord injury with human induced pluripotent stem cells oligodendrocyte progenitors. PLoS ONE 2015, 10, e0116933. [Google Scholar] [CrossRef] [Green Version]
- Egawa, N.; Takase, H.; Josephine, L.; Takahashi, R.; Arai, K. Clinical application of oligodendrocyte precursor cells for cell-based therapy. Brain Circ. 2016, 2, 121–125. [Google Scholar] [CrossRef]
- Karimi-Abdolrezaee, S.; Eftekharpour, E.; Wang, J.; Morshead, C.M.; Fehlings, M.G. Delayed transplantation of adult neural precursor cells promotes remyelination and functional neurological recovery after spinal cord injury. J. Neurosci. 2006, 26, 3377–3389. [Google Scholar] [CrossRef] [Green Version]
- Kawabata, S.; Takano, M.; Numasawa-Kuroiwa, Y.; Itakura, G.; Kobayashi, Y.; Nishiyama, Y.; Sugai, K.; Nishimura, S.; Iwai, H.; Isoda, M.; et al. Grafted Human iPS Cell-Derived Oligodendrocyte Precursor Cells Contribute to Robust Remyelination of Demyelinated Axons after Spinal Cord Injury. Stem Cell Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Manley, N.C.; Priest, C.A.; Denham, J.; Wirth, E.D., 3rd; Lebkowski, J.S. Human Embryonic Stem Cell-Derived Oligodendrocyte Progenitor Cells: Preclinical Efficacy and Safety in Cervical Spinal Cord Injury. Stem Cells Transl. Med. 2017, 6, 1917–1929. [Google Scholar] [CrossRef]
- Priest, C.A.; Manley, N.C.; Denham, J.; Wirth, E.D., 3rd; Lebkowski, J.S. Preclinical safety of human embryonic stem cell-derived oligodendrocyte progenitors supporting clinical trials in spinal cord injury. Regen. Med. 2015, 10, 939–958. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Muguruza, E.; Villar-Gomez, N.; Matias-Guiu, J.A.; Selma-Calvo, B.; Moreno-Jimenez, L.; Sancho-Bielsa, F.; Lopez-Carbonero, J.; Benito-Martin, M.S.; Garcia-Flores, S.; Bonel-Garcia, N.; et al. The Integration of Cell Therapy and Biomaterials as Treatment Strategies for Remyelination. Life 2022, 12, 474. [Google Scholar] [CrossRef]
- Shen, S.; Sandoval, J.; Swiss, V.A.; Li, J.; Dupree, J.; Franklin, R.J.; Casaccia-Bonnefil, P. Age-dependent epigenetic control of differentiation inhibitors is critical for remyelination efficiency. Nat. Neurosci. 2008, 11, 1024–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, F.J.; Zhao, C.; Penderis, J.; Franklin, R.J. The age-related decrease in CNS remyelination efficiency is attributable to an impairment of both oligodendrocyte progenitor recruitment and differentiation. J. Neurosci. 2002, 22, 2451–2459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buntinx, M.; Vanderlocht, J.; Hellings, N.; Vandenabeele, F.; Lambrichts, I.; Raus, J.; Ameloot, M.; Stinissen, P.; Steels, P. Characterization of three human oligodendroglial cell lines as a model to study oligodendrocyte injury: Morphology and oligodendrocyte-specific gene expression. J. Neurocytol. 2003, 32, 25–38. [Google Scholar] [CrossRef]
- Dawson, G.; Dawson, S.A.; Post, G.R. Regulation of phospholipase D activity in a human oligodendroglioma cell line (HOG). J. Neurosci. Res. 1993, 34, 324–330. [Google Scholar] [CrossRef]
- de Arriba Zerpa, G.A.; Saleh, M.C.; Fernandez, P.M.; Guillou, F.; Espinosa de los Monteros, A.; de Vellis, J.; Zakin, M.M.; Baron, B. Alternative splicing prevents transferrin secretion during differentiation of a human oligodendrocyte cell line. J. Neurosci. Res. 2000, 61, 388–395. [Google Scholar] [CrossRef] [PubMed]
