
Injury to Cone Synapses by Retinal Detachment: Differences from Rod Synapses and Protection by ROCK Inhibition

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
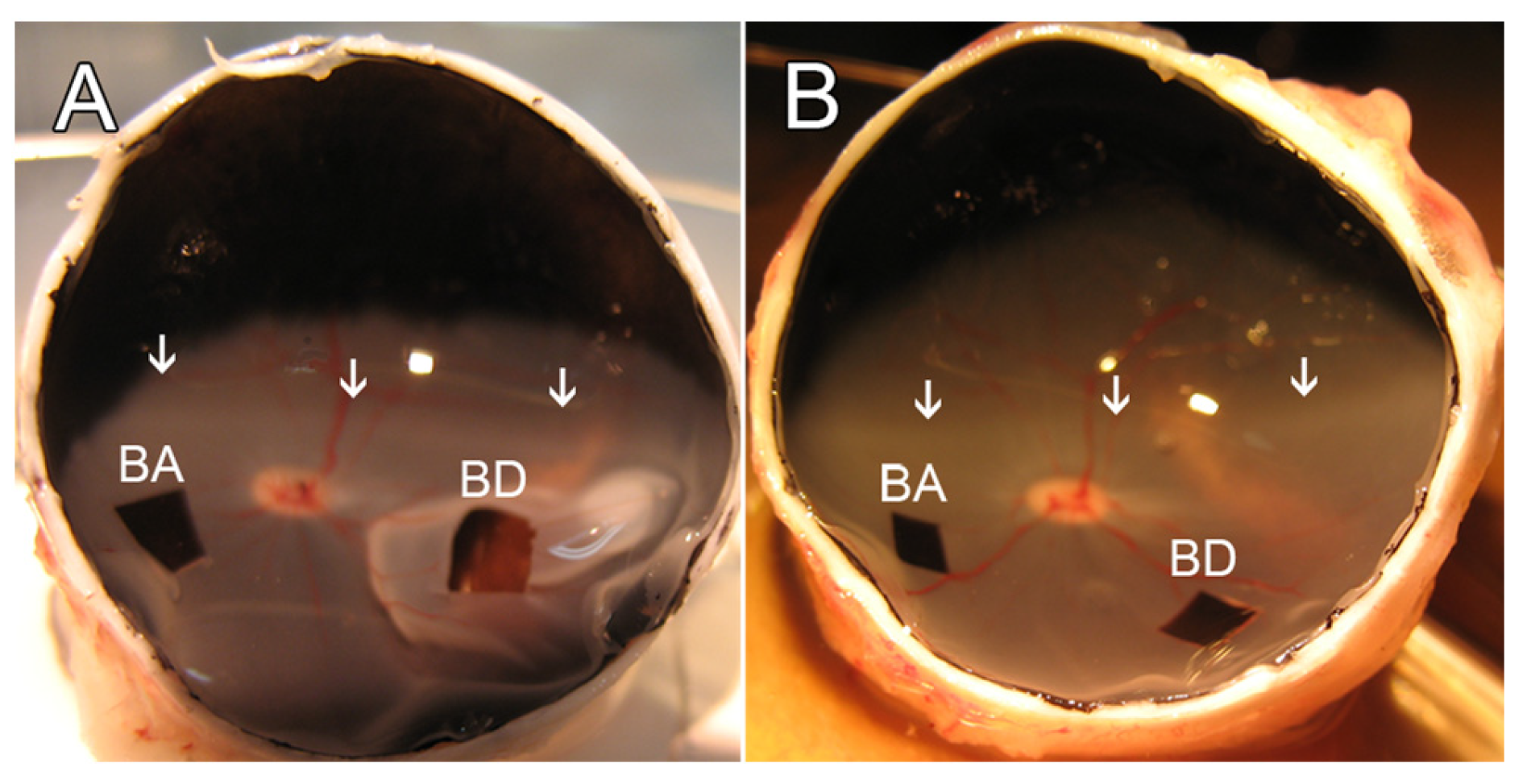
2.2. Retinal Detachment and Experimental Design
2.3. Full-Field Flash Electroretinogram (ffERG), Fundus Photography, Optical Coherence Tomography (OCT)
2.4. Sample Preparation and Immunohistochemistry
2.5. Quantification of Cone Pedicles
2.6. Quantification of Rod Synapses
2.7. Stimulated Emission Depletion (STED) Microscopy and 3D Imaging
2.8. Statistical Analysis
3. Results
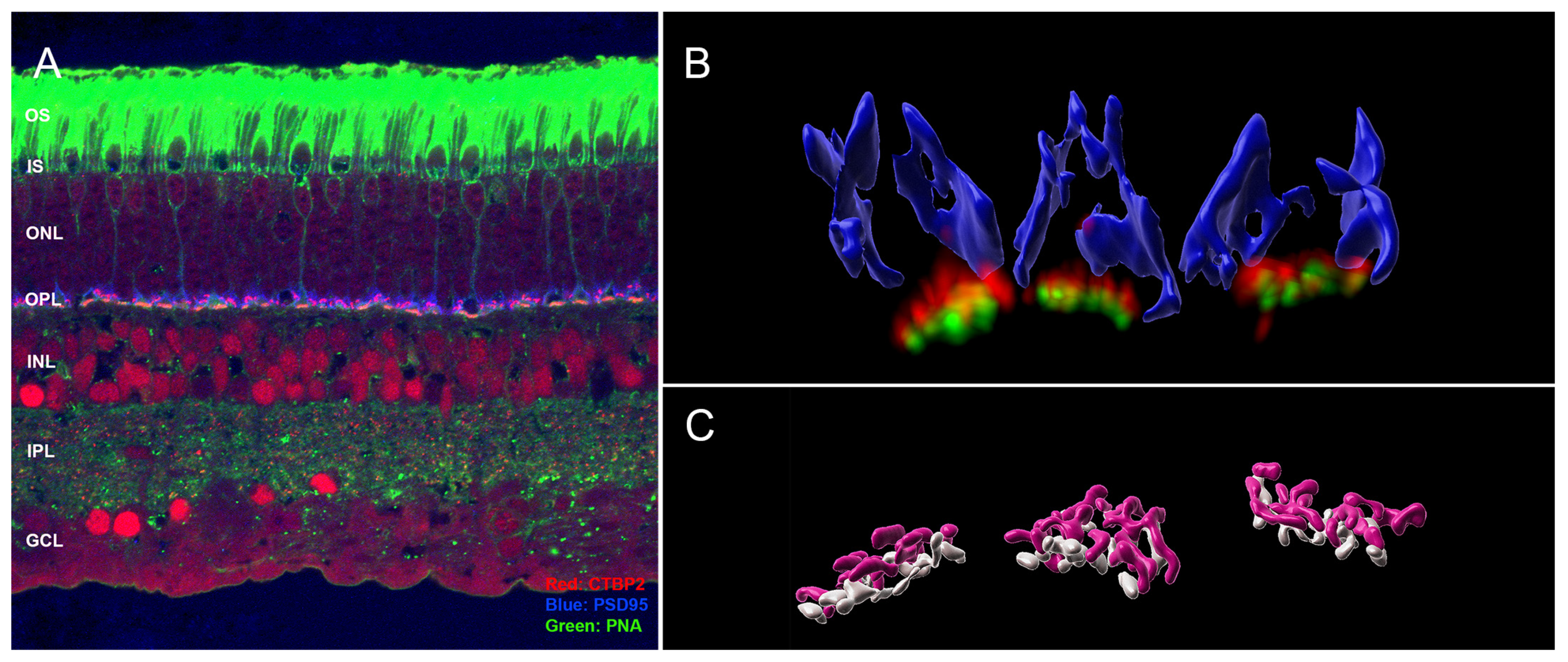
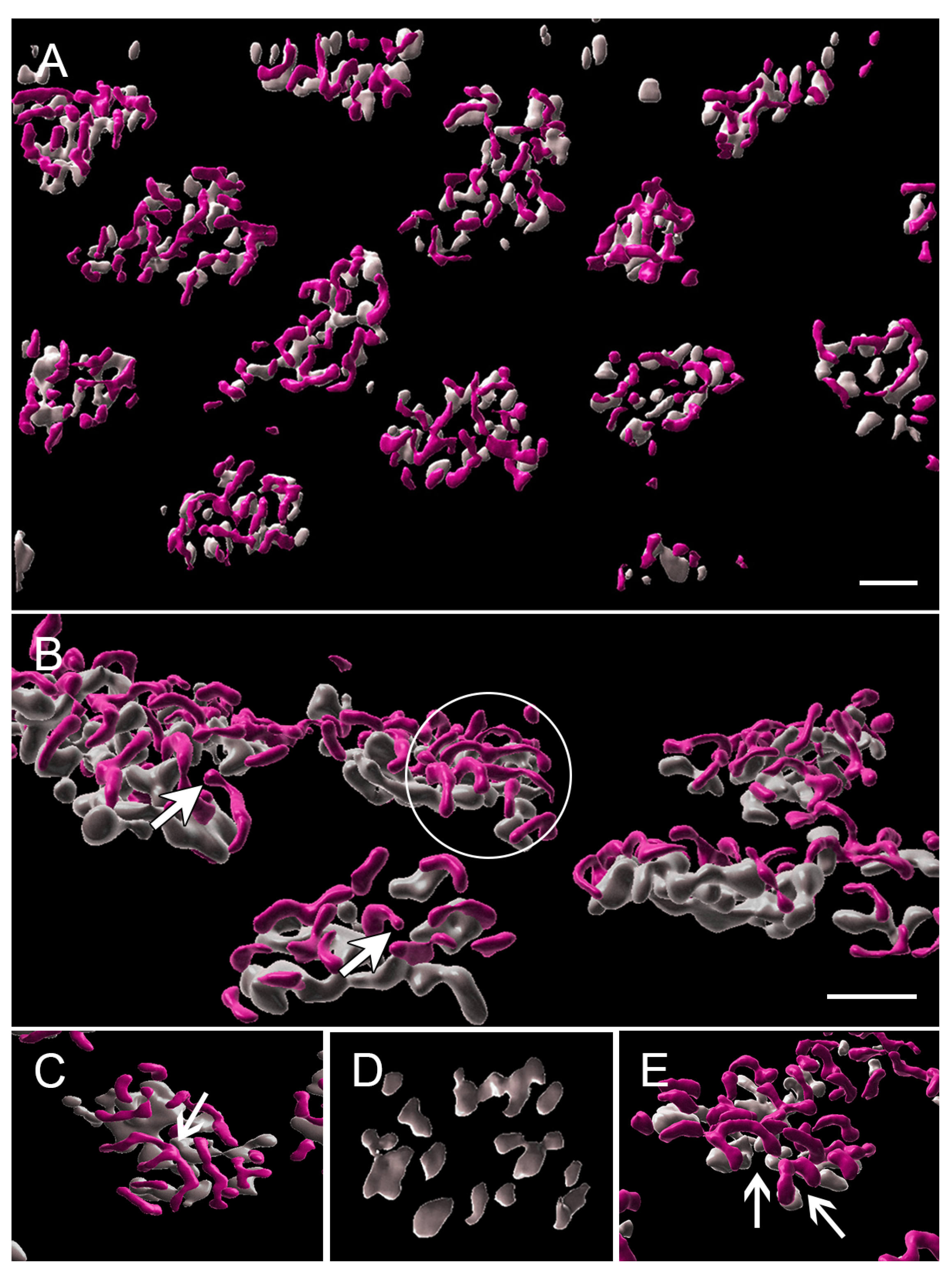
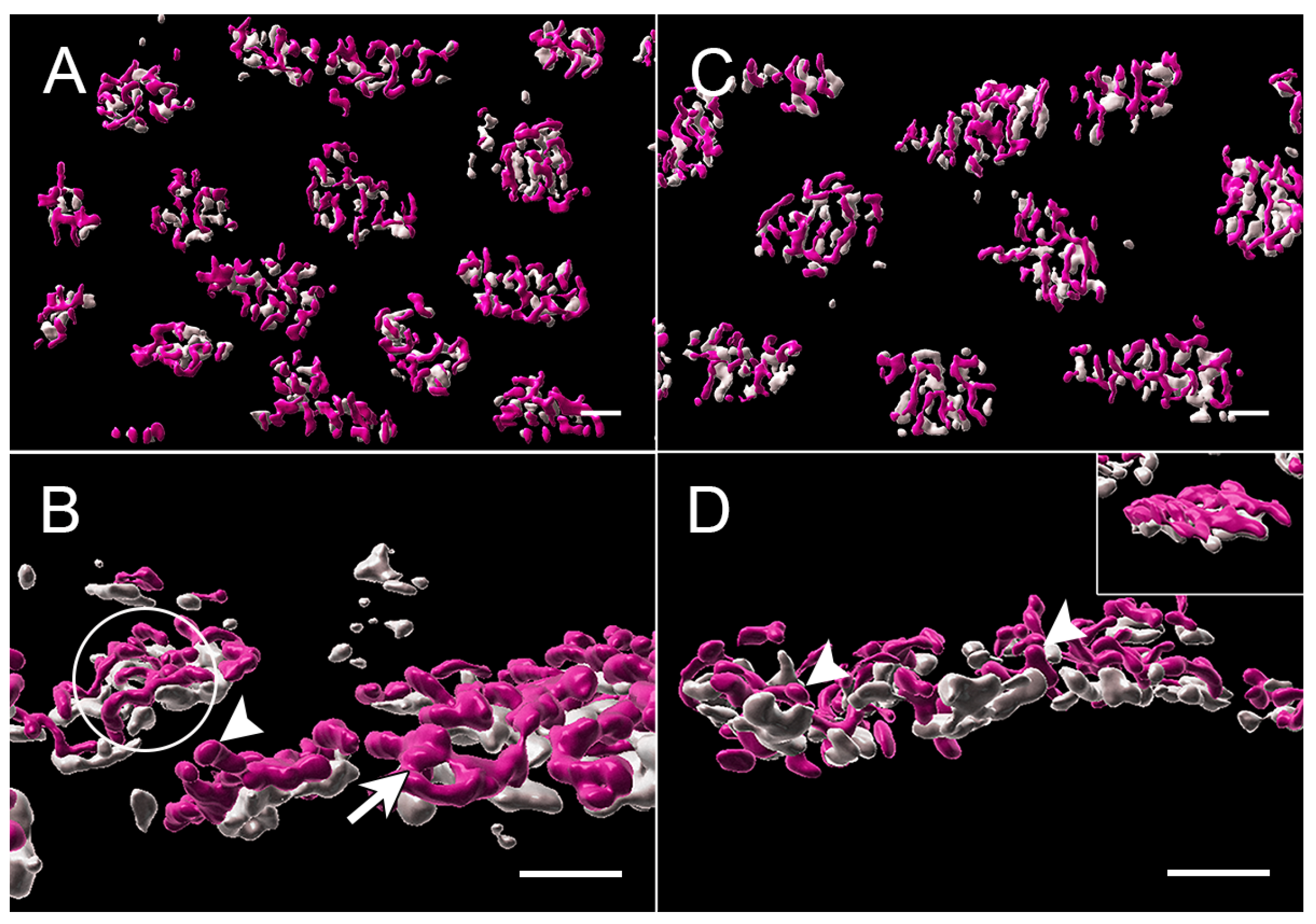
3.1. Normal Cone Synapses
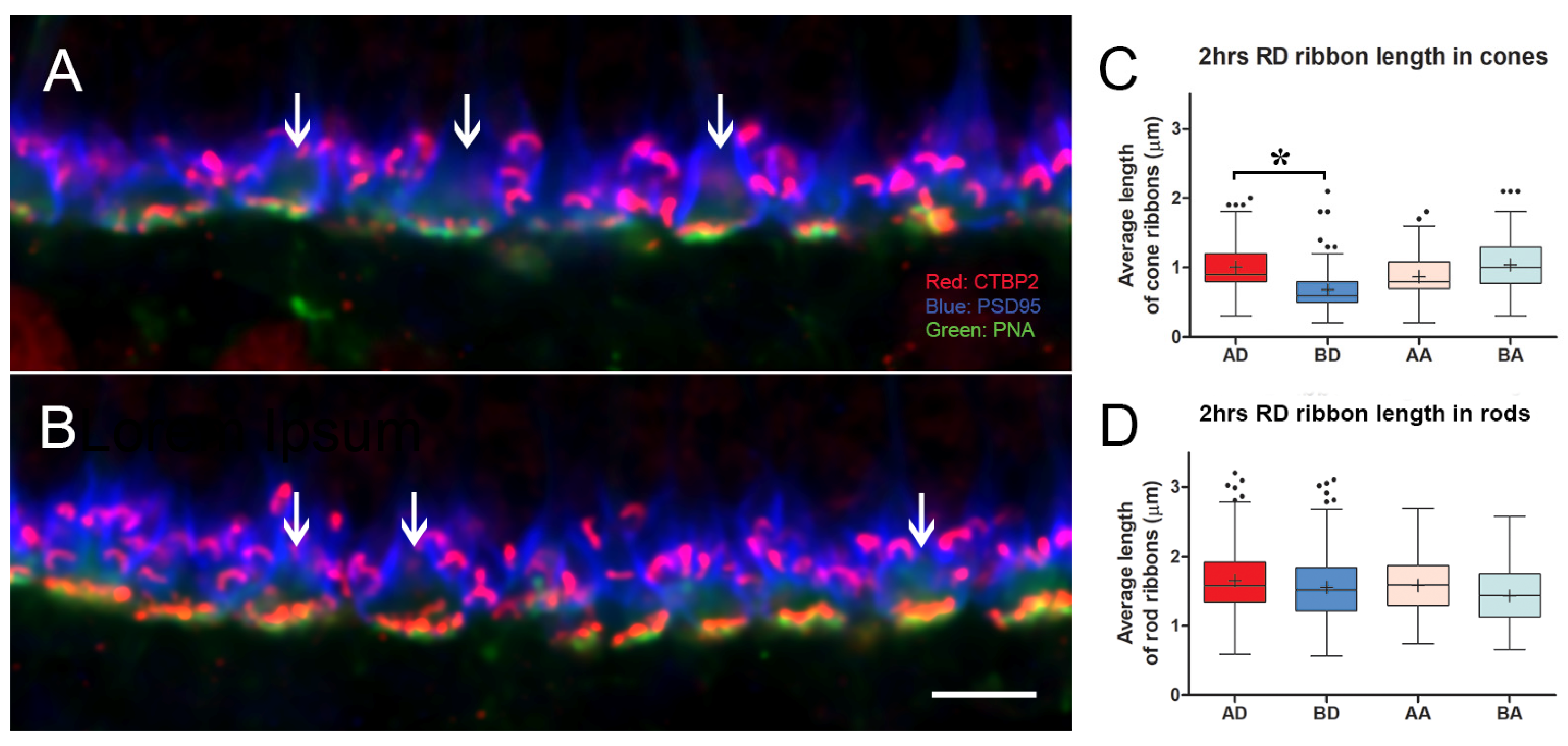
3.2. Intravitreal Administration of AR13503 Decreased the Loss of Cone Ribbons in 2-h Detachments
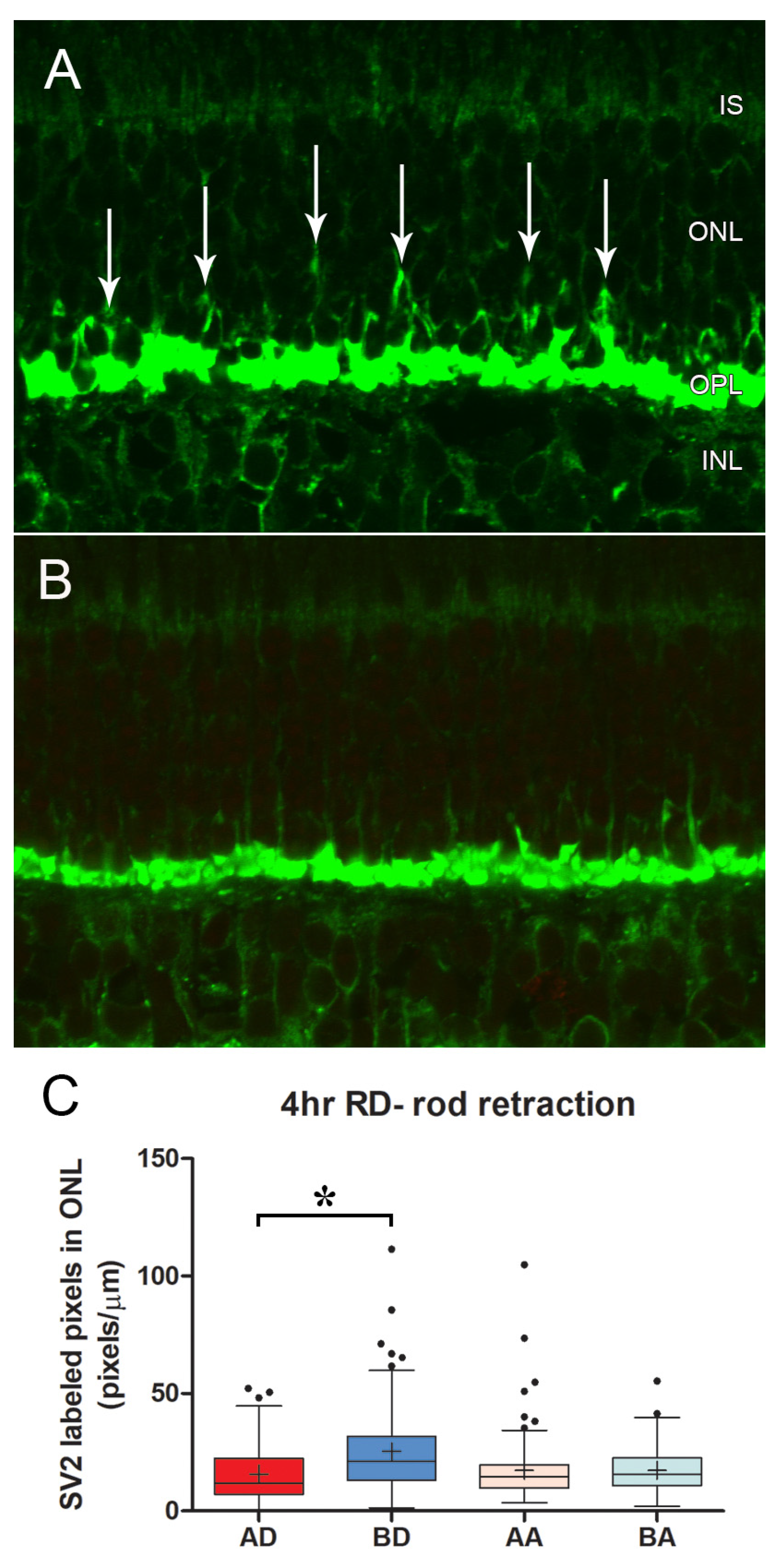
3.3. Delayed Intravitreal Administration of AR13503 Decreased Injury-Induced Changes in Both Cone and Rod Synapses
3.4. Structural and Functional Changes in Cone Synapses after Retinal Detachment and Spontaneous Retinal Reattachment
3.5. Summary
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal ID | Sex | Notes | Depigmentation |
|---|---|---|---|
| Control (terminal) | |||
| DOD6 | F | Data not used because enucleation was several hours after euthanasia | |
| DOD16 | M | Cone and rod data | |
| DOD21 | M | Cone and rod data | |
| Intravitreal 0.75 μm, 2 h RD (terminal) | |||
| 69 | F | Cone and rod data * | depigmentation in od |
| 70 | F | Cone and rod data * | |
| DOD14 | M | Cone and rod data | |
| Intravitreal 0.5 μm, 4 h RD (terminal) | |||
| DOD2 | M | Cone and rod data | depigmentation in ou |
| DOD3 | M | Rod data | |
| DOD4 | M | Rod data | |
| DOD8 | M | Cone data | |
| DOD12 | M | Cone and rod data | depigmentation in os |
| DOD20 | M | Cone data | |
| Subretinal 0.5 μm, 2 Days (survival) | |||
| 45B | F | Cone ERG data ** | depigmentation in ou |
| 48 | F | Cone ERG data ** | intraretinal hemorrhage in od(BSS) |
| 49 | F | Cone ERG data ** | |
| 50 | F | Cone ERG data ** | depigmentation in ou |
| 74 | F | Cone morphological and ERG data ** | depigmentation in ou |
| DOD17 | M | Cone morphological data | |
| DOD18 | M | Cone morphological data | depigmentation in ou |
References
- Lamb, T.D.; Collin, S.P.; Pugh, E.N., Jr. Evolution of the vertebrate eye: Opsins, photoreceptors, retina and eye cup. Nat. Rev. Neurosci. 2007, 8, 960–976. [Google Scholar] [CrossRef]
- Morshedian, A.; Fain, G.L. The evolution of rod photoreceptors. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372. [Google Scholar] [CrossRef]
- Erickson, P.A.; Fisher, S.K.; Anderson, D.H.; Stern, W.H.; Borgula, G.A. Retinal detachment in the cat: The outer nuclear and outer plexiform layers. Investig. Ophthalmol. Vis. Sci. 1983, 24, 927–942. [Google Scholar]
- Fisher, S.K.; Lewis, G.P.; Linberg, K.A.; Verardo, M.R. Cellular remodeling in mammalian retina: Results from studies of experimental retinal detachment. Prog. Retin. Eye Res. 2005, 24, 395–431. [Google Scholar] [CrossRef]
- Lewis, G.P.; Linberg, K.A.; Fisher, S.K. Neurite outgrowth from bipolar and horizontal cells after experimental retinal detachment. Investig. Ophthalmol. Vis. Sci. 1998, 39, 424–434. [Google Scholar]
- Linberg, K.A.; Lewis, G.P.; Fisher, S.K. Retraction and remodeling of rod spherules are early events following experimental retinal detachment: An ultrastructural study using serial sections. Mol. Vis. 2009, 15, 10–25. [Google Scholar]
- Rex, T.S.; Fariss, R.N.; Lewis, G.P.; Linberg, K.A.; Sokal, I.; Fisher, S.K. A survey of molecular expression by photoreceptors after experimental retinal detachment. Investig. Ophthalmol. Vis. Sci. 2002, 43, 1234–1247. [Google Scholar]
- Wang, J.; Zarbin, M.; Sugino, I.; Whitehead, I.; Townes-Anderson, E. RhoA Signaling and Synaptic Damage Occur Within Hours in a Live Pig Model of CNS Injury, Retinal Detachment. Investig. Ophthalmol. Vis. Sci. 2016, 57, 3892–3906. [Google Scholar] [CrossRef]
- Fontainhas, A.M.; Townes-Anderson, E. RhoA and its role in synaptic structural plasticity of isolated salamander photoreceptors. Invesigt. Ophthalmol. Vis. Sci. 2008, 49, 4177–4187. [Google Scholar] [CrossRef]
- Fontainhas, A.M.; Townes-Anderson, E. RhoA inactivation prevents photoreceptor axon retraction in an in vitro model of acute retinal detachment. Investig. Ophthalmol. Vis. Sci. 2011, 52, 579–587. [Google Scholar] [CrossRef]
- Townes-Anderson, E.; Wang, J.; Halasz, E.; Sugino, I.; Pitler, A.; Whitehead, I.; Zarbin, M. Fasudil, a Clinically Used ROCK Inhibitor, Stabilizes Rod Photoreceptor Synapses after Retinal Detachment. Transl. Vis. Sci. Technol. 2017, 6, 22. [Google Scholar] [CrossRef]
- Wang, W.; Halasz, E.; Townes-Anderson, E. Actin Dynamics, Regulated by RhoA-LIMK-Cofilin Signaling, Mediates Rod Photoreceptor Axonal Retraction After Retinal Injury. Investig. Ophthalmol. Vis. Sci. 2019, 60, 2274–2285. [Google Scholar] [CrossRef]
- Wang, W.; Townes-Anderson, E. LIM Kinase, a Newly Identified Regulator of Presynaptic Remodeling by Rod Photoreceptors After Injury. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7847–7858. [Google Scholar] [CrossRef]
- Lessey, E.C.; Guilluy, C.; Burridge, K. From mechanical force to RhoA activation. Biochemistry 2012, 51, 7420–7432. [Google Scholar] [CrossRef]
- Martin-Camara, O.; Cores, A.; Lopez-Alvarado, P.; Menendez, J.C. Emerging targets in drug discovery against neurodegenerative diseases: Control of synapsis disfunction by the RhoA/ROCK pathway. Eur. J. Med. Chem. 2021, 225, 113742. [Google Scholar] [CrossRef]
- Tolias, K.F.; Duman, J.G.; Um, K. Control of synapse development and plasticity by Rho GTPase regulatory proteins. Prog. Neurobiol. 2011, 94, 133–148. [Google Scholar] [CrossRef]