- De Kleijn, K.M.A.; Zuure, W.A.; Peijnenborg, J.; Heuvelmans, J.M.; Martens, G.J.M. Reappraisal of Human HOG and MO3.13 Cell Lines as a Model to Study Oligodendrocyte Functioning. Cells 2019, 8, 1096. [Google Scholar] [CrossRef] [Green Version]
- Post, G.R.; Dawson, G. Characterization of a cell line derived from a human oligodendroglioma. Mol. Chem. Neuropathol. 1992, 16, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Pinilla, E.; Rubio-Sardon, N.; Villar-Conde, S.; Navarro, G.; Del Valle, E.; Tolivia, J.; Franco, R.; Navarro, A. Cuprizone-Induced Neurotoxicity in Human Neural Cell Lines Is Mediated by a Reversible Mitochondrial Dysfunction: Relevance for Demyelination Models. Brain Sci. 2021, 11, 272. [Google Scholar] [CrossRef]
- Antonow-Schlorke, I.; Helgert, A.; Gey, C.; Coksaygan, T.; Schubert, H.; Nathanielsz, P.W.; Witte, O.W.; Schwab, M. Adverse effects of antenatal glucocorticoids on cerebral myelination in sheep. Obstet. Gynecol. 2009, 113, 142–151. [Google Scholar] [CrossRef]
- Huang, W.L.; Harper, C.G.; Evans, S.F.; Newnham, J.P.; Dunlop, S.A. Repeated prenatal corticosteroid administration delays myelination of the corpus callosum in fetal sheep. Int. J. Dev. Neurosci. 2001, 19, 415–425. [Google Scholar] [CrossRef]
- Quinlivan, J.A.; Archer, M.A.; Evans, S.F.; Newnham, J.P.; Dunlop, S.A. Fetal sciatic nerve growth is delayed following repeated maternal injections of corticosteroid in sheep. J. Perinat. Med. 2000, 28, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Zalewska, K.; Hood, R.J.; Pietrogrande, G.; Sanchez-Bezanilla, S.; Ong, L.K.; Johnson, S.J.; Young, K.M.; Nilsson, M.; Walker, F.R. Corticosterone Administration Alters White Matter Tract Structure and Reduces Gliosis in the Sub-Acute Phase of Experimental Stroke. Int. J. Mol. Sci. 2021, 22, 6693. [Google Scholar] [CrossRef] [PubMed]
- Bartholdi, D.; Schwab, M.E. Methylprednisolone inhibits early inflammatory processes but not ischemic cell death after experimental spinal cord lesion in the rat. Brain Res. 1995, 672, 177–186. [Google Scholar] [CrossRef]
- Keegan, B.M.; Noseworthy, J.H. Multiple sclerosis. Annu. Rev. Med. 2002, 53, 285–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lattanzi, S.; Cagnetti, C.; Danni, M.; Provinciali, L.; Silvestrini, M. Oral and intravenous steroids for multiple sclerosis relapse: A systematic review and meta-analysis. J. Neurol. 2017, 264, 1697–1704. [Google Scholar] [CrossRef]
- Repovic, P. Management of Multiple Sclerosis Relapses. Continuum 2019, 25, 655–669. [Google Scholar] [CrossRef]
- Smets, I.; Van Deun, L.; Bohyn, C.; van Pesch, V.; Vanopdenbosch, L.; Dive, D.; Bissay, V.; Dubois, B.; Belgian Study Group for Multiple Sclerosis. Corticosteroids in the management of acute multiple sclerosis exacerbations. Acta Neurol. Belg. 2017, 117, 623–633. [Google Scholar] [CrossRef]
- Pavelko, K.D.; van Engelen, B.G.; Rodriguez, M. Acceleration in the rate of CNS remyelination in lysolecithin-induced demyelination. J. Neurosci. 1998, 18, 2498–2505. [Google Scholar] [CrossRef] [Green Version]
- Triarhou, L.C.; Herndon, R.M. The effect of dexamethasone on L-alpha-lysophosphatidyl choline (lysolecithin)-induced demyelination of the rat spinal cord. Arch. Neurol. 1986, 43, 121–125. [Google Scholar] [CrossRef]