- Halasz, E.; Zarbin, M.A.; Davidow, A.L.; Frishman, L.J.; Gombkoto, P.; Townes-Anderson, E. ROCK inhibition reduces morphological and functional damage to rod synapses after retinal injury. Sci. Rep. 2021, 11, 692. [Google Scholar] [CrossRef]
- Maguire, A.M.; Simonelli, F.; Pierce, E.A.; Pugh, E.N., Jr.; Mingozzi, F.; Bennicelli, J.; Banfi, S.; Marshall, K.A.; Testa, F.; Surace, E.M.; et al. Safety and efficacy of gene transfer for Leber’s congenital amaurosis. N. Engl. J. Med. 2008, 358, 2240–2248. [Google Scholar] [CrossRef]
- Schwartz, S.D.; Hubschman, J.P.; Heilwell, G.; Franco-Cardenas, V.; Pan, C.K.; Ostrick, R.M.; Mickunas, E.; Gay, R.; Klimanskaya, I.; Lanza, R. Embryonic stem cell trials for macular degeneration: A preliminary report. Lancet 2012, 379, 713–720. [Google Scholar] [CrossRef]
- Braekevelt, C.R. Fine structure of the retinal rods and cones in the domestic pig. Graefes Arch. Clin. Exp. Ophthalmol. 1983, 220, 273–278. [Google Scholar] [CrossRef]
- Gerke, C.G., Jr.; Hao, Y.W.; Wong, F. Topography of rods and cones in the retina of domestic the pig. Hong Kong Med. J. 1995, 1, 302–308. [Google Scholar]
- Simoens, P.; De Schaepdrijver, L.; Lauwers, H. Morphologic and clinical study of the retinal circulation in the miniature pig. A: Morphology of the retinal microvasculature. Exp. Eye Res. 1992, 54, 965–973. [Google Scholar] [CrossRef]
- Fernandez de Castro, J.P.; Scott, P.A.; Fransen, J.W.; Demas, J.; DeMarco, P.J.; Kaplan, H.J.; McCall, M.A. Cone photoreceptors develop normally in the absence of functional rod photoreceptors in a transgenic swine model of retinitis pigmentosa. Investig. Ophthalmol. Vis. Sci. 2014, 55, 2460–2468. [Google Scholar] [CrossRef]
- Petters, R.M.; Alexander, C.A.; Wells, K.D.; Collins, E.B.; Sommer, J.R.; Blanton, M.R.; Rojas, G.; Hao, Y.; Flowers, W.L.; Banin, E.; et al. Genetically engineered large animal model for studying cone photoreceptor survival and degeneration in retinitis pigmentosa. Nat. Biotechnol. 1997, 15, 965–970. [Google Scholar] [CrossRef]
- Burton, T.C. Recovery of visual acuity after retinal detachment involving the macula. Trans. Am. Ophthalmol. Soc. 1982, 80, 475–497. [Google Scholar]
- Campo, R.V.; Sipperley, J.O.; Sneed, S.R.; Park, D.W.; Dugel, P.U.; Jacobsen, J.; Flindall, R.J. Pars plana vitrectomy without scleral buckle for pseudophakic retinal detachments. Ophthalmology 1999, 106, 1811–1815; discussion 1816. [Google Scholar] [CrossRef]
- Murtagh, P.J.; Stephenson, K.A.; Rhatigan, M.; McElnea, E.M.; Connell, P.P.; Keegan, D.J. Rhegmatogenous retinal detachments: Primary reattachment rates and visual outcomes over a 4-year period. Ir. J. Med. Sci. 2020, 189, 355–363. [Google Scholar] [CrossRef]
- Ozgür, S.; Esgin, H. Macular function of successfully repaired macula-off retinal detachments. Retina 2007, 27, 358–364. [Google Scholar] [CrossRef]
- Ross, W.H.; Kozy, D.W. Visual recovery in macula-off rhegmatogenous retinal detachments. Ophthalmology 1998, 105, 2149–2153. [Google Scholar] [CrossRef]
- Salicone, A.; Smiddy, W.E.; Venkatraman, A.; Feuer, W. Visual recovery after scleral buckling procedure for retinal detachment. Ophthalmology 2006, 113, 1734–1742. [Google Scholar] [CrossRef]
- Wykoff, C.C.; Smiddy, W.E.; Mathen, T.; Schwartz, S.G.; Flynn, H.W., Jr.; Shi, W. Fovea-sparing retinal detachments: Time to surgery and visual outcomes. Am. J. Ophthalmol. 2010, 150, 205–210.e202. [Google Scholar] [CrossRef]
- Zabel, P.; Zabel, K.; Kazmierczak, K.; Stankiewicz, M.; Jaworski, D.; Suwala, K.; Buszko, K.; Stafiej, J.; Malukiewicz, G.; Kaluzny, J.J. Vascular density and macular sensitivity in eyes after scleral buckling surgery for macula-on rhegmatogenous retinal detachment. PLoS ONE 2023, 18, e0279683. [Google Scholar] [CrossRef]
- Marmor, M.F.; Fulton, A.B.; Holder, G.E.; Miyake, Y.; Brigell, M.; Bach, M. ISCEV Standard for full-field clinical electroretinography (2008 update). Doc. Ophthalmol. 2009, 118, 69–77. [Google Scholar] [CrossRef]
- Liang, K.-Y.; Zeger, S.L. Longitudinal data analysis using generalized linear models. Biometrika 1986, 73, 13–22. [Google Scholar] [CrossRef]
- Schmitz, F.; Königstorfer, A.; Südhof, T.C. RIBEYE, a component of synaptic ribbons: A protein’s journey through evolution provides insight into synaptic ribbon function. Neuron 2000, 28, 857–872. [Google Scholar] [CrossRef]
- tom Dieck, S.; Altrock, W.D.; Kessels, M.M.; Qualmann, B.; Regus, H.; Brauner, D.; Fejtova, A.; Bracko, O.; Gundelfinger, E.D.; Brandstatter, J.H. Molecular dissection of the photoreceptor ribbon synapse: Physical interaction of Bassoon and RIBEYE is essential for the assembly of the ribbon complex. J. Cell. Biol. 2005, 168, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Blackmon, S.M.; Peng, Y.W.; Hao, Y.; Moon, S.J.; Oliveira, L.B.; Tatebayashi, M.; Petters, R.M.; Wong, F. Early loss of synaptic protein PSD-95 from rod terminals of rhodopsin P347L transgenic porcine retina. Brain Res. 2000, 885, 53–61. [Google Scholar] [CrossRef]
- Koulen, P.; Fletcher, E.L.; Craven, S.E.; Bredt, D.S.; Wässle, H. Immunocytochemical localization of the postsynaptic density protein PSD-95 in the mammalian retina. J. Neurosci. 1998, 18, 10136–10149. [Google Scholar] [CrossRef]
- Blanks, J.C.; Johnson, L.V. Specific binding of peanut lectin to a class of retinal photoreceptor cells. A species comparison. Investig. Ophthalmol. Vis. Sci. 1984, 25, 546–557. [Google Scholar]
- Zenisek, D.; Horst, N.K.; Merrifield, C.; Sterling, P.; Matthews, G. Visualizing synaptic ribbons in the living cell. J. Neurosci. 2004, 24, 9752–9759. [Google Scholar] [CrossRef]