- Torre-Fuentes, L.; Moreno-Jimenez, L.; Pytel, V.; Matias-Guiu, J.A.; Gomez-Pinedo, U.; Matias-Guiu, J. Experimental models of demyelination and remyelination. Neurologia (Engl. Ed.) 2020, 35, 32–39. [Google Scholar] [CrossRef]
- Gomez-Pinedo, U.; Cuevas, J.A.; Benito-Martin, M.S.; Moreno-Jimenez, L.; Esteban-Garcia, N.; Torre-Fuentes, L.; Matias-Guiu, J.A.; Pytel, V.; Montero, P.; Matias-Guiu, J. Vitamin D increases remyelination by promoting oligodendrocyte lineage differentiation. Brain Behav. 2020, 10, e01498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chari, D.M.; Zhao, C.; Kotter, M.R.; Blakemore, W.F.; Franklin, R.J. Corticosteroids delay remyelination of experimental demyelination in the rodent central nervous system. J. Neurosci. Res. 2006, 83, 594–605. [Google Scholar] [CrossRef] [PubMed]
- Alonso, G. Prolonged corticosterone treatment of adult rats inhibits the proliferation of oligodendrocyte progenitors present throughout white and gray matter regions of the brain. Glia 2000, 31, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Clarner, T.; Parabucki, A.; Beyer, C.; Kipp, M. Corticosteroids impair remyelination in the corpus callosum of cuprizone-treated mice. J. Neuroendocrinol. 2011, 23, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Halfpenny, C.A.; Scolding, N.J. Immune-modifying agents do not impair the survival, migration or proliferation of oligodendrocyte progenitors (CG-4) in vitro. J. Neuroimmunol. 2003, 139, 9–16. [Google Scholar] [CrossRef]
- Chesik, D.; De Keyser, J. Progesterone and dexamethasone differentially regulate the IGF-system in glial cells. Neurosci. Lett. 2010, 468, 178–182. [Google Scholar] [CrossRef]
- Gomez-Pinedo, U.; Matias-Guiu, J.A.; Benito-Martin, M.S.; Moreno-Jimenez, L.; Sanclemente-Alaman, I.; Selma-Calvo, B.; Perez-Suarez, S.; Sancho-Bielsa, F.; Canales-Aguirre, A.; Mateos-Diaz, J.C.; et al. Intranasal Administration of Undifferentiated Oligodendrocyte Lineage Cells as a Potential Approach to Deliver Oligodendrocyte Precursor Cells into Brain. Int. J. Mol. Sci. 2021, 22, 10738. [Google Scholar] [CrossRef]
- Bello-Morales, R.; Crespillo, A.J.; Garcia, B.; Dorado, L.A.; Martin, B.; Tabares, E.; Krummenacher, C.; de Castro, F.; Lopez-Guerrero, J.A. The effect of cellular differentiation on HSV-1 infection of oligodendrocytic cells. PLoS ONE 2014, 9, e89141. [Google Scholar] [CrossRef] [Green Version]
- Dubois-Dalcq, M.; Behar, T.; Hudson, L.; Lazzarini, R.A. Emergence of three myelin proteins in oligodendrocytes cultured without neurons. J. Cell Biol. 1986, 102, 384–392. [Google Scholar] [CrossRef] [Green Version]
- Post, G.R.; Dawson, G. Regulation of carbachol- and histamine-induced inositol phospholipid hydrolysis in a human oligodendroglioma. Glia 1992, 5, 122–130. [Google Scholar] [CrossRef]
- Qin, J.; Goswami, R.; Dawson, S.; Dawson, G. Expression of the receptor for advanced glycation end products in oligodendrocytes in response to oxidative stress. J. Neurosci. Res. 2008, 86, 2414–2422. [Google Scholar] [CrossRef] [Green Version]
- Testai, F.D.; Landek, M.A.; Goswami, R.; Ahmed, M.; Dawson, G. Acid sphingomyelinase and inhibition by phosphate ion: Role of inhibition by phosphatidyl-myo-inositol 3,4,5-triphosphate in oligodendrocyte cell signaling. J. Neurochem. 2004, 89, 636–644. [Google Scholar] [CrossRef]
- Buntinx, M.; Gielen, E.; Van Hummelen, P.; Raus, J.; Ameloot, M.; Steels, P.; Stinissen, P. Cytokine-induced cell death in human oligodendroglial cell lines. II: Alterations in gene expression induced by interferon-gamma and tumor necrosis factor-alpha. J. Neurosci. Res. 2004, 76, 846–861. [Google Scholar] [CrossRef] [PubMed]
- Buntinx, M.; Moreels, M.; Vandenabeele, F.; Lambrichts, I.; Raus, J.; Steels, P.; Stinissen, P.; Ameloot, M. Cytokine-induced cell death in human oligodendroglial cell lines: I. Synergistic effects of IFN-gamma and TNF-alpha on apoptosis. J. Neurosci. Res. 2004, 76, 834–845. [Google Scholar] [CrossRef] [PubMed]
- Starost, L.; Lindner, M.; Herold, M.; Xu, Y.K.T.; Drexler, H.C.A.; Hess, K.; Ehrlich, M.; Ottoboni, L.; Ruffini, F.; Stehling, M.; et al. Extrinsic immune cell-derived, but not intrinsic oligodendroglial factors contribute to oligodendroglial differentiation block in multiple sclerosis. Acta Neuropathol. 2020, 140, 715–736. [Google Scholar] [CrossRef] [PubMed]
- Diem, R.; Hobom, M.; Maier, K.; Weissert, R.; Storch, M.K.; Meyer, R.; Bähr, M. Methylprednisolone increases neuronal apoptosis during autoimmune CNS inflammation by inhibition of an endogenous neuroprotective pathway. J. Neurosci. 2003, 23, 6993–7000. [Google Scholar] [CrossRef] [PubMed]
- Arnett, H.A.; Wang, Y.; Matsushima, G.K.; Suzuki, K.; Ting, J.P. Functional genomic analysis of remyelination reveals importance of inflammation in oligodendrocyte regeneration. J. Neurosci. 2003, 23, 9824–9832. [Google Scholar] [CrossRef] [Green Version]
- Doring, A.; Sloka, S.; Lau, L.; Mishra, M.; van Minnen, J.; Zhang, X.; Kinniburgh, D.; Rivest, S.; Yong, V.W. Stimulation of monocytes, macrophages, and microglia by amphotericin B and macrophage colony-stimulating factor promotes remyelination. J. Neurosci. 2015, 35, 1136–1148. [Google Scholar] [CrossRef] [Green Version]
- Foote, A.K.; Blakemore, W.F. Inflammation stimulates remyelination in areas of chronic demyelination. Brain 2005, 128, 528–539. [Google Scholar] [CrossRef] [Green Version]
- Miron, V.E.; Franklin, R.J. Macrophages and CNS remyelination. J. Neurochem. 2014, 130, 165–171. [Google Scholar] [CrossRef]
- Moore, C.S.; Cui, Q.L.; Warsi, N.M.; Durafourt, B.A.; Zorko, N.; Owen, D.R.; Antel, J.P.; Bar-Or, A. Direct and indirect effects of immune and central nervous system-resident cells on human oligodendrocyte progenitor cell differentiation. J. Immunol. 2015, 194, 761–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, L.; He, C. Polarization of macrophages and microglia in inflammatory demyelination. Neurosci. Bull. 2013, 29, 189–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, R.; Fernandez-Suarez, D. Alternatively activated microglia and macrophages in the central nervous system. Prog. Neurobiol. 2015, 131, 65–86. [Google Scholar] [CrossRef]
- Heppner, F.L.; Greter, M.; Marino, D.; Falsig, J.; Raivich, G.; Hovelmeyer, N.; Waisman, A.; Rulicke, T.; Prinz, M.; Priller, J.; et al. Experimental autoimmune encephalomyelitis repressed by microglial paralysis. Nat. Med. 2005, 11, 146–152. [Google Scholar] [CrossRef]
- Molina-Gonzalez, I.; Miron, V.E. Astrocytes in myelination and remyelination. Neurosci. Lett. 2019, 713, 134532. [Google Scholar] [CrossRef] [PubMed]