- Kerov, V.; Laird, J.G.; Joiner, M.L.; Knecht, S.; Soh, D.; Hagen, J.; Gardner, S.H.; Gutierrez, W.; Yoshimatsu, T.; Bhattarai, S.; et al. α2δ-4 Is Required for the Molecular and Structural Organization of Rod and Cone Photoreceptor Synapses. J. Neurosci. 2018, 38, 6145–6160. [Google Scholar] [CrossRef] [PubMed]
- Haverkamp, S.; Grünert, U.; Wässle, H. The synaptic architecture of AMPA receptors at the cone pedicle of the primate retina. J. Neurosci. 2001, 21, 2488–2500. [Google Scholar] [CrossRef] [PubMed]
- Sherry, D.M.; Mitchell, R.; Standifer, K.M.; du Plessis, B. Distribution of plasma membrane-associated syntaxins 1 through 4 indicates distinct trafficking functions in the synaptic layers of the mouse retina. BMC Neurosci. 2006, 7, 54. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.W.; Sherman, B.; Moore, L.A.; Laethem, C.L.; Lu, D.W.; Pattabiraman, P.P.; Rao, P.V.; deLong, M.A.; Kopczynski, C.C. Discovery and Preclinical Development of Netarsudil, a Novel Ocular Hypotensive Agent for the Treatment of Glaucoma. J. Ocul. Pharmacol. Ther. 2018, 34, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Hendrickson, A.; Hicks, D. Distribution and density of medium- and short-wavelength selective cones in the domestic pig retina. Exp. Eye Res. 2002, 74, 435–444. [Google Scholar] [CrossRef]
- Chidlow, G.; Chan, W.O.; Wood, J.P.M.; Casson, R.J. Differential Effects of Experimental Retinal Detachment on S- and M/L-Cones in Rats. Mol. Neurobiol. 2022, 59, 117–136. [Google Scholar] [CrossRef]
- Nork, T.M. Acquired color vision loss and a possible mechanism of ganglion cell death in glaucoma. Trans. Am. Ophthalmol. Soc. 2000, 98, 331–363. [Google Scholar] [PubMed]
- Nork, T.M.; Millecchia, L.L.; Strickland, B.D.; Linberg, J.V.; Chao, G.M. Selective loss of blue cones and rods in human retinal detachment. Arch. Ophthalmol. 1995, 113, 1066–1073. [Google Scholar] [CrossRef]
- Fisher, S.K.; Lewis, G.P. Muller cell and neuronal remodeling in retinal detachment and reattachment and their potential consequences for visual recovery: A review and reconsideration of recent data. Vis. Res. 2003, 43, 887–897. [Google Scholar] [CrossRef]
- Fisher, S.K.; Lewis, G.P. Cellular effects of detachemnt and reattachment onthe neural retina and the retinal pigment epithelium. In Retina, 4th ed.; Ryan, S.J., Wilkinson, C.P., Eds.; Sirgical Retina; Mosby: Maryland Heights, MO, USA, 2006; Volume 3, pp. 1991–2012. [Google Scholar]
- Lewis, G.P.; Charteris, D.G.; Sethi, C.S.; Leitner, W.P.; Linberg, K.A.; Fisher, S.K. The ability of rapid retinal reattachment to stop or reverse the cellular and molecular events initiated by detachment. Investig. Ophthalmol. Vis. Sci. 2002, 43, 2412–2420. [Google Scholar]
- Voorn, R.A.; Vogl, C. Molecular Assembly and Structural Plasticity of Sensory Ribbon Synapses—A Presynaptic Perspective. Int. J. Mol. Sci. 2020, 21, 8758. [Google Scholar] [CrossRef] [PubMed]
- Balkema, G.W.; Cusick, K.; Nguyen, T.H. Diurnal variation in synaptic ribbon length and visual threshold. Vis. Neurosci. 2001, 18, 789–797. [Google Scholar] [CrossRef]
- Adly, M.A.; Spiwoks-Becker, I.; Vollrath, L. Ultrastructural changes of photoreceptor synaptic ribbons in relation to time of day and illumination. Investig. Ophthalmol. Vis. Sci. 1999, 40, 2165–2172. [Google Scholar]
- Fuchs, M.; Sendelbeck, A.; Atorf, J.; Kremers, J.; Brandstatter, J.H. Strain differences in illumination-dependent structural changes at mouse photoreceptor ribbon synapses. J. Comp. Neurol. 2013, 521, 69–78. [Google Scholar] [CrossRef]
- Spiwoks-Becker, I.; Glas, M.; Lasarzik, I.; Vollrath, L. Mouse photoreceptor synaptic ribbons lose and regain material in response to illumination changes. Eur. J. Neurosci. 2004, 19, 1559–1571. [Google Scholar] [CrossRef]
- Regus-Leidig, H.; Specht, D.; Tom Dieck, S.; Brandstätter, J.H. Stability of active zone components at the photoreceptor ribbon complex. Mol. Vis. 2010, 16, 2690–2700. [Google Scholar] [PubMed]
- Thoreson, W.B. Kinetics of synaptic transmission at ribbon synapses of rods and cones. Mol. Neurobiol. 2007, 36, 205–223. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.E., Jr.; Perkins, G.A.; Giddabasappa, A.; Chaney, S.; Xiao, W.; White, A.D.; Brown, J.M.; Waggoner, J.; Ellisman, M.H.; Fox, D.A. Spatiotemporal regulation of ATP and Ca2+ dynamics in vertebrate rod and cone ribbon synapses. Mol. Vis. 2007, 13, 887–919. [Google Scholar] [PubMed]
- Muller, T.M.; Gierke, K.; Joachimsthaler, A.; Sticht, H.; Izsvak, Z.; Hamra, F.K.; Fejtova, A.; Ackermann, F.; Garner, C.C.; Kremers, J.; et al. A Multiple Piccolino-RIBEYE Interaction Supports Plate-Shaped Synaptic Ribbons in Retinal Neurons. J. Neurosci. 2019, 39, 2606–2619. [Google Scholar] [CrossRef]
- Sheng, Z.; Choi, S.Y.; Dharia, A.; Li, J.; Sterling, P.; Kramer, R.H. Synaptic Ca2+ in darkness is lower in rods than cones, causing slower tonic release of vesicles. J. Neurosci. 2007, 27, 5033–5042. [Google Scholar] [CrossRef]
- Choi, S.Y.; Jackman, S.; Thoreson, W.B.; Kramer, R.H. Light regulation of Ca2+ in the cone photoreceptor synaptic terminal. Vis. Neurosci. 2008, 25, 693–700. [Google Scholar] [CrossRef] [PubMed]
- Libby, R.T.; Lillo, C.; Kitamoto, J.; Williams, D.S.; Steel, K.P. Myosin Va is required for normal photoreceptor synaptic activity. J. Cell. Sci. 2004, 117, 4509–4515. [Google Scholar] [CrossRef] [PubMed]
- Piccoli, G.; Rutishauser, U.; Bruses, J.L. N-cadherin juxtamembrane domain modulates voltage-gated Ca2+ current via RhoA GTPase and Rho-associated kinase. J. Neurosci. 2004, 24, 10918–10923. [Google Scholar] [CrossRef]
- Iftinca, M.; Hamid, J.; Chen, L.; Varela, D.; Tadayonnejad, R.; Altier, C.; Turner, R.W.; Zamponi, G.W. Regulation of T-type calcium channels by Rho-associated kinase. Nat. Neurosci. 2007, 10, 854–860. [Google Scholar] [CrossRef] [PubMed]