- Ponomarev, E.D.; Maresz, K.; Tan, Y.; Dittel, B.N. CNS-derived interleukin-4 is essential for the regulation of autoimmune inflammation and induces a state of alternative activation in microglial cells. J. Neurosci. 2007, 27, 10714–10721. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Sun, D.; Feng, J.; Tan, W.; Fang, X.; Zhao, M.; Zhao, X.; Pu, Y.; Huang, A.; Xiang, Z.; et al. MSX3 Switches Microglia Polarization and Protects from Inflammation-Induced Demyelination. J. Neurosci. 2015, 35, 6350–6365. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, S.I.; Pickard, M.R.; Khong, M.; Smith, H.L.; Mann, C.L.; Emes, R.D.; Chari, D.M. Identifying the cellular targets of drug action in the central nervous system following corticosteroid therapy. ACS Chem. Neurosci. 2014, 5, 51–63. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Pinedo, U.; Sirerol-Piquer, M.S.; Duran-Moreno, M.; Garcia-Verdugo, J.M.; Matias-Guiu, J. Alexander Disease Mutations Produce Cells with Coexpression of Glial Fibrillary Acidic Protein and NG2 in Neurosphere Cultures and Inhibit Differentiation into Mature Oligodendrocytes. Front. Neurol. 2017, 8, 255. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Pinedo, U.; Sirerol-Piquer, S.; Duran-Moreno, M.; Matias-Guiu, J.A.; Barcia, J.A.; Garcia-Verdugo, J.M.; Matias-Guiu, J. NG2 and GFAP co-expression after differentiation in cells transfected with mutant GFAP and in undifferentiated glioma cells. Neurologia (Engl. Ed.) 2020, 35, 479–485. [Google Scholar] [CrossRef]
- Vigano, F.; Mobius, W.; Gotz, M.; Dimou, L. Transplantation reveals regional differences in oligodendrocyte differentiation in the adult brain. Nat. Neurosci. 2013, 16, 1370–1372. [Google Scholar] [CrossRef] [PubMed]
- Spitzer, S.O.; Sitnikov, S.; Kamen, Y.; Evans, K.A.; Kronenberg-Versteeg, D.; Dietmann, S.; De Faria, O., Jr.; Agathou, S.; Karadottir, R.T. Oligodendrocyte Progenitor Cells Become Regionally Diverse and Heterogeneous with Age. Neuron 2019, 101, 459–471.e455. [Google Scholar] [CrossRef] [Green Version]
- Dimou, L.; Simons, M. Diversity of oligodendrocytes and their progenitors. Curr. Opin. Neurobiol. 2017, 47, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Tomassy, G.S.; Berger, D.R.; Chen, H.H.; Kasthuri, N.; Hayworth, K.J.; Vercelli, A.; Seung, H.S.; Lichtman, J.W.; Arlotta, P. Distinct profiles of myelin distribution along single axons of pyramidal neurons in the neocortex. Science 2014, 344, 319–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietz, K.C.; Polanco, J.J.; Pol, S.U.; Sim, F.J. Targeting human oligodendrocyte progenitors for myelin repair. Exp. Neurol. 2016, 283, 489–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsusue, Y.; Horii-Hayashi, N.; Kirita, T.; Nishi, M. Distribution of corticosteroid receptors in mature oligodendrocytes and oligodendrocyte progenitors of the adult mouse brain. J. Histochem. Cytochem. 2014, 62, 211–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matias-Guiu, J.; Matias-Guiu, J.A.; Montero-Escribano, P.; Barcia, J.A.; Canales-Aguirre, A.A.; Mateos-Diaz, J.C.; Gomez-Pinedo, U. Particles Containing Cells as a Strategy to Promote Remyelination in Patients with Multiple Sclerosis. Front. Neurol. 2020, 11, 638. [Google Scholar] [CrossRef]