- Davison, A.; Lux, U.T.; Brandstätter, J.H.; Babai, N. T-Type Ca2+ Channels Boost Neurotransmission in Mammalian Cone Photoreceptors. J. Neurosci. 2022, 42, 6325–6343. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, J.T.; Wang, H.; Howard, J.; Garrison, J.C.; Barrett, P.Q. T-type calcium channel regulation by specific G-protein betagamma subunits. Nature 2003, 424, 209–213. [Google Scholar] [CrossRef]
- Brabeck, C.; Beschorner, R.; Conrad, S.; Mittelbronn, M.; Bekure, K.; Meyermann, R.; Schluesener, H.J.; Schwab, J.M. Lesional expression of RhoA and RhoB following traumatic brain injury in humans. J. Neurotrauma 2004, 21, 697–706. [Google Scholar] [CrossRef]
- Haverkamp, S.; Grünert, U.; Wässle, H. The cone pedicle, a complex synapse in the retina. Neuron 2000, 27, 85–95. [Google Scholar] [CrossRef]
- Thoreson, W.B. Transmission at rod and cone ribbon synapses in the retina. Pflug. Arch. 2021, 473, 1469–1491. [Google Scholar] [CrossRef]
- Sterling, P.; Matthews, G. Structure and function of ribbon synapses. Trends Neurosci. 2005, 28, 20–29. [Google Scholar] [CrossRef]
- González-Forero, D.; Montero, F.; García-Morales, V.; Domínguez, G.; Gómez-Pérez, L.; García-Verdugo, J.M.; Moreno-López, B. Endogenous Rho-kinase signaling maintains synaptic strength by stabilizing the size of the readily releasable pool of synaptic vesicles. J. Neurosci. 2012, 32, 68–84. [Google Scholar] [CrossRef] [PubMed]
- Taoufiq, Z.; Eguchi, K.; Takahashi, T. Rho-kinase accelerates synaptic vesicle endocytosis by linking cyclic GMP-dependent protein kinase activity to phosphatidylinositol-4,5-bisphosphate synthesis. J. Neurosci. 2013, 33, 12099–12104. [Google Scholar] [CrossRef] [PubMed]
- Mercer, A.J.; Szalewski, R.J.; Jackman, S.L.; Van Hook, M.J.; Thoreson, W.B. Regulation of presynaptic strength by controlling Ca2+ channel mobility: Effects of cholesterol depletion on release at the cone ribbon synapse. J. Neurophysiol. 2012, 107, 3468–3478. [Google Scholar] [CrossRef] [PubMed]
- Holzhausen, L.C.; Lewis, A.A.; Cheong, K.K.; Brockerhoff, S.E. Differential role for synaptojanin 1 in rod and cone photoreceptors. J. Comp. Neurol. 2009, 517, 633–644. [Google Scholar] [CrossRef]
- Sherry, D.M.; Heidelberger, R. Distribution of proteins associated with synaptic vesicle endocytosis in the mouse and goldfish retina. J. Comp. Neurol. 2005, 484, 440–457. [Google Scholar] [CrossRef]
- McCall, M.A.; Gregg, R.G. Comparisons of structural and functional abnormalities in mouse b-wave mutants. J. Physiol. 2008, 586, 4385–4392. [Google Scholar] [CrossRef]
- Gregg, R.G.; Kamermans, M.; Klooster, J.; Lukasiewicz, P.D.; Peachey, N.S.; Vessey, K.A.; McCall, M.A. Nyctalopin expression in retinal bipolar cells restores visual function in a mouse model of complete X-linked congenital stationary night blindness. J. Neurophysiol. 2007, 98, 3023–3033. [Google Scholar] [CrossRef]
- Pearring, J.N.; Bojang, P., Jr.; Shen, Y.; Koike, C.; Furukawa, T.; Nawy, S.; Gregg, R.G. A role for nyctalopin, a small leucine-rich repeat protein, in localizing the TRP melastatin 1 channel to retinal depolarizing bipolar cell dendrites. J. Neurosci. 2011, 31, 10060–10066. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, Y.; Dunn, H.A.; Orlandi, C.; Shultz, N.; Kamasawa, N.; Fitzpatrick, D.; Li, W.; Zeitz, C.; Hauswirth, W.; et al. Interplay between cell-adhesion molecules governs synaptic wiring of cone photoreceptors. Proc. Natl. Acad. Sci. USA 2020, 117, 23914–23924. [Google Scholar] [CrossRef]
- Rothschild, P.R.; Salah, S.; Berdugo, M.; Gelize, E.; Delaunay, K.; Naud, M.C.; Klein, C.; Moulin, A.; Savoldelli, M.; Bergin, C.; et al. ROCK-1 mediates diabetes-induced retinal pigment epithelial and endothelial cell blebbing: Contribution to diabetic retinopathy. Sci. Rep. 2017, 7, 8834. [Google Scholar] [CrossRef]
- Guarino, B.D.; Paruchuri, S.; Thodeti, C.K. The role of TRPV4 channels in ocular function and pathologies. Exp. Eye Res. 2020, 201, 108257. [Google Scholar] [CrossRef] [PubMed]
- Tura, A.; Schuettauf, F.; Monnier, P.P.; Bartz-Schmidt, K.U.; Henke-Fahle, S. Efficacy of Rho-kinase inhibition in promoting cell survival and reducing reactive gliosis in the rodent retina. Investig. Ophthalmol. Vis. Sci. 2009, 50, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Sethi, C.S.; Lewis, G.P.; Fisher, S.K.; Leitner, W.P.; Mann, D.L.; Luthert, P.J.; Charteris, D.G. Glial remodeling and neural plasticity in human retinal detachment with proliferative vitreoretinopathy. Investig. Ophthalmol. Vis. Sci. 2005, 46, 329–342. [Google Scholar] [CrossRef] [PubMed]













| Cone Pedicles | |||
|---|---|---|---|
| 2 h after Detachment | 4 h after Detachment | 2 days after Detachment and with Reattachment | |
| Reduction in ribbon | |||
| Length | xx | xx | o |
| Number | x | xx | o |
| Curvature | x | xx | x |
| Reduction in area of | |||
| PNA-label | o | x | o |
| Rod Spherules | |||
| Retraction of | |||
| terminal | xx * | xx | x * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Townes-Anderson, E.; Halász, É.; Sugino, I.; Davidow, A.L.; Frishman, L.J.; Fritzky, L.; Yousufzai, F.A.K.; Zarbin, M. Injury to Cone Synapses by Retinal Detachment: Differences from Rod Synapses and Protection by ROCK Inhibition. Cells 2023, 12, 1485. https://doi.org/10.3390/cells12111485
Townes-Anderson E, Halász É, Sugino I, Davidow AL, Frishman LJ, Fritzky L, Yousufzai FAK, Zarbin M. Injury to Cone Synapses by Retinal Detachment: Differences from Rod Synapses and Protection by ROCK Inhibition. Cells. 2023; 12(11):1485. https://doi.org/10.3390/cells12111485
Chicago/Turabian StyleTownes-Anderson, Ellen, Éva Halász, Ilene Sugino, Amy L. Davidow, Laura J. Frishman, Luke Fritzky, Fawad A. K. Yousufzai, and Marco Zarbin. 2023. "Injury to Cone Synapses by Retinal Detachment: Differences from Rod Synapses and Protection by ROCK Inhibition" Cells 12, no. 11: 1485. https://doi.org/10.3390/cells12111485