- Ojeda-Hernandez, D.D.; Canales-Aguirre, A.A.; Matias-Guiu, J.; Gomez-Pinedo, U.; Mateos-Diaz, J.C. Potential of Chitosan and Its Derivatives for Biomedical Applications in the Central Nervous System. Front. Bioeng. Biotechnol. 2020, 8, 389. [Google Scholar] [CrossRef]
- Mork, S.J.; Lindegaard, K.F.; Halvorsen, T.B.; Lehmann, E.H.; Solgaard, T.; Hatlevoll, R.; Harvei, S.; Ganz, J. Oligodendroglioma: Incidence and biological behavior in a defined population. J. Neurosurg. 1985, 63, 881–889. [Google Scholar] [CrossRef]
- Van den Bent, M.J.; Reni, M.; Gatta, G.; Vecht, C. Oligodendroglioma. Crit. Rev. Oncol. Hematol. 2008, 66, 262–272. [Google Scholar] [CrossRef]
- Batchu, S.; Diaz, M.J.; Kleinberg, G.; Lucke-Wold, B. Spatial metabolic heterogeneity of oligodendrogliomas at single-cell resolution. Brain Tumor Pathol. 2023, 40, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Shi, J.; Cao, J.; Dong, B.; Guan, W. Emerging Roles and Therapeutic Interventions of Aerobic Glycolysis in Glioma. Onco. Targets Ther. 2020, 13, 6937–6955. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | MP 0.5 µM | MP 5 µM | MP 30 µM | MP 50 µM | Day | |
|---|---|---|---|---|---|---|
| Mean | 97.67 | 72 | 61.67 | 51 | 37.67 | 3 |
| SEM | 0.3333 | 2.309 | 2.186 | 1.732 | 3.383 | |
| Mean | 95.33 | 63.33 | 53 | 46.33 | 34.67 | 5 |
| SEM | 1.202 | 1.856 | 2.517 | 0.8819 | 0.8819 | |
| Mean | 95.33 | 57.33 | 49.33 | 41 | 30 | 10 |
| SEM | 0.3333 | 1.856 | 1.764 | 1.528 | 2.646 |
| Control | MP 0.5 µM | MP 5 µM | MP 30 µM | MP 50 µM | Day | |
|---|---|---|---|---|---|---|
| Mean | 2.285 | 2.172 | 2.021 | 1.125 | 0.6031 | 3 |
| SEM | 0.026 | 0.0142 | 0.099 | 0.075 | 0.0322 | |
| Mean | 2.378 | 1.466 | 0.838 | 0.408 | 0.268 | 5 |
| SEM | 0.036 | 0.061 | 0.095 | 0.014 | 0.065 | |
| Mean | 2.082 | 0.423 | 0.245 | 0.233 | 0.173 | 10 |
| SEM | 0.007 | 0.006 | 0.011 | 0.024 | 0.0211 |
| Control | MP 0.5 µM | MP 5 µM | MP 30 µM | MP 50 µM | Day | |
|---|---|---|---|---|---|---|
| Mean | 0.248 | 0.2023 | 0.141 | 0.0457 | 0.01243 | 3 |
| SEM | 0.0124 | 0.0129 | 0.00406 | 0.00523 | 0.00202 | |
| Mean | 0.287 | 0.182 | 0.054 | 0.0012 | 0.00458 | 5 |
| SEM | 0.00427 | 0.00518 | 0.0865 | 0.002 | 0.000721 | |
| Mean | 0.3432 | 0.0601 | 0.0345 | 0.00259 | 0.00172 | 10 |
| SEM | 0.00723 | 0.00246 | 0.0038 | 0.00053 | 0.0024 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Pinedo, U.; Matías-Guiu, J.A.; Ojeda-Hernandez, D.; de la Fuente-Martin, S.; Kamal, O.M.-F.; Benito-Martin, M.S.; Selma-Calvo, B.; Montero-Escribano, P.; Matías-Guiu, J. In Vitro Effects of Methylprednisolone over Oligodendroglial Cells: Foresight to Future Cell Therapies. Cells 2023, 12, 1515. https://doi.org/10.3390/cells12111515
Gómez-Pinedo U, Matías-Guiu JA, Ojeda-Hernandez D, de la Fuente-Martin S, Kamal OM-F, Benito-Martin MS, Selma-Calvo B, Montero-Escribano P, Matías-Guiu J. In Vitro Effects of Methylprednisolone over Oligodendroglial Cells: Foresight to Future Cell Therapies. Cells. 2023; 12(11):1515. https://doi.org/10.3390/cells12111515
Chicago/Turabian StyleGómez-Pinedo, Ulises, Jordi A. Matías-Guiu, Denise Ojeda-Hernandez, Sarah de la Fuente-Martin, Ola Mohamed-Fathy Kamal, Maria Soledad Benito-Martin, Belen Selma-Calvo, Paloma Montero-Escribano, and Jorge Matías-Guiu. 2023. "In Vitro Effects of Methylprednisolone over Oligodendroglial Cells: Foresight to Future Cell Therapies" Cells 12, no. 11: 1515. https://doi.org/10.3390/cells12111515